Summations
-
No overall association of BDNF-related mutations with MDD can be sustained by current evidence.
-
Nevertheless, BDNF-related mutations could play an important role in stratifying MDD patients, as they appear related to brain morphology and clinico-phenotypical variables.
-
An inter-relation or combination of BDNF SNPs with other parameters such as plasma/serum BDNF levels, methylation status and neuromorphology changes warrants further investigation as a combined MDD biomarker approach.
Considerations
-
Only papers identified through PubMed and Embase search were included.
-
There was substantial heterogeneity among included studies.
Introduction
Brain-derived neurotrophic factor (BDNF) is the major neurotrophic factor in the human brain (Thoenen, Reference Thoenen1995) and associated with the survival, development and differentiation of neurons by influencing activity-dependent synaptic plasticity, such as long-term depression, long-term potentiation and memory (Vicario-Abejón et al., Reference Vicario-Abejón, Owens, McKay and Segal2002; Kojima and Mizui, Reference Kojima and Mizui2017). Therefore, BDNF massively influences crucial processes of morphological, functional and neurodevelopmental change (Michaelsen et al., Reference Michaelsen, Zagrebelsky, Berndt-Huch, Polack, Buschler, Sendtner and Korte2010; Nestler et al., Reference Nestler, Hyman, Holtzman and Malenka2015; Sasi et al., Reference Sasi, Vignoli, Canossa and Blum2017; Numakawa et al., Reference Numakawa, Odaka and Adachi2018) and contributes to a balanced, healthy neuronal environment and functionality (Chen et al., Reference Chen, Wu, Hwang and Yang2017a). As a central regulator of neuroplastic changes in the brain, BDNF was readily imagined and successively shown to have associations with different neuropsychiatric disorders, including major depressive disorder (MDD) (Autry and Monteggia, Reference Autry and Monteggia2012; Numakawa et al., Reference Numakawa, Odaka and Adachi2018).
Increased knowledge of fundamental cellular and molecular mechanisms as well as genetic abnormalities of BDNF-related neurotrophic pathways in MDD may aid in finding novel biomarkers and treatment targets. State-of-the-art pharmacological, psychotherapeutic and somatic treatment of MDD is adequate, but unfortunately limited (Pampallona et al., Reference Pampallona, Bollini, Tibaldi, Kupelnick and Munizza2004; Chen et al., Reference Chen, Zhao, Liu, Fan and Xie2017b; Cipriani et al., Reference Cipriani, Furukawa, Salanti, Chaimani, Atkinson, Ogawa, Leucht, Ruhe, Turner, Higgins, Egger, Takeshima, Hayasaka, Imai, Shinohara, Tajika, Ioannidis and Geddes2018) and treatment resistance is a growing issue (Otte et al., Reference Otte, Gold, Penninx, Pariante, Etkin, Fava, Mohr and Schatzberg2016). On the one hand, some existing treatment modalities are postulated to exert their effects via modulation of neurotrophic systems, as partially evidenced for electroconvulsive therapy (ECT) and various chemical antidepressants already in 1995, and ketamine more recently (Nibuya et al., Reference Nibuya, Morinobu and Duman1995; Wilkinson and Sanacora, Reference Wilkinson and Sanacora2016). Furthermore, serum BDNF levels of MDD patients seem to be positively modified by the administration of commonly used chemical antidepressants [e.g. selective serotonin reuptake inhibitors (SSRIs)], at least in some patients (Gonul et al., Reference Gonul, Akdeniz, Taneli, Donat, Eker and Vahip2005; Huang et al., Reference Huang, Lee and Liu2008; Polyakova et al., Reference Polyakova, Stuke, Schuemberg, Mueller, Schoenknecht and Schroeter2015; Shi et al., Reference Shi, Luan, Song and Zhang2020).
On the other hand, modifications of BDNF (serum, genetic, etc.), potentially amenable to treatment, led to the question of whether BDNF in some form is a possible biomarker for MDD (Polyakova et al., Reference Polyakova, Stuke, Schuemberg, Mueller, Schoenknecht and Schroeter2015; Björkholm and Monteggia, Reference Björkholm and Monteggia2016), considering that MDD subjects may have lower central BDNF levels in various brain regions (Dwivedi et al., Reference Dwivedi, Rizavi, Conley, Roberts, Tamminga and Pandey2003; Youssef et al., Reference Youssef, Underwood, Huang, Hsiung, Liu, Simpson, Bakalian, Rosoklija, Dwork, Arango and Mann2018). Genetically, multiple single-nucleotide polymorphisms (SNPs) and other mutations in the BDNF and BDNF receptor genes, as well as methylation status of BDNF promoters, were linked with MDD. This indicates that certain SNP profiles and methylation states may be used as biomarkers for MDD (Dwivedi et al., Reference Dwivedi, Rizavi, Conley, Roberts, Tamminga and Pandey2003; Gonul et al., Reference Gonul, Akdeniz, Taneli, Donat, Eker and Vahip2005; Huang et al., Reference Huang, Lee and Liu2008; Januar et al., Reference Januar, Ancelin, Ritchie, Saffery and Ryan2015; Polyakova et al., Reference Polyakova, Stuke, Schuemberg, Mueller, Schoenknecht and Schroeter2015; Shi et al., Reference Shi, Luan, Song and Zhang2020). Analyses of mutations in the BDNF gene (Czira et al., Reference Czira, Wersching, Baune and Berger2012; Youssef et al., Reference Youssef, Underwood, Huang, Hsiung, Liu, Simpson, Bakalian, Rosoklija, Dwork, Arango and Mann2018; Zhao et al., Reference Zhao, Chen, Yang, Han, Fang, Qiu, Yang, Qiao, Ma, Wang, Jiang, Song, Zhou, Zhang, Chen, Qi, Yang and Pan2018; De Oliveira et al., Reference De Oliveira, Pereira, De Alcantara, Antunes, Caffarena and De Mesquita2019), methylation states (Fuchikami et al., Reference Fuchikami, Morinobu, Segawa, Okamoto, Yamawaki, Ozaki, Inoue, Kusumi, Koyama, Tsuchiyama and Terao2011; Januar et al., Reference Januar, Ancelin, Ritchie, Saffery and Ryan2015; Hsieh et al., Reference Hsieh, Lin, Lee and Huang2019) and serum levels (Gonul et al., Reference Gonul, Akdeniz, Taneli, Donat, Eker and Vahip2005; Huang et al., Reference Huang, Lee and Liu2008; Shi et al., Reference Shi, Luan, Song and Zhang2020) in mostly small, lower-quality studies found that there is an association between certain BDNF profiles (both genetic and serum) and the presence of MDD. Additionally, BDNF SNPs could mediate the severity of depression, irrespective of peripheral levels of classically implicated neurotransmitters (Czira et al., Reference Czira, Wersching, Baune and Berger2012).
rs6265 (Val66Met) is the most well-known and common human SNP in the BDNF gene (De Oliveira et al., Reference De Oliveira, Pereira, De Alcantara, Antunes, Caffarena and De Mesquita2019; NCBI, 2020). Val66Met has been associated with lower serum BDNF levels, identified as a possible risk factor to develop MDD and could be the mediating factor between stress and depression (Youssef et al., Reference Youssef, Underwood, Huang, Hsiung, Liu, Simpson, Bakalian, Rosoklija, Dwork, Arango and Mann2018; Zhao et al., Reference Zhao, Chen, Yang, Han, Fang, Qiu, Yang, Qiao, Ma, Wang, Jiang, Song, Zhou, Zhang, Chen, Qi, Yang and Pan2018).
Various BDNF SNPs (Val66Met, rs7103411 and rs908867) have been correlated with both promotor I and IV methylation status, where in a small study, a consistent association between methylation status and MDD appeared (Januar et al., Reference Januar, Ancelin, Ritchie, Saffery and Ryan2015). Both BDNF promotor I and IV showed stronger methylation in subjects with MDD, which raised the question of whether BDNF methylation status could be used as a biomarker in MDD (Fuchikami et al., Reference Fuchikami, Morinobu, Segawa, Okamoto, Yamawaki, Ozaki, Inoue, Kusumi, Koyama, Tsuchiyama and Terao2011; Song et al., Reference Song, Miyaki, Suzuki, Sasaki, Tsutsumi, Kawakami, Shimazu, Takahashi, Inoue, Kan, Kurioka and Shimbo2014; Januar et al., Reference Januar, Ancelin, Ritchie, Saffery and Ryan2015). Some hints to the neurobiological underpinnings of promotor IV correlations in animal models already exist (Hing et al., Reference Hing, Davidson, Lear, Breen, Quinn, McGuffin and MacKenzie2012). In a brief review of various meta-analyses, BDNF Val66Met was not associated with an increased risk to develop MDD, though associated with treatment response (Kishi et al., Reference Kishi, Yoshimura, Ikuta and Iwata2018). Plasma and serum levels of BDNF ostensibly decrease in patients with acute MDD versus controls, whereas antidepressants and ECT can increase BDNF levels, strengthening evidence of a BDNF biomarker for treatment response and acute presence of MDD, rather than understanding as a risk factor (Sen et al., Reference Sen, Duman and Sanacora2008; Kishi et al., Reference Kishi, Yoshimura, Ikuta and Iwata2018; Ai et al., Reference Ai, Wang, Chen, Wang, Xu, Gan, Li, Gou, Cao, Lv, Chen, Wang, Ma and Kuang2019). However, the exact role of genetic modifications of BDNF in MDD and specifically the relevance of BDNF-related mutations in MDD remains to be elucidated in detail.
While some reviews already evaluated different aspects of BDNF in MDD (e.g. serum levels), the conflicting results and lack of a comprehensive data summary regarding the exact role of BDNF-related polymorphisms in MDD primed us to systematically review the current evidence in detail. We aimed to correlate these data with pre-investigated BDNF aspects (serum levels, etc.) and elucidate whether there is an overall correlation of BDNF-related mutations with MDD, or whether merely sub-aspects matter. To this end, we focused on paramount factors in MDD management, such as overall correlation (disease risk), treatment response, remission and diagnostic modalities and tried to identify significant factors.
Method
In this systematic review, we adhered to the PRISMA statement guidelines (Moher et al., Reference Moher, Liberati, Tetzlaff and Altman2009). We classified studies according to the Oxford Centre for Evidence-Based Medicine levels of evidence (OCEBM, 2009), as the more recent OECBM evidence classification from 2011 (OCEBM, 2011) considers a more generalised, less sub-classified approach to evidence hierarchy. Hereby, we intended to display the differences in sub-levels of evidence with enhanced clarity. Simpler rules for evidence classification may be useful in clinical heuristics, but could potentially oversimplify (Howick et al., Reference Howick, Chalmers, Glasziou, Greenhalgh, Heneghan, Liberati, Moschetti, Phillips and Thornton2011) in the setting of this review.
Search strategy
We conducted a MeSH (Medical Subject Headings)-guided search of the PubMed database. The exact search phrases were as follows: ((brain derived neurotrophic factor[MeSH Terms]) AND (major depressive disorder[MeSH Terms])) AND (single nucleotide polymorphism[MeSH Terms]); ((bdnf[MeSH Terms]) AND (snps[MeSH Terms])) AND (major depressive disorder[MeSH Terms]); ((bdnf[MeSH Terms]) AND (major depressive disorder[MeSH Terms])) AND (single nucleotide polymorphism[MeSH Terms]); ((bdnf[MeSH Terms]) AND (major depressive disorder[MeSH Terms])) AND (single nucleotide polymorphisms[MeSH Terms])), ((bdnf receptor[MeSH Terms]) AND (major depressive disorder[MeSH Terms])) AND (single nucleotide polymorphism[MeSH Terms])). An inquiry using the abbreviations did not result in an increased number of, or different, studies. A MeSH major topics search yielded fewer results. Further, we searched the Embase database utilising the following search phrases: (‘brain derived neurotrophic factor’ AND ‘major depressive disorder’ AND ‘single nucleotide polymorphism’) and (‘bdnf’ AND ‘major depressive disorder’ AND ‘single nucleotide polymorphism’) in order to identify additional records. The date of first search was July 2019. The most recent search was conducted on June 23, 2022. No restrictions on years of dissemination were placed. Considered publications include studies from 2005 until June 23, 2022. No forward or backward citation search beyond a general screening for discussion purposes was executed.
Eligibility criteria
We included any publications that focused on evaluating BDNF-related SNPs in the context of MDD. After screening of records, published comments, editorials, author replies, letters to the editor, meta-analyses, reviews and book chapters were excluded. Non-human studies (animal models, in vitro studies, etc.) and any human studies that included participants diagnosed with neurodevelopmental disorders, major neurocognitive disorders, Parkinson’s disease and brain injury of any kind were excluded. Studies with non-rigorous methodology (i.e. systematic errors), bipolar disorder (BD), mixed depression cases or undifferentiated psychiatric diagnosis were excluded as well. Studies that did not focus on BDNF SNPs in the setting of MDD were excluded due to lacking relevance in the context of this review. Any foreign, non-english studies, conference papers or unpublished preprints were not considered in the setting of this systematic review. One author (J.H) performed the literature search. Studies were screened and eligibility discussed by both authors (J.H. and B.N.). Any disagreements were resolved by discussion until a consensus was reached.
