



Significant outcomes
-
Acute and subchronic administration of selective serotonin reuptake inhibitors, fluoxetine and escitalopram, failed to promote antidepressant-like effects in WHR submitted to the FST with and without pre-test session.
-
Subchronic imipramine and acute S-ketamine induced antidepressant-like effects in WHR submitted to the FST without a pre-test session.
-
Repeated treatment with fluoxetine and imipramine failed to promote antidepressant-like effects in WHR submitted to the learned helplessness model.
Limitations
-
We did not investigate the effects of the drugs when injected via different routes of administration and variable time-schedule to assess possible pharmacokinetic changes in WHR.
-
We did not test the effect of combining different treatments to investigate if there would be any possible facilitation of the antidepressant effect, as observed in patients showing resistance to monotherapy.
-
We did not investigate the behaviour of female WHR or compared this strain with other Wistar rats.
Introduction
Science has been facing a "reproducibility crisis" in the last few years, consisting of difficulties in replicating clinical and preclinical studies (Begley & Ioannidis, Reference Begley and Ioannidis2015; Hunter, Reference Hunter2017; Fanelli, Reference Fanelli2018). Many factors are being seen as causal, such as the absence of internal and external validity, lack of complete dataset information, and under-powered designs (Olorisade et al., Reference Olorisade, Brereton and Andras2017; Fanelli, Reference Fanelli2018; Pound and Ritskes-Hoitinga, Reference Pound and Ritskes-Hoitinga2018), as well as poor reporting of study results (Avey et al., 2016). Besides that, in preclinical studies, other factors seem to correlate with the lack of reliability, such as the strain, sex and origin of the animals, the genotype, the season of the year and time of day when experiments were performed, the cage density, among others (Chesler et al., Reference Chesler, Wilson, Lariviere, Rodriguez-Zas and Mogil2002; Tillmann, Reference Tillmann2017; Einat et al., Reference Einat, Ezer, Kara and Belzung2018). Therefore, it is fundamentally essential to in-depth characterise the behaviour of a given strain in different animal models, try to reduce the reproducibility deficit and comply with the 3R principles (reduction, replacement and refinement) (Russel & Burch, Reference Russel and Burch1959).
Outbred animals are commonly used in the studies to evaluate or modulate the human population for presenting a genetic variability, being widely used in the investigation of depression neurobiology and treatment (Chia et al., Reference Chia, Achilli, Festing and Fisher2005; Lohmiller & Swing, Reference Lohmiller and Swing2006). The Wistar Hannover rats (WHR), also known as Wistar Han or Han Wistar, were originated in 1964, at Hannover Institute (Germany), after selected breeding from "regular" Wistar rats based on specific phenotypic characteristics, which have not been described in details (Giknis & Clifford, Reference Giknis and Clifford2009). These animals are more commonly used for toxicity studies but are also used in oncology studies, safety and efficacy tests, and a few behavioural studies (Rockett et al., Reference Rockett, Narotsky, Thompson, Thillainadarajah, Blystone, Goetz, Ren, Best, Murrell, Nichols, Schmid, Wolf and Dix2006; Hayakawa et al., Reference Hayakawa, Mimura, Tachibana, Furuya, Kodama, Aoki, Hosokawa, Fukui, Shibata, Yoshida, Masuyama, Narita, Kuwagata, Hisada and Maki2013; Noritake et al., Reference Noritake, Ikeda, Ito, Miwa and Senuma2013; Charles River Laboratories International, 2015; Soares-Cunha et al., Reference Soares-Cunha, Coimbra, Borges, Domingues, Silva, Sousa and Rodrigues2018; Gauvin et al., Reference Gauvin, Dalton, Harter, Holdsworth, May, Tapp, Zimmermann, Kilgus and Baird2019). When exposed to chronic mild stress (CMS), WHR displayed reduced body weight and locomotor activity in the open field test (OFT), increased grooming, and reduced sucrose consumption, characterising an anhedonia-like behaviour (Theilmann et al., Reference Theilmann, Kleimann, Rhein, Bleich, Frieling, Löscher and Brandt2016). In the FST, Kawai and colleagues (2018) showed that both acute and chronic imipramine (30 mg/kg) induced an antidepressant-like effect, although with varying responses depending on the injection time. Other studies have used the WHR in antidepressant research without characterising their response to different antidepressant drugs (Kawai et al., Reference Kawai, Kodaira, Tanaka, Ishibashi, Kudo, Kawashima and Mitsumoto2018).