Bias assessment
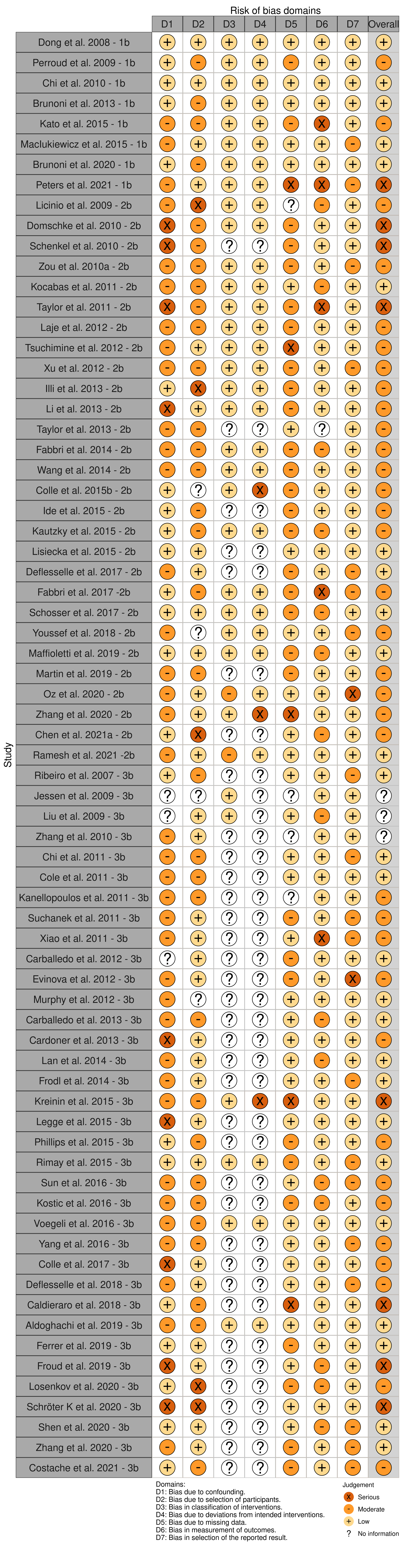
In order to assess for risk of bias in individual studies included and give an overall summary of potential bias in the latter, we utilised the ROBINS-I paradigm for assessment (Sterne et al., Reference Sterne, Hernán, Reeves, Savović, Berkman, Viswanathan, Henry, Altman, Ansari, Boutron, Carpenter, Chan, Churchill, Deeks, Hróbjartsson, Kirkham, Jüni, Loke, Pigott, Ramsay, Regidor, Rothstein, Sandhu, Santaguida, Schünemann, Shea, Shrier, Tugwell, Turner, Valentine, Waddington, Waters, Wells, Whiting and Higgins2016) and the novel robvis tool for easy visualisation of the conducted bias assessment (McGuinness & Higgins, Reference McGuinness and Higgins2021). robvis is an open-source tool that allows for plastic and instant visualisation of potential bias sources identified by means of colour-coding. Various paramount aspects of study quality and methodology were assessed as stringently as possible with the ROBINS-I paradigm for bias due to confounding, bias in selection of participants in the study, bias in classification of interventions, bias due to missing data, bias in measurement of outcomes and bias in selection of reported results. Furthermore, a judgement of overall bias is given.
Results
Our search yielded 427 records in total. After removing duplicates, screening and application of our eligibility criteria, 71 studies were included in the analysis (Fig. 1).

Fig. 1. The PRISMA flow diagram.
Of the 71 studies included, 20 were clinical trials, 17 were cohorts and 34 were case–control studies.
Clinical trials
Treatment response in MDD
Dong et al. (Reference Dong, Wong and Licinio2009) looked at various sequence variations in genes deemed potentially important in the neurobiology of MDD after re-sequencing in 272 Mexican-American MDD patients and 264 healthy controls. Afterwards, in a double-blind RCT setting, after a single-blind placebo lead in phase, patients were randomised to either receive 10-40mg of fluoxetine/day or desipramine 50-200mg/day. One hundred and forty-two MDD patients who enrolled in the pharmacogenetic trial completed 8-week antidepressant treatment (68 treated with desipramine and 74 treated with fluoxetine). Re-sequencing various genes, including neurotrophic receptor tyrosine kinase type 2 (NTRK2, TrkB), resulted in 204 novel SNPs. Two common 3’ UTR polymorphisms in NTRK2 (rs7020204 and rs2013566) were significantly associated with the diagnosis of MDD. Furthermore, regression analyses found that a relative reduction of HAM-D21 scores was associated with six NTRK2 SNPs. Among the SNPs, only two NTRK2 SNPs, rs2289657 and rs56142442, remained statistically significant after correcting for multiple testing in the sample of patients treated with desipramine. Desipramine-treated patients who are homozygous for C allele of rs2289657 or rs56142442 had a 27% higher reduction in Hamilton Depression Rating Scale (HAM-D) scores. Lastly, two haplotypes in NTRK2 (TCG and CAG) in block 3 for desipramine-treated patients were asscoited with remission status.
While BDNF has already been genetically associated with various neurological and psychiatric disorders, Licinio et al. (Reference Licinio, Dong and Wong2009) did a deep sequencing analysis of the BDNF gene with the aim of finding new polymorphisms. Two hundred seventy-two Mexican-American MDD patients and 264 healthy controls were enrolled in this antidepressant treatment trial. Here, 83 novel SNPs were identified: 4 in coding sequences, 30 in untranslated regions, 37 in introns and 12 laying in upstream regions. Three of four rare novel coding SNPs were found to be non-identical. There was a significant association of six NPs with MDD (rs12273539, rs11030103, rs6265, rs28722151, rs41282918 and rs11030101) and two haplotypes in different blocks (one including Val66 and another near exon VIIIh) were significantly linked with MDD, too. Furthermore, an untranslated region SNP (rs61888800) was associated with antidepressant response even after adjusting for a variety of factors including: sex, medication, age as well as baseline scores on the HAM-D scale.
Zou et al. (Reference Zou, Wang, Liu, Feng, Wang, Zang, Yu, Wei, Liu, Liu, Tao, Li, Li, Hu, Li, Zhang, Ye and Xu2010a) ran a controlled clinical trial including 294 MDD patients, providing relatively high-quality evidence on the association of BDNF Val66Met polymorphism with depression severity as well as the efficacy and side effects of fluoxetine treatment. The Chinese cohort showed no association between the efficacy of the SSRI fluoxetine and the BDNF Val66Met polymorphism, but a marginally positive result in favour of remission in heterozygous patients with the Val/Met genotype, in comparison with the Val/Val genotype, where the Val/Met genotype had a significantly higher rate of remission at Week 6. A non-significant trend towards a higher reduction in HAM-D total score and a higher rate of response after 6 weeks of medication in patients carrying the BDNF Val/Met genotype in comparison with the Val/Val homozygotes was observed.
A Taiwanese group (Chi et al., Reference Chi, Chang, Lee, Lee, Gean, Yang, Lu and Chen2010) looked at the association of BDNF Val66Met with MDD in 117 patients and 106 healthy controls and then investigated short-term antidepressant response of these patients to either fluoxetine or venlafaxine in a randomized controlled trial (RCT). While they only found a trend for better treatment response to fluoxetine in Val homozygotes, in the venlafaxine-treated group, Val homozygotes had a significantly higher chance to respond as measured by the HAM-D score.
Domschke et al. (Reference Domschke, Lawford, Laje, Berger, Young, Morris, Deckert, Arolt, McMahon and Baune2010) performed a controlled clinical trial involving 268 (256 for pharmacogenetic analysis) German patients diagnosed with MDD as well as 424 healthy controls. The association of three BDNF polymorphisms including rs7103411, Val66Met (rs6265) and rs7124442 with MDD and antidepressant treatment response was evaluated. Additionally, 10 BDNF markers and their influence on outcome in the context of citalopram treatment in the STAR*D trial sample was analysed by the team. In this context, BDNF was not associated with a diagnosis of MDD, and Domschke et al. reported no support of an (overall) association between genetic variation in BDNF and antidepressant treatment response or remission. However, the BDNF rs7124442 TT genotype was significantly related to worse treatment outcome over a 6-week period in MDD, particularly in anxious depression in this sample of Germans. BDNF SNPs rs7103411 and rs6265 both predicted worse treatment response over 6 weeks in one clinical subtype of depression, namely melancholic depression. All SNPs had major effects on antidepressant treatment response in ANOVA models (with remaining SNPs as covariate variables). The STAR*D analyses did not yield significant data on any of the 10 BDNF markers. Conclusively, there was preliminary support for a potential minor role that genetic variation in BDNF may play in antidepressant treatment outcomes, specifically in the melancholic and anxious depression subtypes.
A small clinical sample of 60 Japanese MDD patients treated with paroxetine were evaluated by Tsuchimine et al. (Reference Tsuchimine, Yasui-Furukori, Nakagami, Sato and Kaneko2012) concerning the association of dopamine receptor D3 (DRD3) and BDNF Val66Met genotype with treatment response to paroxetine. No significant differences in Montgomery–Åsberg Depression Rating Scale (MADRS) scores or clinical improvement related to the BDNF Val66Met polymorphism were found.
Laje et al. (Reference Laje, Lally, Mathews, Brutsche, Chemerinski, Akula, Kelmendi, Simen, McMahon, Sanacora and Zarate2012) report on another small clinical MDD sample (n = 62) treated with intravenous ketamine infusion (0.5mg/kg over 40 minutes) and the effect of BDNF Val66Met on treatment response to ketamine. Twenty-eight per cent of the variance in ketamine response was attributed to genotype in this sample. Mean baseline and end point HAM-D scores for Met carriers were 22.9 and 17.8, respectively. Mean baseline and end point scores for Val carriers were 20.8 and 12.2, respectively. The mean percent change in scores (improvement) was 24% for the Met carriers and 41% for the Val carriers. In the Caucasian group only (n = 58), the mean change was 20% for the Met carriers (n = 18) and 40% for the Val carriers (n = 40).
Brunoni et al. (Reference Brunoni, Kemp, Shiozawa, Cordeiro, Valiengo, Goulart, Coprerski, Lotufo, Brunoni, Perez, Fregni and Benseñor2013) re-evaluated a patient cohort from a RCT named SELECT-TDCS. They focused on the impact that BDNF (Val66Met) and 5-HTTLPR polymorphisms have on MDD and treatment with sertraline versus transcranial direct current stimulation (tDCS). One hundred and twenty MDD patients [moderate-to-severe major depressive episode (MDE)], antidepressant-free, from the previous trials which evaluated the efficacy of sertraline versus tDCS versus placebo (in different combinations) were included. The Val66Met SNP was not significantly associated with response to treatment. However, the serotonin transporter length polymorphic region (5-HTTLPR) polymorphism predicted the tDCS effects as long/long homozygotes showed more improvement when comparing the active versus sham tDCS. The short-allele carriers did not display this effect.
Another trial looking at the influence of BDNF polymorphisms [as well as glial cell line-derived neurotrophic factor (GDNF)] on SSRI treatment response in MDD was conducted by Wang et al. (Reference Wang, Xu, Chen, Xia and Liu2014). This clinical trial involved 298 Chinese MDD patients who were clinically assessed, genotyped and administered 20 mg of paroxetine and followed up after 6 weeks (including plasma paroxetine-level assessment). At the 6-week follow-up, 219 of the 298 patients (73.5%) were responders and 79 patients (26.5%) were non-responders to paroxetine treatment. The allele types for the SNPs rs6265, rs2973049 and rs2216711 demonstrated significant associations with paroxetine treatment remission at Week 6. In addition, the lower threshold concentration of paroxetine for response was 50 ng/mL, where a linear relationship was detected between paroxetine plasma concentration and clinical response.
Fabbri et al. (Reference Fabbri, Marsano, Albani, Chierchia, Calati, Drago, Crisafulli, Calabrò, Kasper, Lanzenberger, Zohar, Juven-Wetzler, Souery, Montgomery, Mendlewicz and Serretti2014) analysed the relationship between several SNPs and treatment response and remission at 4 weeks in several samples of treatment-resistant MDD patients (N = 4414) and found evidence for the involvement of rs11030101 and rs 11030104.
Pooling the data from two RCTs, a Japanese group (Kato et al., Reference Kato, Serretti, Nonen, Takekita, Wakeno, Azuma and Kinoshita2015) looked at 168 MDD patients treated with either paroxetine (n = 81), fluvoxamine (n = 42) or milnacipran (n = 45) and searched for significant associations with various SNPs, as considered relevant according to genomic meta-analyses. They found no significant effect of BDNF Val66Met on treatment response.
In a naturalistic, open-label, non-randomised trial, Kreinin et al. (Reference Kreinin, Lisson, Nesher, Schneider, Bergman, Farhat, Lejbkowicz, Yadid, Raskin, Koman and Pinhasov2015) looked at 51 MDD patients and 38 controls in order to clarify the association between serum BDNF level, severity of depression and the BDNF Val66Met SNP. They found a gender-specific positive correlation between serum BDNF level and severe depression amongst untreated women. However, they found no effect of the BDNF Val66Met SNP on severity, duration of illness, age or BDNF serum levels.
A European group of Maciukiewicz and associates (Maciukiewicz et al., Reference Maciukiewicz, Marshe, Tiwari, Fonseka, Freeman, Rotzinger, Foster, Kennedy, Kennedy and Müller2015), in double-blind RCT, looked at the effect of various SNPs in inflammatory genes and BDNF Val66Met on treatment response to duloxetine (60mg) in 215 MDD patients versus placebo (235 MDD patients). They found no significant association of BDNF Val66Met with treatment response to duloxetine in this European, mostly Caucasian sample.
In a nested analysis from the STAR*D trial, looking at 220 almost exclusively Caucasian patients with treatment-resistant major depressive disorder (trMDD), Fabbri et al. (Reference Fabbri, Crisafulli, Calati, Albani, Forloni, Calabrò, Martines, Kasper, Zohar, Juven-Wetzler, Souery, Montgomery, Mendlewicz and Serretti2017) showed that the Val allele of Val66Met was associated with remission to venlafaxine. In plus, they found that the BDNF rs11030104 SNP A allele was associated with remission to venlafaxine.
An Italian group (Maffioletti et al., Reference Maffioletti, Gennarelli, Gainelli, Bocchio-Chiavetto, Bortolomasi and Minelli2019) looked at the influence of baseline serum BDNF level and Val66Met status on treatment response to ECT in trMDD patients (n = 74, 73% female). They found no significant influence of either one on outcome to ECT treatment in trMDD patients.
In a recent paper, Brunoni et al. (Reference Brunoni, Carracedo, Amigo, Pellicer, Talib, Carvalho, Lotufo, Benseñor, Gattaz and Cappi2020) reported on an ancillary study to the ELECT-TDCS trial which looked for associations between various SNPs [including BDNF, serotonin receptor type 2a (HTR2A), tryptophan hydroxylase 1 (TPH1), 5-HTTLPR and catechol-O-methyl-transferase (COMT)] with outcomes of tDCS and escitalopram (a SSRI) versus placebo. Patients (n = 195 available) were randomised to receive escitalopram/tDCS-sham (n = 75), tDCS/placebo-pill (n = 75) or placebo-pill/sham-tDCS (n = 45). Group comparisons (tDCS vs. placebo, tDCS vs. escitalopram and escitalopram vs. placebo) did not find that any alleles associated with depression improvement. Additionally, exploratory analyses also did not identify any SNP unequivocally associated with improvement of depression in any treatment group.
In a recent RCT, Peters et al. (Reference Peters, Xavier, Mondin, Cardoso, Ferreira, Teixeira, Gräeff, Quevedo, Jansen, Souza, Oses, Pinheiro, da Silva and Ghisleni2021) looked at the effects of the Val66Met SNP on resilience scores, and whether this link could explain variations in response to cognitive therapy. They enrolled 106 MDD patients, which were psychometrically evaluated and underwent molecular analysis from blood samples. It was shown that Met carriers had higher baseline resilience scores than Val homozygotes. Furthermore, there was a statistically significant interactive effect of Val66Met with sex to predict an increase in total resilience scores during cognitive treatment. This meant that female Met allele carriers had higher resilience scores in response to cognitive therapy.
In a recent nested genome-wide association study (GWAS) in the context of a clinical trial on subanaesthetic ketamine infusions for trMDD, Chen et al. (Reference Chen, Kao, Tsai, Li, Lin, Hong, Bai, Tu and Su2021a) analysed over 600.000 SNPs in 65 trMDD patients from their clinical trial. Twelve candidate genes were selected for the study, including BDNF, NTRK2 and multiple other signalling pathway genes, where 388 SNPs were mapped onto those genes. They found statistically significant associations for multiple SNPs in the BDNF-TrkB cascade, as well as glutamatergic and GABAergic pathways in association with treatment response to low-dose ketamine infusions.
Suicidality in MDD
Perroud et al. (Reference Perroud, Aitchison, Uher, Smith, Huezo-Diaz, Marusic, Maier, Mors, Placentino, Henigsberg, Rietschel, Hauser, Souery, Kapelski, Bonvicini, Zobel, Jorgensen, Petrovic, Kalember, Schulze, Gupta, Gray, Lewis, Farmer, McGuffin and Craig2009) aimed to elucidate whether there are genetic predictors of suicidality in MDD patients in the context of antidepressant treatment. Seven hundred ninety-six adult MDD patients were treated with an adjustable dosage of escitalopram or nortriptyline in the context of the GENDEP trial. Nine candidate genes were included based on previous research on the genetics of suicidality and suicidal ideation. Using logistic regression models, 123 SNPs of the 9 candidate genes were compared between subjects, which showed increased suicidal ideation, and those without any increase in suicidal ideation. BDNF polymorphisms were shown to be significantly associated with increased suicidal ideation. The strongest association was observed for rs962369 in BDNF. Also, there was a significant interaction between variants in BDNF and NTRK2.
Schosser et al. (Reference Schosser, Carlberg, Calati, Serretti, Massat, Spindelegger, Linotte, Mendlewicz, Souery, Zohar, Montgomery and Kasper2017) performed a clinical trial in the context of a European multi-centre study on resistant depression, which included 250 patients with MDD, adequately treated with an antidepressant for at least 4 weeks. The patients were then being evaluated using psychometric scaling on depression and suicidality to analyse an association between BDNF polymorphisms and suicidality in MDD. On this cohort, genotyping was performed for the BDNF Val66Met polymorphism as well as seven additional BDNF SNPs (rs11030096, rs925946, rs10501087, rs12273363, rs908867, rs1491850 and rs1491851). None of the BDNF polymorphisms and neither haplotypes were associated with suicide risk and lifetime history of suicide attempts. However, the Val66Met as well as rs10501087 polymorphism (genotypic + haplotypic association) were significantly associated with suicide risk in remitting MDD cases (13,6 %).
Cohort studies
Treatment response in MDD
Kocabas et al. (Reference Kocabas, Antonijevic, Faghel, Forray, Kasper, Lecrubier, Linotte, Massat, Mendlewicz, Noro, Montgomery, Oswald, Snyder, Zohar and Souery2011) set out to analyse treatment response in association with BDNF polymorphisms in a naturalistic setting. Eight SNPs (rs11030096, rs925946, rs10501087, rs6265, rs12273363, rs908867, rs1491850 and rs1491851) including Val66Met were analysed in 206 MDD patients as well as 76 control patients. Neither Val66Met nor any of the other seven SNPs were significantly associated with MDD after permutation correction. An association for rs10501087 & Val66Met with non-response to antidepressant treatment was found. Combined SNP analysis yielded a three-marker combination, which showed significance in the mediation of treatment response (rs10501087, Val66Met and rs1491850).
One hundred fifty-nine Chinese Han MDD patients (78 men and 81 women) were included in a naturalistic, prospective cohort by Xu et al. (Reference Xu, Lin, Rao, Dang, Ouyang, Mai and Zhang2012) in an effort to elucidate the effect that the BDNF Val66Met polymorphism may have on antidepressant [SSRIs vs. serotonin and norepinephrine reuptake inhibitors (SNRIs)] treatment response. The change of depression scoring at week 4 in the SSRI group [total n = 104, with fluoxetine (n = 6), fluvoxamine (n = 3), citalopram (n = 51), paroxetine (n = 17) and sertraline (n = 27)] was significantly different between respective genotype groups. More specifically, changes have been significantly higher among Met carriers versus Val/Val homozygotes. In addition, the rate of response in Met/Met genotype was marginally higher (though reaching statistical significance) than that in Val/Met carriers and Val/Val carriers (89.7 vs. 70.8 and 63%). The adjusted odds ratio (OR) for response was 4.85 in Met allele carriers compared with Val/Val genotype carriers (adjusted for age at onset, number of previous depressive episodes, initial severity of the depression and gender). However, no significant difference in improvement was found between BDNF genotype groups in the SSRI-treated group at week 6. On the other hand, for the venlafaxine-treated group (n = 55), the changes of depression scores after 4 weeks and 6 weeks of antidepressant treatment were not significantly different according to BDNF genotype.
A Finnish group around Illi et al. (Reference Illi, Viikki, Poutanen, Setälä-Soikkeli, Nuolivirta, Kampman, Leinonen, Huhtala, Mononen and Lehtimäki2013) conducted a prospective treatment study involving 106 Finnish MDD patients and 386 healthy controls. The endeavour was initiated in order to examine the role of two BDNF gene polymorphisms (rs11030101 and rs61888800) in relation to the response to SSRIs (citalopram, fluoxetine or paroxetine). Patients were clinically evaluated, genotyped as well as prescribed SSRI antidepressants and re-evaluated at 3 and 6 weeks post-treatment initiation. Three hundred eighty-six control DNA samples were genotyped. No significant differences in the distribution of the two BDNF polymorphisms in the patient population were found, neither in relation to remission nor in relation to response to treatment with an SSRI. Additionally, there were no significant differences between the patient group and the controls.
In a naturalistic treatment study by Colle et al. (Reference Colle, Gressier, Verstuyft, Deflesselle, Lépine, Ferreri, Hardy, Guilloux, Petit, Fève, Falissard, Becquemont and Corruble2015b) on BDNF-related mutations in MDD, their group evaluated the remission of Caucasian MDD patients in association with the Val66Met polymorphism. Three hundred forty-five Caucasian MDD subjects were included, genotyped and administered either an SSRI or an SNRI/tricyclic antidepressant (TCA). Response as well as remission were explained by Val66Met genotype. SSRI-treated patients with Val/Val genotype had a higher response rate at 3-month post-treatment, than Met carriers (68.1% vs. 44%; adjusted-OR: 3.04). In the SNRI/TCA group, Val/Val patients had a lower remission rate 6 months post-treatment versus Met carriers (33.3% vs. 60.9%, adjusted OR: 0.27, CI (95%) [0.09; 0.76]).
Kautzky et al. (Reference Kautzky, Baldinger, Souery, Montgomery, Mendlewicz, Zohar, Serretti, Lanzenberger and Kasper2015) used computer algorithms and machine learning in order to find an association of 12 prominent SNPs (previously associated with MDD and including BDNF Val66Met) with treatment response in a sample of 225 trMDD patients within the European Group for The Study of Resistant Depression (GSRD). In this study, it was shown that in 62% of patients the allelic combination of GG-GG-TT for rs6265 (BDNF), rs7430 [protein kinase C gamma type (PPP3CC)] and rs6313 (HTR2A) for the respective genes and without the clinical feature of melancholia showed a significant prediction of treatment response (HAM-D decline under 17; compared to about 34% of the whole study sample). With a random forests prediction model, 25% of responders could be correctly identified.
In another naturalistic study from Deflesselle et al. (Reference Deflesselle, Verstuyft, Petit, Hardy, David, Falissard, Fève, Rigal, Becquemont, Corruble and Colle2017), 569 MDD patients were genotyped for eight NTRK2 SNPs (rs1187352, rs1439050, rs1778933 rs2289656, rs2289657, rs2289658, rs3824519 and rs56142442) and prospectively assessed response and remission after 6 months of antidepressant treatment. No effects of the NTRK2 SNPs on 6-month response or remission were found.
In a Turkish cohort consisting of 133 MDD patients, Oz et al. (Reference Oz, Baskak, Uckun, Artun, Ozdemir, Ozel, Ozguven and Suzen2020) investigated the frequency of sexual dysfunction in the context of SSRI (either sertraline or citalopram) treatment with relation to serotonin and BDNF pathway polymorphisms. While there were some positive associations for serotonin-related SNPs, no association with BDNF polymorphisms was found.
Ramesh et al. (Reference Ramesh, Venkatesan, Chellathai and Silamban2021) analysed a cohort of 50 Indian MDD patients with moderate to severe illness regarding both serum biomarker levels and BDNF polymorphisms in association with treatment response to SSRIs. Treatment responders showed significantly increased serum BDNF and decreased high-sensitivity c-reactive protein (hsCRP) levels versus non-responders. Furthermore, Val homozygous patients showed greater reduction in HAM-D scores than Met carriers did.
Treatment resistance, recurrence and remission in MDD
Concerning remission from a MDE and the BDNF Val66Met polymorphism, Taylor et al. (Reference Taylor, McQuoid, Ashley-Koch, MacFall, Bridgers, Krishnan and Steffens2011) evaluated the association of Val66Met genotype, white matter lesions and clinical depression assessment in a prospective cohort of 229 elderly Caucasian subjects. At the 3-month evaluation, the BDNF Val66Met genotype was not associated with remission. When multiple comparisons were not controlled for, Met carriers were more likely to experience remission at 6 months with an OR of 1.82. Even when controlling for lesion volume and social support, this effect remained. Alas, elderly MDD subjects, which are Met carriers, had higher odds of remission than Val homozygotes.