Given the current establishment of the WHR colony in our campus and the lack of detailed information regarding their response to antidepressants, we evaluated the behavioural and the antidepressant response of WHR in two widely used animal models of depression: the forced swim test (FST) and the learned helplessness paradigm (LH). Both FST and LH are extensively used in the screening of new antidepressant drugs and investigation of their mechanisms of action (Cryan et al., Reference Cryan, Markou and Lucki2002; Pryce et al., Reference Pryce, Azzinnari, Spinelli, Seifritz, Tegethoff and Meinlschmidt2011; Slattery & Cryan, Reference Slattery and Cryan2014; Kara et al., Reference Kara, Stukalin and Einat2018; Ribeiro et al., Reference Ribeiro, Casarotto, Staquini, Silva, Biojone, Wegener and Joca2019). Porsolt et al. (Reference Porsolt, Le and Jalfre1977) originally described that the FST is based on the animal’s pre-exposure to a stressful event, the forced swim (pre-test). In the test session, usually performed 24 h after the pre-test, the animal initially presents intensive struggling to escape from stress, followed by an immobile behaviour, which is reduced by acute/subacute treatment (3× in 24 h) with different classes of antidepressant drugs (Porsolt et al., Reference Porsolt, Le and Jalfre1977, Reference Porsolt, Bertin and Jalfre1978b; Cryan & Lucki, Reference Cryan and Lucki2000). The FST has low cost, easy implementation and relatively good reproducibility. Therefore, it is one of the most widely used behavioural readouts in the study of depression neurobiology and searches for new treatments (Cryan et al., Reference Cryan, Markou and Lucki2002, Reference Cryan, Valentino and Lucki2005; Abelaira et al., Reference Abelaira, Réus and Quevedo2013; Slattery & Cryan, Reference Slattery and Cryan2014; Wang et al., Reference Wang, Timberlake, Prall and Dwivedi2017).
In the LH, the rodents are subjected to inescapable and unpredictable shocks that lead to cognitive deficits characterised by an increased number of escape failures presented in during the subsequent session with escapable aversive stimuli (test) (Joca et al., Reference Joca, Padovan and Guimarães2003, Reference Joca, Zanelati and Guimarães2006; Pryce et al., Reference Pryce, Azzinnari, Spinelli, Seifritz, Tegethoff and Meinlschmidt2011; Maier & Seligman, Reference Maier and Seligman2016). This model’s relevance is due to its good face and construct validity (Vollmayr & Gass, Reference Vollmayr and Gass2013). Exposure to inescapable aversive stimuli leads to behavioural and pathophysiological changes that resemble those observed in patients with depression (Pryce et al., Reference Pryce, Azzinnari, Spinelli, Seifritz, Tegethoff and Meinlschmidt2011; Ribeiro et al., Reference Ribeiro, Casarotto, Staquini, Silva, Biojone, Wegener and Joca2019). In addition, LH meets the predictive validity criteria since helplessness behaviour can be reversed by chronic treatment with antidepressants, while treatment with anxiolytics, neuroleptics, stimulants or depressants of the central nervous system does not affect behaviour in this test (Sherman et al., Reference Sherman, Sacquitne and Petty1982).
Differences in strains of mice and rats can be a source of considerable variability in the FST results (Porsolt et al., Reference Porsolt, Bertin and Jalfre1978b; Lahmanme & Armario, Reference Lahmanme and Armario1996; Lucki et al., Reference Lucki, Dalvi and Mayorga2001; Will et al., Reference Will, Aird and Redei2003; Jin et al., Reference Jin, Chen, Ran, Li, Xiong, Zheng, Gao and Li2017). For instance, Will et al. (Reference Will, Aird and Redei2003) demonstrated that Wistar-Kyoto (WKY) rats are more immobile than Wistar, Fisher 344 and Sprague-Dawley (SD) rats in FST (Will et al., Reference Will, Aird and Redei2003). The discrepant behaviour of different mice strains on FST can modify the response of these animals to the different antidepressant classes (Porsolt et al., Reference Porsolt, Bertin and Jalfre1978b; Lucki et al., Reference Lucki, Dalvi and Mayorga2001; Jin et al., Reference Jin, Chen, Ran, Li, Xiong, Zheng, Gao and Li2017). Moreover, some rat strains present ‘endogenous phenotypes’ associated with depression or resistance to treatment, such as the Flinders Sensitive Line (FSL) and the WKY rats, respectively (Paré, Reference Paré1994; Lahmanme & Armario, Reference Lahmanme and Armario1996; Overstreet et al., Reference Overstreet, Friedman, Mathé and Yadid2005; Overstreet & Wegener, Reference Overstreet and Wegener2013). In such cases, pre-exposure to the pre-test session is not necessary to reveal the antidepressant effect of drugs (Lahmame et al., Reference Lahmame, Del, Pazos, Yritia and Armario1997; Overstreet & Wegener, Reference Overstreet and Wegener2013). Although there is less evidence than with the FST, the strain also seems to influence behaviour in the LH. Wieland et al. (Reference Wieland, Boren, Consroe and Martin1986) demonstrated that the inbred WKY strain and SD rats from Charles River Holtzman are more susceptible to LH training than other strains or suppliers, which may influence both strain and suppliers helplessness behaviour.