A multi-centre, longitudinal study conducted by Li et al. (Reference Li, Zhang, Wang, Chen, Fan, Guan, Zhang, Yuan, Hong, Wang, Wu, Huang, Hu, Cao, Yi, Cui, Yu and Fang2013) provided data on the role of BDNF gene, the BDNF receptor gene (NTRK2) and their interaction in the development of trMDD. Nine hundred forty-eight MDD patients were recruited for the 12-week longitudinal study. Patients were clinically evaluated, genotyped and followed over the course of 12 weeks. There was a significant association between NTRK2 allele rs1565445 and trMDD with an excess of the T allele in the trMDD group, compared to non-trMDD group (OR = 1.43). Furthermore, patients with genotype C/C and T/C in NTRK2 rs1565445 were less likely to develop trMDD than those carrying T/T (OR = 0.52; OR = 0.72, respectively). The haplotype T-T (NTRK2 SNPs rs1565445 and rs1387923) had 1.41x increased risk of trMDD. Lastly, there were significant four-locus (rs1387923-rs1565445-rs2769605-rs6265) gene-gene interactions as being detected by the multifactor-dimensionality reduction method. In conclusion, BDNF and NTRK2 polymorphisms may significantly interact in and influence the development of treatment resistance in MDD.
As treatment resistance to SSRIs in MDD is not rare and predictive factors are lacking until now, Zhang et al. (Reference Zhang, Li, Pang, Zhao, Cao, Wang, Wang and Li2020b) took several tagSNPs and used a support vector machine (SVM) learning classifier on 857 patients with recurrent MDD to find potential predictive SNPs. They found multiple accurately predictive models, and that adding the combined BDNF (BDNF: rs18035210 and rs7124442) + cAMP response element-binding protein (CREB1) SNPs (CREB1: rs2551645 and rs4675690) into the SVM prediction model significantly increased classification accuracy.
Suicidality in MDD
Schenkel et al. (Reference Schenkel, Segal, Becker, Manfro, Bianchin and Leistner-Segal2010) performed a retrospective cohort study on 120 patients who were admitted to the Emergency Hospital of Porto Alegre, Brazil, due to a suicide attempt. Though BD was allowed for in this cohort, the regression model excluded both BD and alcohol abuse as variates; hence, we included this study in our review. Nonetheless, the following results have to be interpreted with utmost caution: initial univariate analyses showed that sex, BDNF genotype, intent and method of suicide attempt were all risk factors for high lethality in suicide attempts. After logistic regression analysis, male sex (OR = 3.03) and the Met allele (OR = 2.62) were significantly and independently associated with the high lethality in suicide attempts. This showed that a Met carrier state is an independent predictor of high lethality in suicide attempts of depressed (alas MDD, since neuropsychiatric comorbidities showed no regression model-associated relevance) patients.
A retrospective cohort study by Youssef et al. (Reference Youssef, Underwood, Huang, Hsiung, Liu, Simpson, Bakalian, Rosoklija, Dwork, Arango and Mann2018) looked at the association of the BDNF Val66Met polymorphism, cerebral BDNF levels with MDD, suicide and early-life adversity (ELA). For this purpose, post-mortem brain tissue from 37 suicide decedents and 53 non-suicide decedents was analysed. BDNF levels were determined by Western blot in various limbic and non-limbic structures implicated in mood control and associated with MDD: (A) dorsolateral prefrontal cortex (dlPFC), (B) anterior cingulate cortex (ACC), (C) caudal brainstem, and (D) rostral brainstem. Overall, subjects with the Met allele had an increased risk for depression. MDD patients (N = 45) had lower BDNF levels in the ACC and caudal brainstem than non-depressed subjects (N = 45). No association of history of suicide death or ELA with genotype was observed. Subjects who had been exposed to ELA and/or died by suicide were shown to have lower BDNF levels in the ACC when compared to non-suicide decedents and subjects with no reported ELA.
Neuroimaging and BDNF in MDD
Taylor et al. (Reference Taylor, Zhao, Ashley-Koch, Payne, Steffens, Krishnan, Hauser and MacFall2013) set out to investigate the relationship between magnetic resonance imaging (MRI)-detected white matter hyperintensities, BDNF Val66Met, the angiotensin II receptor type 1 (AGTR1) A1166C polymorphism and depression. They conducted a retrospective cohort study including 54 depressed and 37 non-depressed elderly subjects. High-resolution MRI as well as genotyping (only 62 Caucasian subjects of 91 subjects were genotyped: 29 depressed subjects + 33 non-depressed elderly subjects) was performed. The depressed cohort exhibited a significantly greater lesion ratio only in the left upper cingulum near the cingulate gyrus, supporting past work implicating cingulate dysfunction in the pathogenesis of depression. BDNF Met allele carriers exhibited greater lesion ratios only in the frontal corpus callosum. AGTR1 C1166 carriers exhibited greater lesion ratios across multiple tracts including the anterior thalamic radiation and inferior fronto-occipital fasciculus. Hence, different fibre tract lesions may relate to genetic status as well as vascular processes in late-life depression.
A Japanese prospective cohort by Ide et al. (Reference Ide, Kakeda, Watanabe, Yoshimura, Abe, Hayashi, Ueda, Kishi, Katsuki, Umene-Nakano, Iwata, Nakamura and Korogi2015) set out to investigate the relationship between Val66Met genotype and voxel-based morphology (VBM) findings [similar to Cardoner et al. (Reference Cardoner, Soria, Gratacòs, Hernández-Ribas, Pujol, López-Solà, Deus, Urretavizcaya, Estivill, Menchón and Soriano-Mas2013)] for 38 first episode and generally antidepressant/drug-naïve MDD patients and 42 matched, healthy controls. The entirety of the cohort was stratified according to BDNF genotype, and subsequently the effects of diagnosis and genotype, as well as the genotype–diagnosis interaction in relation to brain morphology, were assessed. Met carriers with MDD had a significantly smaller middle frontal gyrus on VBM analysis versus healthy controls. Ergo, the BDNF polymorphism was linked with PFC atrophy in MDD patients. There was a significant difference in overall grey matter volume in MDD subjects versus healthy controls.
Lisiecka et al. (Reference Lisiecka, O’Hanlon, Fagan, Carballedo, Morris, Suckling and Frodl2015) demonstrated another neurophysiological alteration in genotype-stratified subgroups from 37 MDD patients and 39 healthy controls in a small prospective cohort. They assessed respective functional MRI (fMRI) patterns in response to an emotional stimulus (emotional scene/visual) and compared the results according to the respective genotype. Met carriers with MDD had increased activation in subcortical regions responsible for visceral reaction to emotional stimuli. Val/Val homozygotes with MDD were associated with having decreased neural activation in areas responsible for cognitive appraisal of emotional scenes. During a MDE Met carriers displayed higher levels of activation in brain areas associated with cognitive appraisal of emotional information in comparison to Val/Val homozygous individuals with a current MDE. It follows that allelic variations of Val66Met may lead to specific neural activation patterns, correlating with MDD. Hence, different mechanisms of MDD in the two allelic groups regarding neural circuit activation/inhibition may exist, which could become a useful tool in MDD screening.
Insulin resistance and MDD treatment
Martin et al. (Reference Martin, Colle, El Asmar, Rigal, Vievard, Feve, Becquemont, Verstuyft and Corruble2019) looked at the influence of BDNF Val66Met genotype on insulin resistance in MDD patients (n = 148). Met carriers had a higher risk of insulin resistance after antidepressant treatment, indicating the need for insulin resistance screening in antidepressant-treated MDD patients, specifically Met carriers.
Case–control and cross-sectional studies
Overall association of BDNF with MDD (disease risk)
Ribeiro et al. (Reference Ribeiro, Busnello, Cantor, Whelan, Whittaker, Deloukas, Wong and Licinio2007) conducted a case–control study reporting on 278 Mexican-American MDD patients and 320 controls of the same ethnicity, which were assessed clinically by HAM-D rating as well as genotyping. BDNF Val66Met SNP showed a significant association with the diagnosis of MDD for a single-marker association test between MDD and the control group. Here, Val/Val homozygous individuals were 70% more likely to be depressed, supporting an association of BDNF Val66Met with MDD in this ethnic group.
Liu et al. (Reference Liu, Xu, Jiang, Cui, Qian and Jiang2009) ran a case-only cross-sectional study on 105 Chinese trios (grandparents, parents and children; N = 315) in order to elucidate the involvement of various BDNF polymorphisms in MDD. They did a single-marker, paired marker as well as haplotype analysis of three BDNF SNPs: Val66Met, rs10835210 and rs2030324. There were no significant associations of the three SNPs with MDD. Pairwise analysis revealed substantial linkage disequilibrium among three SNPs. Multiple-marker transmission-disequilibrium testing (TDT) indicated that there was no association of the haplotypes and MDD. As statistical power was adequate, it was concluded that SNPs rs6265, rs10835210 and rs2030324 of the BDNF gene are unlikely to play a critical role in the pathogenesis of MDD in Chinese Han patients.
As gene–gene interactions of singularly pre-identified risk genes were considered likely to play a role in the mediation of an association with MDD, Zhang et al. (Reference Zhang, Yang, Xu, Sun, Yang, Liu, Xu and Shen2010) looked at whether BDNF and BDNF signalling cascade polymorphisms [glycogen synthase kinase-3 beta (GSK3B) and protein kinase B alpha (AKT1)] may have an association with MDD. Four hundred forty-seven Chinese Han patients with MDD and 432 age- and gender-matched control subjects were recruited, and association analyses were conducted. Five SNPs were utilised. An allelic association between the GSK3B SNP rs6782799 and MDD was found in the sample with an OR of 1.25. There was a significant effect of a two-locus BDNF/GSK3B interaction with MDD (GSK3B rs6782799 and BDNF rs7124442), as well as three-locus interaction (GSK3B rs6782799, BDNF rs6265 and BDNF rs7124442). Combination of two risk alleles showed an OR of 4.00, while combination of three risk alleles gave an OR of 4.46. It was concluded that indeed GSK3B alleles are a significant susceptibility factor in Chinese Han patients suffering from MDD.
On a Polish population of 116 in-patients with recurrent MDD and 218 healthy subjects, Suchanek et al. (Reference Suchanek, Owczarek, Kowalczyk, Kucia and Kowalski2011) performed a case–control study in order to evaluate whether there is an association between the BDNF Val66Met and BDNF rs28383487 SNP and MDD. The entire sample was genotyped. Whole-group, group-stratified as well as sex-stratified analyses were conducted. This study failed to find an association between C-281A polymorphism with recurrent MDD. There was a significant association between Val66Met and MDD however. The Val/Val genotype was more frequent in MDD patients compared to the control group (regarding both total analysis and after sex-based stratification). The Val allele is connected to a higher risk of recurrent MDD development in men than in women. Correspondence analysis has shown that the co-presence of genotypes Val/Val and C/C is connected with a higher risk of recurrent MDD development compared to other genotype combinations.
In a study on Taiwanese population conducted by Chi et al. (Reference Chi, Lee, Chang, Yang, Lin and Chen2011), there were no significant differences between MDD patients (N = 198) and controls (N = 106) regarding the BDNF rs6265 polymorphism, but when the researchers compared the allele distribution in their control group with other populations from the HapMap database, namely the Han Chinese in Beijing, US residents with European ancestry, Japanese in Tokyo, and Yoruba in Ibadan, they found significant differences.
A Slovak study conducted by Evinova et al. (Reference Evinova, Babusikova, Straka, Ondrejka and Lehotsky2012) on 134 inpatients diagnosed with MDD and 143 healthy controls showed that there is no significant difference in between these groups concerning the BDNF G196A polymorphism.
As a complex interplay between genes and environmental factors likely contributes to the development of MDD, a Chinese group (Yang et al., Reference Yang, Sun, Liu, Li, Xu and Zhang2016) looked at the interactive effects of the BDNF gene, PRKCG gene and negative life events in 406 Chinese MDD patients and 391 controls. They found that there was a significant gene × environment interaction of negative life events score, BDNF Val66Met and the PRKCG rs3745406 SNP. Furthermore, in a subanalysis, both the Val and Met allele of the BDNF SNP significantly influenced the risk of MDD in interaction with higher levels of negative life events and the C allele of PRKCG.
Serotonin transporter as well as BDNF polymorphisms were investigated by Sun et al. (Reference Sun, Yang, Liu, Li, Xu and Zhang2016) in a Chinese Han population. Four hundred fifty-nine MDD patients as well as 412 healthy controls were selected and genotyped for the 5-HTTLPR and Val66Met polymorphisms. BDNF Val66Met showed no correlation with MDD. When interaction with BDNF was modelled, for individuals with BDNF (rs6265), genotype GG, cases in the heterozygous group had even higher odds of MDD than those in the combined homozygous group of 5-HTTLPR polymorphism, suggesting that there may be significant interactions between the 5-HTT and BDNF gene in relation to MDD.
One of the more recent case–control studies by Aldoghachi et al. (Reference Aldoghachi, Tor, Redzun, Lokman, Razaq, Shahbudin, Badamasi, Cheah, Stanslas, Veerakumarasivam, Rosli, Ibrahim, Lye and Ling2019) evaluated three BDNF polymorphisms (though all but Val66Met were excluded from further analysis due to monomorphism) and plasma BDNF levels in 300 Malaysian MDD patients (where most were on pharmacotherapy: only 7,4% had either no medications or unknown status) versus 300 matched controls. High-resolution melting (HRM) and statistical analysis showed that BDNF Val66Met in the Malaysian population increases the odds of developing MDD by 2.05-fold. Two hundred and six randomly selected cases as well as 206 randomly selected controls underwent plasma BDNF assessment and a significant decrease in plasma BDNF levels of MDD cases as versus controls was observed. Nonetheless, there was no evidence of the effect of the Val66Met genotype on the plasma BDNF level. Hence, another type of variable may be the mediator of effect in lower plasma BDNF levels in MDD versus healthy subjects without MDD.
Looking at 138 MDD patients (only 41 male) and 94 controls (27 male), Losenkov et al. (Reference Losenkov, Mulder, Levchuk, Vyalova, Loonen, Bosker, Simutkin, Boiko, Bokhan, Wilffert, Hak, Schmidt and Ivanova2020) found no significant association of Val66Met genotype with diagnosis; however they found an association with depression severity as measured by HAMD-17. Additionally, the group did not find an association of plasma BDNF level with either diagnosis or severity of MDD.
In a cohort of 259 young Chinese Han people (105 MDD patients and 154 healthy controls), Zhang et al. (Reference Zhang, Ran, Ai, Wang, Chen, Wu, Liu, Jin, Wang and Kuang2020a) found no significant associations of geno- or haplotype frequencies with MDD after targeted sequencing (and false discovery rate correction) of the BDNF gene.
In a recent study, a Romanian group looked at various genetic polymorphisms, including Val66Met in the context of MDD (Costache et al., Reference Costache, Riza, Popescu, Streaţă, Dincă, Glăvan, Vladu, Udriştoiu and Ioana2021). Their study included a small sample of 82 MDD subjects and 286 healthy controls. No significant allelic, recessive or dominant association could be demonstrated for Val66Met or other investigated SNPs.
A group around Schröter and colleagues looked at allelic and genotype frequencies of BDNF Val66Met in 49 MDD patients and 57 controls (as well as BD patients). They found no statistically significant difference in neither genotype nor allelic frequency between any of the three groups.
Treatment resistance, recurrence and remission in MDD
In an effort to elucidate the hypothesis of stress having neurotoxic effects and influencing neurotrophic signalling pathways, Xiao et al. (Reference Xiao, Liu, Gao, Wan, Yang, Wang, Wang, Wang and Liu2011) analysed whether CRHR1 (corticotropin-releasing hormone receptor 1) polymorphisms (rs1876828, rs242939 and rs242941) and the BDNF Val66Met polymorphism influence susceptibility to recurrent depression. One hundred and eighty-one patients with recurrent MDD as well as 186 controls were clinically evaluated and genotyped. Here, CRHR1 (rs1876828 and rs242941) and BDNF (rs6265) alleles were found to have no association with the risk of recurrent MDD, while an allelic association between CRHR1 rs242939 and recurrent MDD was found with an OR of 0.454.
Suicidality in MDD
Voegeli et al. (Reference Voegeli, Ramoz, Shekhtman, Courtet, Gorwood and Kelsoe2016) took a prospective sample of 78 MDD patients and 312 controls in order to elucidate the effect of various SNPs on antidepressant-worsening suicidal ideation (AWSI), a worrisome, clinically relevant phenomenon, which occurs with antidepressant treatment in some patients. Patients were taken from the GENESE sample. They looked at various BDNF as well as NTRK2 SNPs and found one SNP of the latter (NTRK2 rs1439050), which remained significantly associated with AWSI after correction for potential confounders.
In a nested analysis from the METADAP study, Deflesselle et al. (Reference Deflesselle, Colle, Rigal, David, Vievard, Martin, Becquemont, Verstuyft and Corruble2018) were interested in the influence of TrkB-related SNPs on suicide risk in 624 patients with MDD with a current MDE. Indeed, the TRKB rs2289656 CC genotype was associated with a 2.2-fold higher risk of acute suicide attempt in their study.
Neuroimaging and BDNF in MDD
Jessen et al. (Reference Jessen, Schuhmacher, von Widdern, Guttenthaler, Hofels, Suliman, Scheef, Block, Urbach, Maier and Zobel2009), building on work of Frodl et al. (Reference Frodl, Schüle, Schmitt, Born, Baghai, Zill, Bottlender, Rupprecht, Bondy, Reiser, Möller and Meisenzahl2007) who ostensibly first reported on the association of the BDNF Val66Met polymorphism with hippocampal volume in MDD, did a replication case–control study in a German sample. Seventy-nine MDD patients and 84 healthy controls were recruited and genotyping for the BDNF Val66Met polymorphism, as well as high-resolution MRI was performed. Hippocampal volumes of MDD patients were significantly smaller than those of the controls, confirming previous reports. After adjusting for genotype, however, there was no Val66Met effect on hippocampal volume in either group.
Kanellopoulos et al. (Reference Kanellopoulos, Gunning, Morimoto, Hoptman, Murphy, Kelly, Glatt, Lim and Alexopoulos2011) did a covariance analysis of 33 elderly MDD patients as well as 23 controls without psychiatric disease, based on structural MRI-based data of the hippocampal volumes (right and left), Val66Met genotype (Val/Val and Met carrier) and diagnosis (depressed and non-depressed). Additionally, age, gender, education, as well as whole brain volume were included as covariates. Elderly BDNF Val/Val homozygotes with MDD had significantly higher right hippocampal volumes compared with non-depressed Val homozygous controls. There was no difference between the depressed and non-depressed Met carriers. Depressed Met carriers had an earlier age of onset (AOO) of depressive illness than depressed Val/Val homozygotes. It was concluded that Val/Val homozygosity may mediate a BDNF-based neuroprotective role against pathophysiological processes in adults with late-onset depression.
Cole et al. (Reference Cole, Weinberger, Mattay, Cheng, Toga, Thompson, Powell-Smith, Cohen-Woods, Simmons, McGuffin and Fu2011) analysed whether the 5-HTTLPR and/or BDNF Val66Met polymorphisms may influence the hippocampal morphology of patients with MDD in a case–control study. Parallel samples of 84 MDD patients and 111 healthy individuals were collected, and MRI-based (3D) hippocampal shape mapping was performed as well as genotyping conducted. There was no significant difference between 5-HTTLPR short-allele carriers and long/long homozygotes or between BDNF Met allele carriers and Val/Val homozygotes in the group of healthy individuals. Moreover, there was no significant difference in normalised hippocampal volumes between 5-HTTLPR di-allelic and tri-allelic classifications or between the BDNF Val66Met genotypes in MDD patients.
Carballedo et al. (Reference Carballedo, Amico, Ugwu, Fagan, Fahey, Morris, Meaney, Leemans and Frodl2012) performed a MRI-based association study on 37 MDD patients as well as 42 healthy controls in order to elucidate the connection between white matter tract lesions in limbic and frontal areas as well as BDNF Val66Met genotype. A significant interaction was found in the uncinate fasciculus between BDNF Val66Met alleles and diagnosis. Here, Met allele had smaller fractional anisotropy (FA) in the uncinate fasciculus compared to those patients homozygous for Val allele and compared to healthy subjects carrying the Met allele. A significant three-way interaction was detected between the cingulum (dorsal, rostral and parahippocampal regions), brain hemisphere and BDNF genotype. Larger FA was detectable in the left rostral cingulum for Met allele carriers when compared to Val/Val homozygotes. The Met allele of the BDNF polymorphism seems to render subjects more vulnerable to neurophysiological circuitry dysfunctions associated with the uncinate fasciculus, a tract known to be related to negative emotional-cognitive processing bias, declarative memory problems and self-awareness.
Murphy et al. (Reference Murphy, Carballedo, Fagan, Morris, Fahey, Meaney and Frodl2012) were one of the few groups, which analysed the BDNF receptor gene (NTRK2) as well as the BDNF Val66Met genotype in the context brain morpho-functional analyses and genetic background. In a case–control setting, they performed MR imaging as well as genotyping on a group of 45 MDD patients and 45 age- and gender-matched controls. High-angular-resolution diffusion images were obtained and analysed via tract-based spatial statistics, which yielded an interactive effect between NTRK2 and depression diagnosis maximally affecting the cingulum. MDD patients homozygous for the A allele of NTRK2 showed significantly reduced FA compared with depressed patients with at least one copy of the G allele or control subjects with either the A/A or G carrier genotypes in the following regions: left and right corona radiata, left uncinate fasciculus, left inferior fronto-occipital fasciculus, left cerebral peduncle, posterior thalamic radiation, and middle cerebral peduncle. Significantly smaller grey matter volume was seen in frontal lobe regions in patients homozygous for the A allele. Additionally, there was no significant effect of BDNF Val66Met polymorphism or ELA on white matter diffusion.
Cardoner et al. (Reference Cardoner, Soria, Gratacòs, Hernández-Ribas, Pujol, López-Solà, Deus, Urretavizcaya, Estivill, Menchón and Soriano-Mas2013) analysed cerebral morphological differences according to BDNF Val66Met genotype in 37 in-patients with MDD and presence of melancholic features. On MRI (+ VBM), a grey matter volume reduction in the left hippocampus was observed in Met carriers, while there was a volume increase in the right orbitofrontal cortex. This decrease was inversely correlated to days to remission, while a significant negative correlation between left hippocampal volume and days to remission was found in valine homozygotes.
In another study, Carballedo et al. (Reference Carballedo, Morris, Zill, Fahey, Reinhold, Meisenzahl, Bondy, Gill, Möller and Frodl2013) studied the interaction between BDNF Val66Met polymorphism and hippocampal volume in 62 patients with MDD and 71 matched healthy controls from Dublin and Munich. They found no association between BDNF genotype and MDD but showed that Met allele carriers with a history of ELA had significantly lower hippocampal volumes when compared with Met allele carriers without history of ELA. Subjects homozygous for the Val allele did not develop lower hippocampal volumes if they had a history of ELA. These results were found in both MDD patients and healthy controls. However, Met allele carriers without history of ELA had larger hippocampal volumes than participants homozygous for the Val allele without history of ELA. Taken together, these results suggest that Met allele might not be the only factor involved in developing smaller hippocampal volumes.
Lan et al. (Reference Lan, Ogden, Huang, Oquendo, Sullivan, Miller, Milak, Mann and Parsey2014) wanted to elucidate the influence of the BDNF cascade on the serotonergic system. They recruited 50 healthy volunteers and 50 acutely depressed, unmedicated patients with MDD in a case–control study. The entire sample underwent positron emission tomography (PET) scanning with the 5-HT1A receptor ligand, [11C]WAY-100635 and a metabolite-corrected arterial input function. Additionally, subjects were genotyped for the BDNF Val66Met polymorphism. A linear mixed effects (covariate) model compared 5-HT1A receptor binding potential (BP(F)), proportional to the number of available receptors, in 13 brain regions of interest between Met carriers and Val/Val homozygotes using sex and C-1019G genotype of the 5-HT1A receptor promoter functional polymorphism as covariates. There was an interaction between diagnosis and allele, such that Met allele carriers had 17.4% lower BP(F) than non-met carriers in the control group, but not in the MDD group. Their data are consistent with a model where the Met allele of the Val66met polymorphism causes less proliferation of serotonin synapses, and therefore fewer 5-HT1A receptors. In MDD, however, the effect of the BDNF Val66Met polymorphism is not detectable, which may be possibly due to a ceiling effect of over-expression of 5-HT1A receptors in mood disorders.
Frodl et al. (Reference Frodl, Skokauskas, Frey, Morris, Gill and Carballedo2014) looked at the interaction between childhood adversity, BDNF Val66met genotype and the formation/morphology of hippocampal subfields. Thirty-eight MDD patients and 44 matched healthy controls underwent high-resolution MRI. Patients with MDD had significantly smaller cornu ammonis 4/dentate gyrus (CA4/DG) and cornu ammonis 2/3 (CA2/3) volumes compared to healthy controls. Furthermore, there was a significant interactive effect of BDNF allele and childhood adversity on CA2/3 and CA4/DG volumes. Met carriers without childhood adversity had larger and with childhood adversity smaller CA4/DG and CA2/3 volumes than Val/Val homozygotes. This highlighted that stress is relevant for gene interaction effects on hippocampal volume reductions, in particular, for the subfield CA2/3 and the DG.
Another neuromorphology association study was performed by Legge et al. (Reference Legge, Sendi, Cole, Cohen-Woods, Costafreda, Simmons, Farmer, Aitchison, McGuffin and Fu2015). This group looked at the association of brain morphology modifications in extended prefrontal and associated regions in correlation with the BDNF Val66Met polymorphism, more specifically amygdala, anterior cingulate, middle frontal and orbitofrontal regions. MR imaging of 79 subjects with MDD and 74 healthy controls was obtained. Genotyping of the entire sample was performed. It was found that Met carriers showed significantly reduced caudal middle frontal thickness in both groups. Significant interactions were found in the anterior cingulate and rostral middle frontal regions, where in the MDD group Met carriers showed the greatest reduction in surface area.