In light of insufficient evidence regarding the response of WHR to antidepressants and the increasing use of such strain in psychopharmacology studies, we aimed to characterise the behaviour of WHR in two different protocols of FST and LH. For FST, we used a similar version of Porsolt’s protocol, with pre-test (15 min) and test (5 min), with 24-h interval (Porsolt et al., Reference Porsolt, Le and Jalfre1977; Detke et al., Reference Detke, Rickels and Lucki1995), and a single session of FST (10 min, no pre-swim, as previously described for identifying the behavioural effect of antidepressant drugs in animals with increased vulnerability to stress, such as the FSL animals) (Schiller et al., Reference Schiller, Pucilowski, Wienicke and Overstreet1992a; Pucilowski & Overstreet, Reference Pucilowski and Overstreet1993; Overstreet et al., Reference Overstreet, Friedman, Mathé and Yadid2005). Concerning the LH, we used: 1. a traditional protocol consisting of two sessions: pre-test (inescapable shocks for 40 min) and test (30 escapable shocks and preceded by a warning tone) (Joca et al., Reference Joca, Padovan and Guimarães2003, Reference Joca, Zanelati and Guimarães2006; Stanquini et al., Reference Stanquini, Biojone, Guimarães and Joca2017); 2. an adapted protocol with three sessions: pre-test (inescapable shocks for 40 min), test 1 and test 2 (both with 30 escapable shocks preceded by a warning tone), which considers the animals’ response pattern to stress (Reinés et al., Reference Reinés, Cereseto, Ferrero, Sifonios, Podestá and Wikinski2008; Macedo et al., Reference Macedo, Cladouchos, Sifonios, Cassaneli and Wikinski2013). Moreover, we investigated the response of WHR to antidepressants of different pharmacological classes after single and repeated treatment.
Experimental procedures
Animals and housing
Male WHR [Crl:WI(Han)] with 6–7 weeks (200–250 g), purchased from the local farm facility of the University of São Paulo, Campus of Ribeirão Preto, were used. The animals were brought to the animal house of the Laboratory of Pharmacology (School of Pharmaceutical Sciences) and remained undisturbed for at least 1 week before the start of the experiments. Animals were housed in a controlled temperature room (24 ± 1°C), under a 12-h light/dark cycles (lights on at 6 a.m.) in pairs in acrylic boxes (20 × 30 × 13 cm) with wood bedding (sawdust), no environmental enrichment, with ad libitum access to food (commercial rodent chow, Nuvilab – Quimtia – Paraná, Brazil) and tap water. In the LH, after the pre-test session, the rats were kept isolated in individual acrylic boxes (20 × 30 × 13 cm). The bedding was changed three times per week. All the efforts were made to reduce the suffering of the animals during the experiments, and all the procedures were conducted in conformity with the National Council for the Control of Animal Experimentation (CONCEA), complying with international laws and politics. The local Ethics Committee (University of São Paulo, Campus of Ribeirão Preto) approved all the experimental protocols (19.1.582.60.3 and 19.1.248.60.6). The total number of animals used in the present study was 305.