Phillips et al. (Reference Phillips, Batten, Tremblay, Aldosary, Du and Blier2015) set out to investigate a set of monoaminergic-related genes, which are hypothesised and partially evidenced to be involved in MDD. Twenty-six outpatients with trMDD and 27 matched healthy controls were included in the study. Both groups underwent genotyping for a set of six SNPs in monoaminergic-related genes (serotonin transporter (SLC6A4), norepinephrine transporter (SLC6A2), serotonin 1A and 2A receptors (HTR1A and HTR2A), COMT, and BDNF) as well as MRI (and algorithmic determination of hippocampal volumes) in order to elucidate possible influences of monoaminergic-related gene polymorphisms on MDD. There was some evidence for norepinephrine and serotonin-related genes being involved in MDD; however, there was no association between the 5-HT2A, COMT, and BDNF SNPs and hippocampal volume.
Kostic et al. (Reference Kostic, Canu, Agosta, Munjiza, Novakovic, Dobricic, Maria Ferraro, Miler Jerkovic, Pekmezovic, Lecic Tosevski and Filippi2016) expanded the available literature on the effect of BDNF polymorphisms on brain structure in MDD by not only analysing BDNF SNPs, but rather looking at the cumulative effect various SNPs of candidate genes on MDD and brain morphology/structural integrity. In a case–control setting, the effect of accumulation of SERT, BDNF and COMT gene SNPs on brain morphology was analysed. Seventy-seven MDD patients and 66 healthy controls underwent a clinical psychiatric assessment, genetic testing and MRI scanning. They showed that compared with controls, patients were more BDNF-Val homozygotes, COMT-Met carriers and SERT-L carriers. Subsequently, subjects were split into three groups: 1. high-frequency susceptibility polymorphism group (hfSP, subjects with all three susceptibility polymorphisms (SPs)); 2. intermediate-frequency SP group (ifSP, two SPs); and 3. low-frequency SP group (lfSP, one/no SP). Upon MR imaging, cortical thickness, volume of the hippocampi, amygdala and subcortical structures as well as white matter tract integrity were assessed. Compared to controls, hfSP patients showed thinning of the middle frontal cortex bilaterally, left frontal pole and right lateral occipital cortex and smaller hippocampal volume bilaterally. Both hfSP and lfSP groups showed thinning of the left inferior parietal cortex and reduced WM integrity of the corpus callosum. Compared to patients, hfSP controls showed greater integrity of the fronto-occipital cortices and corpus callosum. Conclusively, it was affirmed that cortical prefrontal and occipital damage in MDD patients is modulated by polymorphism accumulation, while damage to the parietal cortex and corpus callosum seems to be independent of genetic accumulation.
In a case–control study, Shen et al. (Reference Shen, Lu, Jiang, Ye, Zhou, He, Li, Xu and Cheng2020) investigated the effect of the Val66Met SNP on cortical thickness in 105 first-episode, drug-naïve MDD patients and 111 healthy controls. They found a main effect for MDD diagnosis in the left rostral ACC, right inferior temporal and right lateral orbitofrontal, while main genotype effects were observed in the left posterior cingulate cortex. A diagnosis-by-genotype interaction effect was located in the left rostral ACC. Here, MDD Met carriers showed thinner left rACC structures than healthy control Met carriers. However, neither symptom severity nor illness duration correlated significantly with cortical thickness.
Inflammation and MDD
An analysis of association between inflammation, BDNF polymorphisms and serum BDNF levels was concocted by Caldieraro et al. (Reference Caldieraro, McKee, Leistner-Segal, Vares, Kubaski, Spanemberg, Brusius-Facchin, Fleck and Mischoulon2018) in 73 MDD patients. This group found that the Met allele was significantly associated with higher BDNF and lower tumour necrosis factor-alpha (TNF-alpha) levels, even after correction and considering possible confounders. Respectively, a low BDNF and high level of inflammatory markers may be affected by the Val66Met polymorphism and the neurotrophic and inflammatory signalling systems may interact, which has to be analysed in further studies.
Diet in MDD
Dietary quality, BDNF Val66Met polymorphism as well as plasma BDNF levels were recently assessed by Froud et al. (Reference Froud, Murphy, Cribb, Ng and Sarris2019) in 187 MDD patients as well as 55 healthy controls. The control group had a significantly higher dietary quality than the participants with MDD. A logistic regression model (age, sex, serum BDNF levels, dietary quality and depression) concluded that lower dietary quality, and surprisingly, higher BDNF levels, were associated with increased depression risk. Neither seasonality (at time of patient and control recruitment) nor the BDNF Val66Met polymorphism was associated with BDNF levels. Additionally, no evidence of interaction between the Val66Met polymorphism, serum BDNF, dietary quality and depression was demonstrated. Higher dietary quality was associated with decreased depression incidence and severity, plus Val66Met appeared not to predict serum BDNF levels, depression incidence or modify the relationship between dietary quality and BDNF.
Neurocognitive performance and MDD
Ferrer et al. (Reference Ferrer, Labad, Salvat-Pujol, Barrachina, Costas, Urretavizcaya, de Arriba-Arnau, Crespo, Soriano-Mas, Carracedo, Menchón and Soria2019) looked at the effect of BDNF variants and methylation status on cognition in MDD. In a case–control study, their sample included 64 MDD patients and 70 healthy controls. Significant associations between neurocognitive performance and two BDNF SNPs (including rs908867 and rs925946) were found. This effect was significantly mediated by methylation values at (specific) promoter I sites. Furthermore, significant associations between neurocognitive test results and methylation status as well as its interactions with a diagnosis of MDD, sex and Childhood Trauma Questionnaire (CTQ) scores were found.
Childhood and adolescent MDD
A Hungarian group around Rimay et al. (Reference Rimay, Benak, Kiss, Baji, Feher, Juhasz, Strauss, Kennedy, Barr, Kovacs, Vetro and Kapornai2015) performed a retrospective cohort study analysing the influence of Val66Met in a specific subgroup of patients rarely analysed separately: children. Five hundred eighty-three depressed individuals were involved in this study [162 with melancholic subtype; childhood-onset melan cholic depression (COMD)] in an effort to elucidate a possible association between the BDNF Val66Met polymorphism and COMD. Furthermore, it aimed to examine the effect of stressful life events (SLEs) and the Val66Met polymorphism on the risk of COMD. Results showed that 27.8% of the participants fulfilled the criteria for melancholy and the proportion of females amongst them was higher (53.1%). Genotype and allele frequency showed no significant differences when comparing the melancholic with the non-melancholic subtype MDD group (27,8% vs. 72,2%). Additionally, it was ascertained that the Val66Met SNP and SLE interaction was not significantly associated with melancholy as an outcome.
Plasma BDNF level
In a second study by their group, Colle et al. (Reference Colle, Trabado, David, Brailly-Tabard, Hardy, Falissard, Fève, Becquemont, Verstuyft and Corruble2017) tried to elucidate the connection between the BDNF pathway, its components and the inter-relation to clinical features of MDD in 328 MDD patients with a current MDE. They analysed plasma BDNF levels, BDNF genotype for Val66Met polymorphism and performed an extensive clinical psychiatric assessment of the MDD patients. Here, it was shown that plasma BDNF levels were significantly and linearly associated with respective BDNF Val66Met genotypes, where the Met/Met genotype carriers had lower plasma BDNF levels than the Val/Met and Val/Val carriers. Additionally, three clinical characteristics namely AOO, MDD duration and number of previous episodes were associated with Val66Met genotype for plasma BDNF levels. In Met carriers (Met/Met genotype + Met/Val genotype), plasma BDNF levels were negatively correlated with AOO and positively correlated with MDD duration and number of previous MDE (not so for Val/Val genotype).
As numerous variables have been investigated in studies on BDNF-related mutations in MDD, with correspondingly different levels of evidence, Table 1 summarises the evidence for and against associations between specific subgroups and sub-stratifications in MDD according to BDNF polymorphisms.
Table 1. Variable-dependent associations of BDNF-related mutations with MDD

BDNF, brain-derived neurotrophic factor; ELA, early-life adversity; fMRI, functional magnetic resonance imaging; GSK3B, glycogen synthase kinase type 3 beta; GxE, gene–environment interaction; HAM-D, Hamilton Rating Scale for Depression; HPA, hypothalamic–pituitary–adrenal axis; MDD, major depressive disorder; NTRK2, neurotrophic receptor tyrosine kinase type 2; rACC, rostral anterior cingulate cortex; SLE, stressful life events; SNRI, serotonin norepinephrine reuptake inhibitor; SSRI, selective serotonin reuptake inhibitor; TCA, tricyclic antidepressant.
Bias assessment
A sample-size weighted bar graph of percentages displays the fractions of respectively biased studies per evaluated domain (Fig. 2). Additionally, we provide a traffic light plot (Supplementary Figure 1) on the specific outcomes of the ROBINS-I-based bias assessment in the Supplementary Materials. The traffic light plot is ordered by descending quality of evidence; hence, studies with higher level of evidence are situated towards the top of the table. With this methodology of displaying risk of (though not publication) bias in association with evidence quality, we hope to circumvent classical pitfalls of Funnel plots, involving, for example, lack of study precision and poor interpretability by researchers (Terrin et al., Reference Terrin, Schmid and Lau2005; Lau et al., Reference Lau, Ioannidis, Terrin, Schmid and Olkin2006).

Fig. 2. Bias assessment – percentage of bias per domain.
Discussion
According to the overall scientific data, there is a possibly major pathophysiological role for BDNF, neurotrophic systems, and hence neuroplastic changes to play in MDD. However, both pre-existing scepticism on the rectilinearity of BDNF involvement in MDD (Groves, Reference Groves2007) and inconsistent present evidence already point towards a manifold complexity of BDNF in MDD that has only been partially appreciated until now (Castrén, Reference Castrén2005; Groves, Reference Groves2007). Our inter-comparison and chronological synthesis of evidence on the role of BDNF-related mutations in particular shows that likely no overall association of BDNF-related mutations with MDD can be sustained based on accumulated evidence. On the one hand, it can be appreciated that over time evidence emerged on specific significant sub-conditions, stratifications and variables. Somehow counterintuitively tough, it becomes increasingly certain that there is no overall association of BDNF-related mutations with MDD, especially considering higher-quality clinical trials (i.e. RCTs) and meta-analyses (Gratacòs et al., Reference Gratacòs, González, Mercader, de Cid, Urretavizcaya and Estivill2007, Reference Gratacòs, Soria, Urretavizcaya, González, Crespo, Bayés, de Cid, Menchón, Vallejo and Estivill2008; Chen et al., Reference Chen, Lawlor, Lewis, Yuan, Abdollahi, Timpson, Day, Ebrahim, Smith and Shugart2008; López-León et al., Reference López-León, Janssens, González-Zuloeta Ladd, Del-Favero, Claes, Oostra and van Duijn2008; Sullivan et al., Reference Sullivan, de Geus, Willemsen, James, Smit, Zandbelt, Arolt, Baune, Blackwood, Cichon, Coventry, Domschke, Farmer, Fava, Gordon, He, Heath, Heutink, Holsboer, Hoogendijk, Hottenga, Hu, Kohli, Lin, Lucae, Macintyre, Maier, McGhee, McGuffin, Montgomery, Muir, Nolen, Nöthen, Perlis, Pirlo, Posthuma, Rietschel, Rizzu, Schosser, Smit, Smoller, Tzeng, van Dyck, Verhage, Zitman, Martin, Wray, Boomsma and Penninx2009; Verhagen et al., Reference Verhagen, van der Meij, van Deurzen, Janzing, Arias-Vásquez, Buitelaar and Franke2010; Domschke et al., Reference Domschke, Lawford, Laje, Berger, Young, Morris, Deckert, Arolt, McMahon and Baune2010; Lewis et al., Reference Lewis, Ng, Butler, Cohen-Woods, Uher, Pirlo, Weale, Schosser, Paredes, Rivera, Craddock, Owen, Jones, Jones, Korszun, Aitchison, Shi, Quinn, MacKenzie, Vollenweider, Waeber, Heath, Lathrop, Muglia, Barnes, Whittaker, Tozzi, Holsboer, Preisig, Farmer, Breen, Craig and McGuffin2010; Gyekis et al., Reference Gyekis, Yu, Dong, Wang, Qian, Kota and Yang2013; Kishi et al., Reference Kishi, Yoshimura, Ikuta and Iwata2018). This may explain the equivocatory nature of results of even higher-quality studies, reflecting no overall association with MDD, but associations with specific subgroups or variables in MDD, and why the neurotrophin hypothesis of depression as a clear-cut, neurobiological pathway dysfunction is insufficient (Groves, Reference Groves2007). Specifically, a) when considering treatment response and remission (Licinio et al., Reference Licinio, Dong and Wong2009; Zou et al., Reference Zou, Ye, Feng, Su, Pan and Liao2010b; Kocabas et al., Reference Kocabas, Antonijevic, Faghel, Forray, Kasper, Lecrubier, Linotte, Massat, Mendlewicz, Noro, Montgomery, Oswald, Snyder, Zohar and Souery2011; Xu et al., Reference Xu, Lin, Rao, Dang, Ouyang, Mai and Zhang2012) and specific patient subgroups, such as gender- (Verhagen et al., Reference Verhagen, van der Meij, van Deurzen, Janzing, Arias-Vásquez, Buitelaar and Franke2010; Suchanek et al., Reference Suchanek, Owczarek, Kowalczyk, Kucia and Kowalski2011) or ethnicity-stratified groups (Ribeiro et al., Reference Ribeiro, Busnello, Cantor, Whelan, Whittaker, Deloukas, Wong and Licinio2007; Wang et al., Reference Wang, Xu, Chen, Xia and Liu2014; Zou et al., Reference Zou, Wang, Liu, Feng, Wang, Zang, Yu, Wei, Liu, Liu, Tao, Li, Li, Hu, Li, Zhang, Ye and Xu2010a, Reference Zou, Ye, Feng, Su, Pan and Liao2010b; Colle et al., Reference Colle, Gressier, Verstuyft, Deflesselle, Lépine, Ferreri, Hardy, Guilloux, Petit, Fève, Falissard, Becquemont and Corruble2015b), potentially relevant interactions appear. Furthermore, b) in brain morphological studies or functional neurophysiological analyses for detection of depression, BDNF polymorphisms may play a role in identification and/or (sub-) stratification of MDD patients (Kanellopoulos et al., Reference Kanellopoulos, Gunning, Morimoto, Hoptman, Murphy, Kelly, Glatt, Lim and Alexopoulos2011; Carballedo et al., Reference Carballedo, Amico, Ugwu, Fagan, Fahey, Morris, Meaney, Leemans and Frodl2012; Murphy et al., Reference Murphy, Carballedo, Fagan, Morris, Fahey, Meaney and Frodl2012; Cardoner et al., Reference Cardoner, Soria, Gratacòs, Hernández-Ribas, Pujol, López-Solà, Deus, Urretavizcaya, Estivill, Menchón and Soriano-Mas2013; Taylor et al., Reference Taylor, Zhao, Ashley-Koch, Payne, Steffens, Krishnan, Hauser and MacFall2013; Frodl et al., Reference Frodl, Skokauskas, Frey, Morris, Gill and Carballedo2014; Legge et al., Reference Legge, Sendi, Cole, Cohen-Woods, Costafreda, Simmons, Farmer, Aitchison, McGuffin and Fu2015; Ide et al., Reference Ide, Kakeda, Watanabe, Yoshimura, Abe, Hayashi, Ueda, Kishi, Katsuki, Umene-Nakano, Iwata, Nakamura and Korogi2015; Kostic et al., Reference Kostic, Canu, Agosta, Munjiza, Novakovic, Dobricic, Maria Ferraro, Miler Jerkovic, Pekmezovic, Lecic Tosevski and Filippi2016). Lastly, c) an inter-relation or combination with other parameters such as plasma/serum BDNF levels (Sen et al., Reference Sen, Duman and Sanacora2008; Colle et al., Reference Colle, Trabado, David, Brailly-Tabard, Hardy, Falissard, Fève, Becquemont, Verstuyft and Corruble2017; Caldieraro et al., Reference Caldieraro, McKee, Leistner-Segal, Vares, Kubaski, Spanemberg, Brusius-Facchin, Fleck and Mischoulon2018) or methylation status (Carlberg et al., Reference Carlberg, Scheibelreiter, Hassler, Schloegelhofer, Schmoeger, Ludwig, Kasper, Aschauer, Egger and Schosser2014; Ferrer et al., Reference Ferrer, Labad, Salvat-Pujol, Barrachina, Costas, Urretavizcaya, de Arriba-Arnau, Crespo, Soriano-Mas, Carracedo, Menchón and Soria2019), representing a potential future combinatory biomarker approach, warrants further investigation as well. Especially, since methylation status could potentially differentiate, not only against healthy controls, but BD patients as well (Carlberg et al., Reference Carlberg, Scheibelreiter, Hassler, Schloegelhofer, Schmoeger, Ludwig, Kasper, Aschauer, Egger and Schosser2014).
Though results are ambiguous in nature (Elfving et al., Reference Elfving, Buttenschøn, Foldager, Poulsen, Andersen, Grynderup, Hansen, Kolstad, Kaerlev, Mikkelsen, Thomsen, Børglum, Wegener and Mors2012; Halaris et al., Reference Sharma, Meresh, Ghanshyam, Kang, Hage, Garlenski and Sinacore2015; Froud et al., Reference Froud, Murphy, Cribb, Ng and Sarris2019), peripheral BDNF levels on their own seem unlikely to predict depression accurately in a differentiated manner (Terracciano et al., Reference Terracciano, Piras, Lobina, Mulas, Meirelles, Sutin, Chan, Sanna, Uda, Crisponi and Schlessinger2013; Molendijk et al., Reference Molendijk, Spinhoven, Polak, Bus, Penninx and Elzinga2014). Hence, we hypothesise that combined BDNF diagnostics could increase accuracy and precision of sub-stratifying MDD patient groups, differentiating amongst polarity, severity and versus healthy controls, plus identifying treatment resistance [consider Shi et al. (Reference Shi, Luan, Song and Zhang2020)]. A recent study on BDNF in MDD has shown that novel biomarker directions could yield promising results. Preliminarily, Lin and colleagues demonstrate that a novel-specific mature BDNF (mBDNF) enzyme-linked immunosorbent assay (ELISA) kit yields good diagnostic sensitivity and specificity for both MDD and BD (Lin et al., Reference Lin, Fu, Zhou, Liu, Bobrovskaya and Zhou2021). Ergo, combination with methylation and/or polymorphism status could increase power to differentiate of pre-existing peripheral BDNF assessment approaches (Halaris et al., Reference Sharma, Meresh, Ghanshyam, Kang, Hage, Garlenski and Sinacore2015; Zhao et al., Reference Zhao, Zhang, Chen, Su, Zhou, Wang, Xia, Huang, Wang, Hu, Cao, Guo, Yuan, Wang, Yi, Lu, Wu, Wu, Hong, Peng and Fang2017; Lin et al., Reference Lin, Fu, Zhou, Liu, Bobrovskaya and Zhou2021). To the best of our knowledge, there have only been some additional relevant reviews and meta-analyses on the influence of BDNF-related mutations in MDD (Yan et al., Reference Yan, Wang, Kuang, Xu, Li, Chen and Yang2014; Colle et al., Reference Colle, Deflesselle, Martin, David, Hardy, Taranu, Falissard, Verstuyft and Corruble2015a; Zhao et al., Reference Zhao, Chen, Yang, Han, Fang, Qiu, Yang, Qiao, Ma, Wang, Jiang, Song, Zhou, Zhang, Chen, Qi, Yang and Pan2018; Kim et al., Reference Kim, Ham and Han2019), but none that includes all the aspects evaluated to date. Colle et al. (Reference Colle, Deflesselle, Martin, David, Hardy, Taranu, Falissard, Verstuyft and Corruble2015a) looked at the influence of BDNF + BDNF receptor polymorphisms on antidepressant efficacy in MDD. They found 5 GWAS and 30 association studies, most evaluated the Val66Met SNP. Therein, the presence of a Met carrier state was associated with a higher antidepressant efficacy, but only in Asian patients, partially confirming previous ethnicity-stratified studies and warranting further research into this direction. An earlier meta-analysis confirmed the effects of the BDNF Val66Met polymorphism on SSRI response in Asians (Yan et al., Reference Yan, Wang, Kuang, Xu, Li, Chen and Yang2014). Hence, ethnicity stratifications are of utmost importance when considering further trials on BDNF-related mutations in MDD, especially when looking at clinical response. In this case, precedent data on population-related specificities of BDNF polymorphisms should be taken into account (Cargill et al., Reference Cargill, Altshuler, Ireland, Sklar, Ardlie, Patil, Lane, Lim, Kalyanaraman, Nemesh, Ziaugra, Friedland, Rolfe, Warrington, Lipshutz, Daley and Lander1999; Tsai et al., Reference Tsai, Cheng, Yu, Chen and Hong2003; Itoh et al., Reference Itoh, Hashimoto, Kumakiri, Shimizu and Iyo2004; Shimizu et al., Reference Shimizu, Hashimoto and Iyo2004; Choi et al., Reference Choi, Kang, Lim, Oh and Lee2006; Pivac et al., Reference Pivac, Kim, Nedić, Joo, Kozarić-Kovačić, Hong and Muck-Seler2009; Yeebo, Reference Yeebo2015). Pharmacological action of antidepressant drugs may result in differing treatment (Gratacòs et al., Reference Gratacòs, Soria, Urretavizcaya, González, Crespo, Bayés, de Cid, Menchón, Vallejo and Estivill2008) and side effects (Zou et al., Reference Zou, Wang, Liu, Feng, Wang, Zang, Yu, Wei, Liu, Liu, Tao, Li, Li, Hu, Li, Zhang, Ye and Xu2010a) due to BDNF polymorphisms. Ketamine as a novel, rapidly acting glutamatergic antidepressant, seems to work better in Val than Met carriers acutely (Laje et al., Reference Laje, Lally, Mathews, Brutsche, Chemerinski, Akula, Kelmendi, Simen, McMahon, Sanacora and Zarate2012; Chen et al., Reference Chen, Lin, Wu, Cheng, Li, Hong, Tu, Bai, Tsai and Su2019, Reference Chen, Kao, Tsai, Li, Lin, Hong, Bai, Tu and Su2021a, 2021b); however, Met carriers show similar effects after repeated drug exposure (Salvadore et al., Reference Salvadore, Singh, Xi, Kolb, Favis, Van Nueten and Drevets2015).
Commonly used SSRIs could be more efficacious in Asian Met carriers (Choi et al., Reference Choi, Kang, Lim, Oh and Lee2006; Xu et al., Reference Xu, Lin, Rao, Dang, Ouyang, Mai and Zhang2012; Yan et al., Reference Yan, Wang, Kuang, Xu, Li, Chen and Yang2014; Wang et al., Reference Wang, Xu, Chen, Xia and Liu2014; Colle et al., Reference Colle, Deflesselle, Martin, David, Hardy, Taranu, Falissard, Verstuyft and Corruble2015a; Ramesh et al., Reference Ramesh, Venkatesan, Chellathai and Silamban2021), though having higher efficacy in Caucasian Val/Val homozygotes (Colle et al., Reference Colle, Gressier, Verstuyft, Deflesselle, Lépine, Ferreri, Hardy, Guilloux, Petit, Fève, Falissard, Becquemont and Corruble2015b). On the contrary, SNRIs and TCAs seem less efficacious in Caucasian Val/Val homozygotes (Colle et al., Reference Colle, Gressier, Verstuyft, Deflesselle, Lépine, Ferreri, Hardy, Guilloux, Petit, Fève, Falissard, Becquemont and Corruble2015b), and hence antidepressant class could potentially be selected according to ethnicity and Val66Met genotype, though there are some contradictory results (Tsai et al., Reference Tsai, Cheng, Yu, Chen and Hong2003; Rajewska-Rager et al., Reference Rajewska-Rager, Skibińska, Szczepankiewicz, Kapelski, Dmitrzak-Weglarz, Leszczyńska-Rodziewicz and Hauser2008; Alexopoulos et al., Reference Alexopoulos, Glatt, Hoptman, Kanellopoulos, Murphy, Kelly, Morimoto, Lim and Gunning2010; Chi et al., Reference Chi, Chang, Lee, Lee, Gean, Yang, Lu and Chen2010). Indeed, treatment effect correlations are most well elucidated in Asian populations (Yan et al., Reference Yan, Wang, Kuang, Xu, Li, Chen and Yang2014; Colle et al., Reference Colle, Deflesselle, Martin, David, Hardy, Taranu, Falissard, Verstuyft and Corruble2015a; Kang et al., Reference Kang, Bae, Kim, Shin, Hong, Ahn, Jeong, Yoon and Kim2016). Interestingly, treatment effect–genotype interactions also appear as correlations with a distinct influence on peripheral BDNF levels by different chemical antidepressants (Matrisciano et al., Reference Matrisciano, Bonaccorso, Ricciardi, Scaccianoce, Panaccione, Wang, Ruberto, Tatarelli, Nicoletti, Girardi and Shelton2009; Deuschle et al., Reference Deuschle, Gilles, Scharnholz, Lederbogen, Lang and Hellweg2012), though ECT and exercise do not seem to have such an effect (Krogh et al., Reference Krogh, Rostrup, Thomsen, Elfving, Videbech and Nordentoft2014; Ryan et al., Reference Ryan, Dunne and McLoughlin2018).