Drugs
The following drugs were as follows: imipramine (Abcam, USA or Sigma Aldrich, #I7379), a tricyclic antidepressant, at the doses 5, 10, 15 and 20 mg/kg; fluoxetine (Prati-Donaduzzi, BR), a selective serotonin reuptake inhibitor (SSRI), at the doses of 5, 10 and 20 mg/kg; escitalopram (Prati-Donaduzzi, BR), an SSRI, at the doses of 3, 10 and 20 mg/kg; and s-ketamine (Cristália, BR), a dissociative anaesthetic with fast-acting antidepressant effects (Berman et al., Reference Berman, Cappiello, Anand, Oren, Heninger, Charney and Krystal2000; Murrough et al., Reference Murrough, Iosifescu, Chang, Al Jurdi, Green, Perez, Pillemer, Foulkes and Mathew2013), at the dose of 10 mg/kg. All drugs were freshly diluted in saline 0,9%, excepted fluoxetine, which was dissolved on tween 80 and 90% saline 0,9%. The drugs and their respective vehicle were systemically administered (i.p.) in a volume of 1 ml/kg. The doses used were based on previously published studies and preliminary data from our group (Porsolt et al., Reference Porsolt, Anton, Blavet and Jalfre1978a; Zazpe et al., Reference Zazpe, Artaiz, Labeaga, Lucero and Orjales2007; Reinés et al., Reference Reinés, Cereseto, Ferrero, Sifonios, Podestá and Wikinski2008; France et al., Reference France, Li, Owens, Koek, Toney and Daws2009; Macedo et al., Reference Macedo, Cladouchos, Sifonios, Cassaneli and Wikinski2013; Fernández-guasti et al., Reference Fernández-guasti, Olivares-nazario, Reyes and Martínez-mota2017; Kawai et al., Reference Kawai, Kodaira, Tanaka, Ishibashi, Kudo, Kawashima and Mitsumoto2018).
Behavioural experiments
Animals were brought to the experimental room for acclimatisation 30 min before the start of the experiments. All experiments were performed between 8 a.m. and 2 p.m for FST and between 8 a.m. and 4 p.m for LH. All rooms had standard conditions of illumination (lights on from 6 a.m. to 6 p.m.), ventilation and temperature (23 ± 1C) throughout the experiment. The animals were in blocks randomly assigned to the different experimental conditions. Similarly, the animals were also randomly assigned to testing to avoid potential circadian bias.
To characterise the behaviour and response to antidepressant drugs, independent groups of WHR were submitted to two different protocols of FST and LH.
FST: (a) with pre-swimming session 24 h before the test (Fig. 1a); (b) without pre-swimming (Fig. 2a). All FST experiments were recorded using a Sony video camera (model DCR-SR47), and data were analysed by an experimenter blind to treatment conditions.

Fig. 1. Effect of imipramine, fluoxetine and escitalopram in WHR submitted to forced swim test with pre-swim (b, d) and open field test (OFT) (c, e). (a) Experimental design of forced swim test with pre-swim: the animals were submitted to the pre-test (PT) section of forced swim and, immediately after, received the first i.p. injection (0 h), the second and third injection were administrated 5 h and 23 h after PT, respectively. One hour after the last injection, the WHR were submitted to OFT and FST. The bars represent mean ± standard error (one-way ANOVA followed by Dunnett’s post hoc test or Student’s t-test, *p < 0.05 compared to the vehicle group (Veh), n = 6–10 per group).
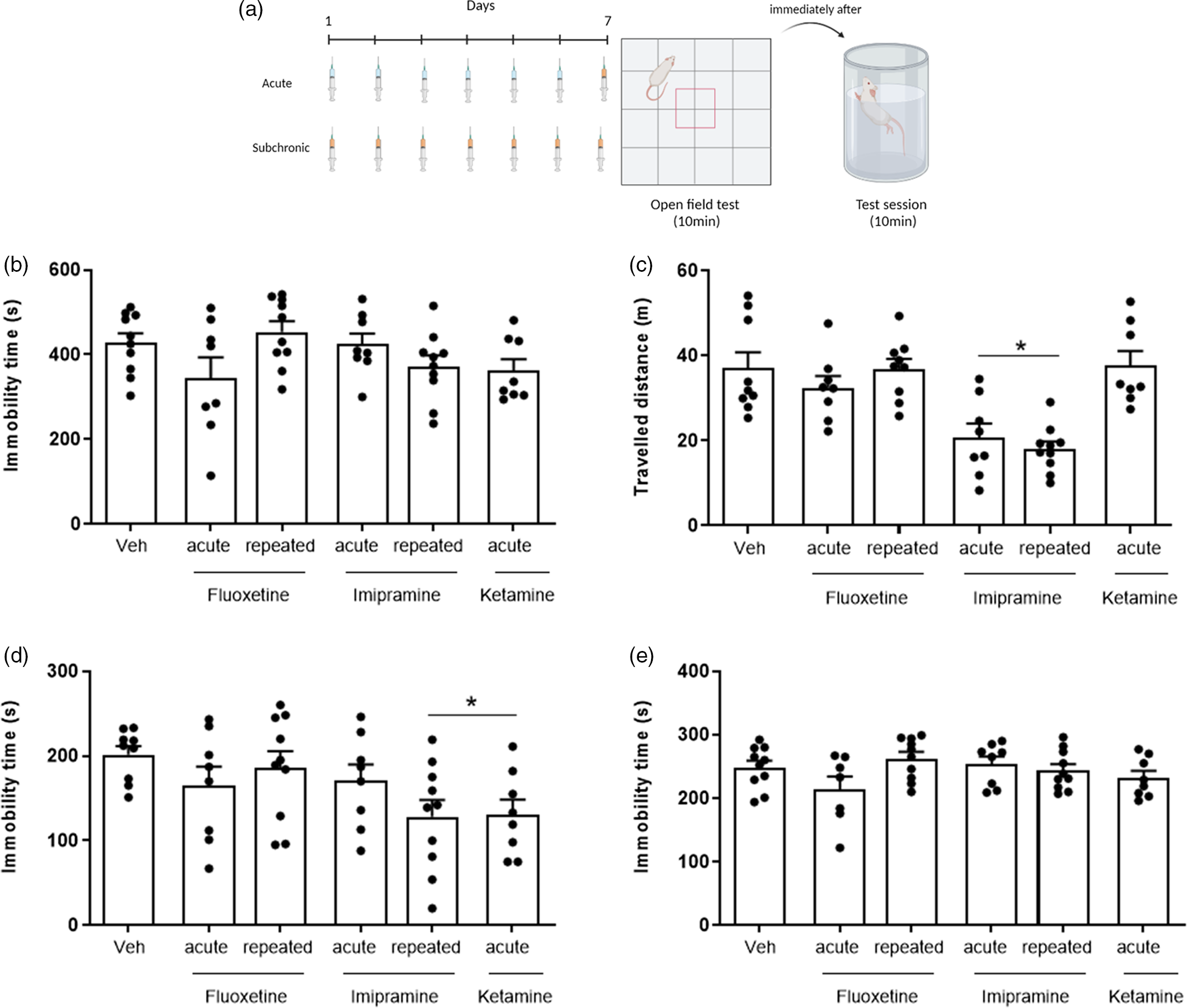
Fig. 2. Effects of the acute and repeated administration of fluoxetine (20 mg/kg), imipramine (20 mg/kg) and s-ketamine (10 mg/kg) in rats submitted to forced swimming test (FST) without pre-swim and open field test (OFT). (a) Experimental design of forced swim test without pre-swim: the animals were treated systematically during 7 days with antidepressant or vehicle (subchronic treatment) or received the vehicle for 6 days and received the drugs only on the last day (acute treatment). On the seventh day, the OFT and FST were performed 1 h after the last treatment. (b) Immobility time during 10 min of FST; (c) locomotor activity during 10 min of OFT; (d) immobility time in the first 5 min of FST; and (e) immobility time in the last 5 min of FST. The bars represent mean ± standard error (one-way ANOVA followed by Dunnett’s post hoc test, *p < 0.05 compared to the vehicle group (Veh), n = 8–10 per group).
LH: (a) with pre-test and test, described previously by Joca et al. (Reference Joca, Padovan and Guimarães2003, Reference Joca, Zanelati and Guimarães2006) and Stanquini et al. (Reference Stanquini, Biojone, Guimarães and Joca2017) (Fig. 4a); (b) with three sessions, pre-test, test 1 and test 2, (Figs. 5a and 6a). All parameters analysed (number of failures, escapes, avoidances or intertrial crossings) were recorded by Esquiva Ativa software, which also controls the shocks release, intervals and sounds during the sessions.

Fig. 3. Effects of the acute and repeated administration of escitalopram (20 mg/kg) and s-ketamine (10 mg/kg) in rats submitted to forced swim test (FST) without pre-swim and open field test (OFT). (a) Experimental design of FST without pre-swim; (b) immobility time during 10 min of FST; (c) locomotor activity during 10 min of OFT; (d) immobility time in the first 5 min of FST; and (e) immobility time in the last 5 min of FST. The bars represent mean ± standard error (one-way ANOVA followed by Dunnett’s post hoc test, *p < 0.05 compared to the vehicle group (Veh), n = 9–10 per group).