The presence of the Met allele significantly modulated the association between stress and depression in further trials (Hosang et al., Reference Hosang, Shiles, Tansey, McGuffin and Uher2014; Bîlc et al., Reference Bîlc, Vulturar, Chiş, Buciuman, Nuţu, Bunea, Szentágotai-Tătar and Miu2018), specifically life stress. Less so, but childhood adversity appears to modulate these associations as well (Hosang et al., Reference Hosang, Shiles, Tansey, McGuffin and Uher2014; Rahman et al., Reference Rahman, Millischer, Zeebari, Forsell and Lavebratt2017; Bîlc et al., Reference Bîlc, Vulturar, Chiş, Buciuman, Nuţu, Bunea, Szentágotai-Tătar and Miu2018) and interactions of Val66Met with resilience may play a role therein (Peters et al., Reference Peters, Xavier, Mondin, Cardoso, Ferreira, Teixeira, Gräeff, Quevedo, Jansen, Souza, Oses, Pinheiro, da Silva and Ghisleni2021). Here, correlations with the hypothalamic–pituitary–adrenal (HPA) axis (Schule et al., Reference Schule, Zill, Baghai, Eser, Zwanzger, Wenig, Rupprecht and Bondy2006) and neuroinflammation-related pathways (Anisman and Hayley, Reference Anisman and Hayley2012; Dowlati et al., Reference Dowlati, Herrmann, Swardfager, Liu, Sham, Reim and Lanctôt2010; Liu et al., Reference Liu, Ho and Mak2012), which are a potentially major factor in MDD pathogenesis relating to stress (van Praag, Reference van Praag2004; Troubat et al., Reference Troubat, Barone, Leman, Desmidt, Cressant, Atanasova, Brizard, El Hage, Surget, Belzung and Camus2021), have to be considered. These strongly imply alterations of hippocampal neuromorphology in MDD (Hariri et al., Reference Hariri, Goldberg, Mattay, Kolachana, Callicott, Egan and Weinberger2003; Videbech, Reference Videbech2004; Neto et al., Reference Neto, Borges, Torres-Sanchez, Mico and Berrocoso2011), though hippocampal alterations solely due to Val66Met seem increasingly unlikely (Harrisberger et al., Reference Harrisberger, Spalek, Smieskova, Schmidt, Coynel, Milnik, Fastenrath, Freytag, Gschwind, Walter, Vogel, Bendfeldt, de Quervain, Papassotiropoulos and Borgwardt2014, Reference Harrisberger, Smieskova, Schmidt, Lenz, Walter, Wittfeld, Grabe, Lang, Fusar-Poli and Borgwardt2015). Further evidence on neuromorphological changes in the PFC, hippocampus, amygdala and specifically the rostral ACC (Kim et al., Reference Kim, Ham and Han2019; Shen et al., Reference Shen, Lu, Jiang, Ye, Zhou, He, Li, Xu and Cheng2020) in MDD, affected by BDNF-related mutations (including BDNF Val66Met) (Youssef et al., Reference Youssef, Underwood, Huang, Hsiung, Liu, Simpson, Bakalian, Rosoklija, Dwork, Arango and Mann2018), strengthen the need for meta-analysis. Larger trials and correlation of neuroimaging and neurogenetic data with specific patient and phenotypical factors are required (Legge et al., Reference Legge, Sendi, Cole, Cohen-Woods, Costafreda, Simmons, Farmer, Aitchison, McGuffin and Fu2015; Kim et al., Reference Kim, Ham and Han2019; Shen et al., Reference Shen, Lu, Jiang, Ye, Zhou, He, Li, Xu and Cheng2020).
In late-onset depression, Met carrier state of Val66Met may contribute to the development of depression (Hwang et al., Reference Hwang, Tsai, Hong, Yang, Lirng and Yang2006; Czira et al., Reference Czira, Wersching, Baune and Berger2012) or Val homozygosity delay or prevent the onset of geriatric depression (Kanellopoulos et al., Reference Kanellopoulos, Gunning, Morimoto, Hoptman, Murphy, Kelly, Glatt, Lim and Alexopoulos2011). Potential other factors specific to the elderly may be at play, hence implicating probable age-related specificities, especially since BDNF-related SNPs seem to play little role in adolescent, young adult (here Chinese Han) patients (Zhang et al., Reference Zhang, Ran, Ai, Wang, Chen, Wu, Liu, Jin, Wang and Kuang2020a). Furthermore, gender-stratified analyses hint towards increased gender-specific relevance of BDNF Val66Met in men with MDD (Verhagen et al., Reference Verhagen, van der Meij, van Deurzen, Janzing, Arias-Vásquez, Buitelaar and Franke2010; Suchanek et al., Reference Suchanek, Owczarek, Kowalczyk, Kucia and Kowalski2011; Czira et al., Reference Czira, Wersching, Baune and Berger2012).
This study has several limitations. For one, PubMed and Embase were the only databases which were searched. Although PubMed and Embase are comprehensive and widely used, including the vast majority of papers, some studies (e.g. Cochrane database or ScienceDirect) might not have been identified by our search strategy. Secondly, some studies were excluded based on depressive episodes appearing in the context of BD. While we considered this a necessary measure, polarity of depression did not affect associations of BDNF with depression in some studies (Ryan et al., Reference Ryan, Dunne and McLoughlin2018) and BD patients were in the minority in most mixed MDD/BD trials (Sarchiapone et al., Reference Sarchiapone, Carli, Roy, Iacoviello, Cuomo, Latella, di Giannantonio, Janiri, de Gaetano and Janal2008; Hennings et al., Reference Hennings, Kohli, Uhr, Holsboer, Ising and Lucae2019). Alas, bipolar depression may share similar BDNF-related characteristics with MDD. However, comparative reviews on the influence of BDNF polymorphisms in MDD versus BD might bring valuable information regarding separate modes of pathogenesis, diagnosis (Zhao et al., Reference Zhao, Zhang, Chen, Su, Zhou, Wang, Xia, Huang, Wang, Hu, Cao, Guo, Yuan, Wang, Yi, Lu, Wu, Wu, Hong, Peng and Fang2017) and neuromorphology changes, especially with respect to manic or psychotic symptoms and severity of depression (Post, Reference Post2007; Pae et al., Reference Pae, Chiesa, Porcelli, Han, Patkar, Lee, Park, Serretti and De Ronchi2012). Post-mortem data hints towards both shared and differential BDNF cascade protein and volumetric changes in MDD versus BD in limbic structures (Dunham et al., Reference Dunham, Deakin, Miyajima, Payton and Toro2009; Cao et al., Reference Cao, Bauer, Sharma, Mwangi, Frazier, Lavagnino, Zunta-Soares, Walss-Bass, Glahn, Kapczinski, Nielsen and Soares2016). Also, increased BDNF promotor I methylation appears to differentiate MDD against both BD and healthy controls (Carlberg et al., Reference Carlberg, Scheibelreiter, Hassler, Schloegelhofer, Schmoeger, Ludwig, Kasper, Aschauer, Egger and Schosser2014) and furthermore be affected by antidepressant therapy in MDD (Kim et al., Reference Kim, Cho, Jang, Shin, Park, Lee, Cho, Choi and Lee2010; Carlberg et al., Reference Carlberg, Scheibelreiter, Hassler, Schloegelhofer, Schmoeger, Ludwig, Kasper, Aschauer, Egger and Schosser2014). Apart from associations with diagnosis (Fuchikami et al., Reference Fuchikami, Morinobu, Segawa, Okamoto, Yamawaki, Ozaki, Inoue, Kusumi, Koyama, Tsuchiyama and Terao2011; Song et al., Reference Song, Miyaki, Suzuki, Sasaki, Tsutsumi, Kawakami, Shimazu, Takahashi, Inoue, Kan, Kurioka and Shimbo2014; Januar et al., Reference Januar, Ancelin, Ritchie, Saffery and Ryan2015; Hsieh et al., Reference Hsieh, Lin, Lee and Huang2019; Ferrer et al., Reference Ferrer, Labad, Salvat-Pujol, Barrachina, Costas, Urretavizcaya, de Arriba-Arnau, Crespo, Soriano-Mas, Carracedo, Menchón and Soria2019) and disease risk (Kim et al., Reference Kim, Stewart, Kang, Bae, Kim, Shin, Hong, Ahn, Jeong and Yoon2015), promotor methylation status could influence neurocognitive performance and interact with biological sex and childhood trauma in MDD (Ferrer et al., Reference Ferrer, Labad, Salvat-Pujol, Barrachina, Costas, Urretavizcaya, de Arriba-Arnau, Crespo, Soriano-Mas, Carracedo, Menchón and Soria2019).
To summarise, BDNF polymorphisms most probably exert manifold, phenotypical neurobiological changes on a molecular level, depending on the respective individual genetic and epigenetic make-up, gene–gene interactions (GxGs) and adversity-associated gene–environment interactions (GxEs), for example, due to stress, ELA, etc. (van Velzen et al., Reference van Velzen, Schmaal, Jansen, Milaneschi, Opmeer, Elzinga, van der Wee, Veltman and Penninx2016; Bîlc et al., Reference Bîlc, Vulturar, Chiş, Buciuman, Nuţu, Bunea, Szentágotai-Tătar and Miu2018; Zhao et al, Reference Zhao, Chen, Yang, Han, Fang, Qiu, Yang, Qiao, Ma, Wang, Jiang, Song, Zhou, Zhang, Chen, Qi, Yang and Pan2018). Pathomechanistically, MDD endophenotype could be influenced by reciprocally interactive BDNF-related SNPs (Zhang et al., Reference Zhang, Yang, Xu, Sun, Yang, Liu, Xu and Shen2010) predisposing to neuromorphological changes (of e.g. classically implicated limbic structures) (Hajek et al., Reference Hajek, Kopecek and Höschl2012) by means of altered signalling cascades (Zhang et al., Reference Zhang, Yang, Xu, Sun, Yang, Liu, Xu and Shen2010; Juhasz et al., Reference Juhasz, Dunham, McKie, Thomas, Downey, Chase, Lloyd-Williams, Toth, Platt, Mekli, Payton, Elliott, Williams, Anderson and Deakin2011; Pulay and Réthelyi, Reference Pulay and Réthelyi2016). These changes in turn could contribute to depressive symptomatology, by for example, cognitive alterations (Juhasz et al., Reference Juhasz, Dunham, McKie, Thomas, Downey, Chase, Lloyd-Williams, Toth, Platt, Mekli, Payton, Elliott, Williams, Anderson and Deakin2011), resulting in phenotypically manifest MDD. Such data can add to an explanation of MDD as a ‘major depressive spectrum disorder’ (Angst and Merikangas, Reference Angst and Merikangas1997; Caruso et al., Reference Caruso, Nanni, Riba, Sabato, Mitchell, Croce and Grassi2017), rather than a homogenous psychiatric disorder. This is in line with clinical experience in psychiatry and accumulating understanding of many common psychiatric diseases as spectrum disorders (Angst and Merikangas, Reference Angst and Merikangas1997; Akiskal and Benazzi, Reference Akiskal and Benazzi2006; Benazzi, Reference Benazzi2006; Berrocal et al., Reference Berrocal, Ruiz Moreno, Rando, Benvenuti and Cassano2008). While a BDNF-related biomarker approach for MDD seems feasible, it most likely will have to combine serum, genetic, and potentially, neuroimaging markers. Importantly, further research into extra-promotor regions of the BDNF gene (Hing et al., Reference Hing, Sathyaputri and Potash2018) and detailed elucidation of uncertainties regarding genotypic specificities of BDNF genetic variants in different populations (Hong et al., Reference Hong, Huo, Yen, Tung, Pan and Tsai2003; Ferreira Fratelli et al., Reference Ferreira Fratelli, Willatan Siqueira, Rodrigues Gontijo, de Lima Santos, de Souza Silva and Rodrigues da Silva2021) and relating to neuromorphology (Devlin et al., Reference Devlin, Cao and Stanfill2021) are required to improve knowledge of potential genetic candidates for such an endeavour. Here, deep sequencing approaches and subsequent analysis could aid significantly, as already suggested elsewhere (Licinio et al., Reference Licinio, Dong and Wong2009).
Conclusions
Although no overall association of BDNF-related mutations with MDD can be sustained, based on currently available evidence, future research should elucidate specific BDNF-MDD associations based on demographic, clinico-phenotypical and neuromorphological variables. The evidence-related inconsistencies are likely explained by relevance of specific aspects versus overall association of BDNF-related polymorphisms in MDD and genetic specificities differing amongst populations, implicating pre-described stratification effects. Additionally, biomarker approaches, especially combinatory paradigms (utilising genetic, serum and neuroimaging markers) involving BDNF seem promising and should be further investigated.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/neu.2022.22
Author contributions
JH and BN conceived the study protocol, including search strategy and inclusion and exclusion criteria. JH performed the literature search, data extraction, and data interpretation and wrote the first draft. BN supervised study eligibility and data extraction and edited the final draft. Both authors reviewed and approved the final version of the manuscript.
Financial support
This research received no specific grant from any funding agency, commercial or not-for-profit sectors.
Statement of interest
None.