Fig. 4. Effects of fluoxetine (20 mg/kg) at number of failures, avoidances, escapes and intertrial crossings of Wistar Hannover rats subjected to learned helplessness (traditional protocol). (a) Experimental design used for LH traditional protocol. The animals were divided into non-stressed and stressed. On the first day of the experiment (pre-test), the non-stressed animals were just exposed to the shuttle box for 40 min without shocks, while the stressed ones were subjected to 40 inescapable shocks (0.6 mA, 10 s duration, 30–90 s interval). The animals were treated once daily for 7 days with vehicle (sterile saline) or fluoxetine 20 mg/kg/mL and 1 h after the last injection was tested with 30 escape shocks (0.6 mA, 10 s duration, 30–90 s interval) preceded by a warning tone (60 dB, 670 Hz, 5 s). (b) Number of failures. (c) Number of avoidance. (d) Number of escape. (e) Number of intertrial crossings. NST-VEH (non-stressed vehicle), ST-VEH (stressed-vehicle), ST-FLX (stressed-fluoxetine 20 mg/kg/mL). Values are expressed as mean ± SEM. Kruskal–Wallis test followed by Dunn’s test indicated that there was no statistical difference between the groups.

Fig. 5. Effects of fluoxetine (20 mg/kg) at number of failures, avoidances, escapes and intertrial crossings of Wistar Hannover rats subjected to learned helplessness (adapted protocol). (a) Experimental design used for adapted LH protocol. The animals were divided into non-stressed and stressed. On the first day of the experiment (pre-test), the non-stressed animals were just exposed to the shuttle box for 40 min without shocks, while the stressed ones were subjected to 40 inescapable shocks (0.6 mA, 10 s duration, 30–90 s interval). Twenty-four hours later, the stressed animals were subjected to test 1 (T1) (0.6 mA, 10 s duration, 30–90 s interval) and classified according to their number of failures in resilient (0–10 failures) or susceptible (15–30 failures). The non-stressed group was again just exposed to the shuttle box without shocks. The animals groups were treated once daily for 7 days with: non-stressed (sterile saline); stressed-resilient (sterile saline), stressed-susceptible-vehicle (sterile saline) and stressed-susceptible-fluoxetine (20 mg/kg/mL). One hour after the last administration, the animals were exposed to test 2 (0.6 mA, 10 s duration, 30–90 s interval) preceded by a warning tone (60 dB, 670 Hz, 5 s). (b) Number of failures after T1. (c) Number of avoidance after T1. (d) Number of escapes after T1. (e) Number of intertrial crossings after T1. Data are expressed as mean ± SEM. The Mann–Whitney test indicated a significant difference in the number of failures, avoidances and escapes between resilient (n = 6) and susceptible (n = 22) animals. *p < 0.05. (f) Number of failures after T2. (g) Number of avoidance after T2. (h) Number of escapes after T2. (i) Number of intertrial crossings after T2. NST-VEH (non-stressed vehicle), ST-RESIL (stressed-resilient), ST-SUSC-VEH (stressed-susceptible-vehicle), ST-SUSC FLX (stressed-susceptible-fluoxetine). Data are expressed as mean ± SEM. The Kruskal–Wallis test followed by Dunn’s test indicated that there was no significant difference in the number of failures between the ST-SUSC VEH × ST-SUSC FLX groups. *p < 0.05. n = 6–11 per group.

Fig. 6. Effects of imipramine (20 mg/kg) at number of failures, avoidances, escapes and intertrial crossings of Wistar Hannover rats subjected to learned helplessness (adapted protocol). (a) Experimental design used for adapted LH protocol. The animals were divided into non-stressed and stressed. On the first day of the experiment (pre-test), the non-stressed animals were just exposed to the shuttle box for 40 min without shocks, while the stressed ones were subjected to 40 inescapable shocks (0.6 mA, 10 s duration, 30–90 s interval). Twenty-four hours later, the stressed animals were subjected to test 1 (T1) (0.6 mA, 10 s duration, 30–90 s interval) and classified according to their number of failures in resilient (0–10 failures) or susceptible (15–30 failures). The non-stressed group was again just exposed to the shuttle box without shocks. The animals groups were treated once daily for 7 days with: non-stressed (sterile saline); stressed-resilient (sterile saline), stressed-susceptible-vehicle (sterile saline) and stressed-susceptible-fluoxetine (20 mg/kg/mL). One hour after the last administration, the animals were exposed to test 2 (0.6 mA, 10 s duration, 30–90 s interval) preceded by a warning tone (60 dB, 670 Hz, 5 s). (b) Number of failures after T1. (c) Number of avoidance after T1. (d) Number of escapes after T1. (e) Number of intertrial crossings after T1. Data are expressed as mean ± SEM. The Mann– Whitney test indicated a significant difference in the number of failures, avoidances, escapes and crossings between resilient (n = 19) and susceptible (n = 29) animals. *p < 0.05. (f) Number of failures after T2. (g) Number of avoidance after T2. (h) Number of escape after T2. (i) Number of intertrial crossings after T2. NST-VEH (non-stressed vehicle), ST-RESIL (stressed-resilient), ST-SUSC-VEH (stressed-susceptible-vehicle), ST-SUSC IMIP (stressed-susceptible-imipramine). Data are expressed as mean ± SEM. The Kruskal–Wallis test followed by Dunn’s test indicated that there was no significant difference in the number of failures between the ST-SUSC VEH × ST-SUSC IMIP groups. *p < 0.05. n = 10–19 per group.
(a) FST with pre-swim. It was performed according to a modified Porsolt’s protocol (Porsolt et al., Reference Porsolt, Le and Jalfre1977; Detke et al., Reference Detke, Rickels and Lucki1995; Cryan et al., Reference Cryan, Valentino and Lucki2005), with adaptations described in Diniz et al. (Reference Diniz, Casarotto, Fred, Biojone, Castrén and Joca2018). Briefly, the protocol consisted of placing the animals to individually swim in a plastic cylinder (30 cm in diameter, 40 cm in height, containing 30 cm of water, with a temperature of 24 ± 1°C, for 15 min – pre-test, PT). After this period, the animals were removed and placed separately to dry before returning to their original boxes. Twenty-four hours later, the animals were submitted to the OFT (see below) and, immediately after, to a 5-min forced swimming session (test, T). The total immobility time was analysed by an experimenter unaware of the treatments. The animal was considered immobile when it remained still performing only small movements necessary for flotation. In the test session, the water was changed after each animal to avoid the influence of possible alarm substances. The treatment protocol consisted of three injections in an interval of 24 h: the first injection was administered immediately after PT, the second at the end of the day (5 p.m.) and the third injection 1 h before the OFT.
(b) FST without pre-swim. This protocol was adapted from the protocol used for testing the effect of antidepressants in FSL rats, characterised by higher vulnerability to stress, and no need to be exposed to pre-test to reveal the antidepressant effect of drugs (Schiller et al., Reference Schiller, Pucilowski, Wienicke and Overstreet1992a; Overstreet et al., Reference Overstreet, Friedman, Mathé and Yadid2005; Sales et al., Reference Sales, Fogaça, Sartim, Pereira, Wegener, Guimarães and Joca2018). The apparatus and conditions were the same described above; however, rats were submitted to a single session of OFT and FST, with 10 min of duration each. The water was changed after each test. The animals were treated daily for 7 days, and the test was performed 1 h after the last injection. The acutely treated group received the drug only on the last day, 1 h before the OFT, and vehicle on the other 6 days; the repeatedly treated group received the drugs (imipramine, fluoxetine or escitalopram) or vehicle during the 7 days of treatment.
Open field test (OFT). Before the FST, the rats were submitted to the OFT to investigate any possible influence of the drugs on locomotor activity. The WHR were placed individually into a wood square arena (with dimensions 72 × 72 × 40 cm high) for 5 or 10 min before FST with and without pre-swim, respectively. The locomotor activity was recorded and evaluated using AnyMaze software (Stoelting, United Kingdom).
(c) Learned helplessness (LH, protocol 1). The experiments were carried out in a shuttle box (37 × 33 × 54 cm, manufactured by Insight), with a metal grid on the floor, which delivers foot shocks. This apparatus consists of two compartments of equal sizes separated by a wall with a central opening through which the animals could cross from one compartment to another (Joca et al., Reference Joca, Padovan and Guimarães2003, Reference Joca, Zanelati and Guimarães2006; Ribeiro et al., Reference Ribeiro, Ferraz-de-Paula, Pinheiro and Palermo-Neto2009; Stanquini et al., Reference Stanquini, Biojone, Guimarães and Joca2017). The WHR were exposed to a pre-test (PT) session consisting of 40 inescapable foot shocks (0.6 mA, 10 s duration, 30–90 s interval) and then treated daily for 7 days with fluoxetine or vehicle until the test (T) that was performed 1 h after the last injection. At this session, the rats were submitted to 30 escapable foot shocks (0.4 mA, 10 s, 30–90 s interval), preceded (5 s) by a warning tone (60 dB, 670 Hz). In the test session, animals could avoid the shocks by crossing from one compartment to the other during tone presentation or escape by interrupting the shock when crossing the shuttle box during shock presentation. An escape failure was defined as the absence of avoidance or escape (Ribeiro et al., Reference Ribeiro, Ferraz-de-Paula, Pinheiro and Palermo-Neto2009; Stanquini et al., Reference Stanquini, Biojone, Guimarães and Joca2017). Repeated antidepressant treatments, such as fluoxetine, decrease the number of escape failures (Sherman et al., Reference Sherman, Sacquitne and Petty1982; Reinés et al., Reference Reinés, Cereseto, Ferrero, Sifonios, Podestá and Wikinski2008; Macedo et al., Reference Macedo, Cladouchos, Sifonios, Cassaneli and Wikinski2013). Therefore, escape failures were used in the present work as a parameter indicative of LH behaviour. The number of intertrial crossings was used as a parameter indicative of locomotor activity (Geoffroy & Christensen, Reference Geoffroy and Christensen1993). The animals were divided into two groups: stressed and non-stressed. The stressed group was submitted to both PT and T sessions. The non-stressed group was exposed to the shuttle box during the PT session without shocks, while the T session was conducted as usual.
(d) Learned helplessness (LH, protocol 2). This protocol was developed considering that, like humans, rats show significant heterogeneity in their response to stress and may be susceptible or resilient (resistant) (Reinés et al., Reference Reinés, Cereseto, Ferrero, Bonavita and Wikinski2004; Russo et al., Reference Russo, Murrough, Han, Charney and Nestler2012; Drugan et al., Reference Drugan, Christianson, Warner and Kent2013; Wood & Bhatnagar, Reference Wood and Bhatnagar2015; Febbraro et al., Reference Febbraro, Svenningsen, Tran and Wiborg2017). Rats were submitted to the same apparatus and conditions described above; however, they were submitted to three sessions, PT, T1 and T2. First, they were exposed to a pre-test that consisted of 40 inescapable foot shocks (0.6 mA, 10 s duration, 30–90 s interval). In this session, the non-stressed group was exposed to the shuttle box without shocks. Twenty-four hours later, the animals were subjected to T1 and, after this session, classified according to the number of failures: resilient (0–10 failures) or susceptible (15–30 failures). Only the susceptible animals were treated daily for 7 days with the antidepressants (fluoxetine or imipramine) until T2, which was performed 1 h after the last injection. At T1 and T2, 30 shocks (0.6 mA) were delivered, with the same duration and intervals as the PT session, but the shocks were escapable and preceded by a warning tone (60 dB, 670 Hz, 5 s) (Joca et al., Reference Joca, Padovan and Guimarães2003, Reference Joca, Zanelati and Guimarães2006; Reinés et al., Reference Reinés, Cereseto, Ferrero, Sifonios, Podestá and Wikinski2008; Macedo et al., Reference Macedo, Cladouchos, Sifonios, Cassaneli and Wikinski2013; Stanquini et al., Reference Stanquini, Biojone, Guimarães and Joca2017; Ribeiro et al., Reference Ribeiro, Casarotto, Staquini, Silva, Biojone, Wegener and Joca2019). The escape failures number was used as a parameter indicative of LH behaviour, and the number of intertrial crossings was used as a parameter of locomotor activity (Geoffroy & Christensen, Reference Geoffroy and Christensen1993).
Experimental design
Experiment 1: Effects of imipramine, fluoxetine and escitalopram in WHR submitted to the OFT and FST with pre-swim
This experiment aimed to investigate the behavioural response of WHR to antidepressants of different pharmacological classes in the FST under standardised experimental conditions. The animals received three systemic injections (0, 5 h and 23 h after PT) of imipramine (5, 10, 15 and 20 mg/kg), fluoxetine (3 and 10 mg/kg), escitalopram (3 and 10 mg/kg) or vehicle (1 mL/kg). The animals were submitted to the OFT 1 h after the last injection, followed by the FST immediately after.
Experiment 2: Effects of imipramine, fluoxetine, escitalopram and s-ketamine in WHR submitted to the OFT and FST without pre-swim
Some rat strains are more vulnerable to stress and do not require pre-exposure to forced swim to reveal increased immobility, such as the FSL rats, and repeated antidepressant treatment is often required to attenuate stress effects (Schiller et al., Reference Schiller, Pucilowski, Wienicke and Overstreet1992b; Pucilowski & Overstreet, Reference Pucilowski and Overstreet1993; Overstreet et al., Reference Overstreet, Friedman, Mathé and Yadid2005; Overstreet & Wegener, Reference Overstreet and Wegener2013; Schiller et al., Reference Schiller, Pucilowski, Wienicke and Overstreet1992a; Pucilowski & Overstreet, Reference Pucilowski and Overstreet1993; Overstreet et al., Reference Overstreet, Friedman, Mathé and Yadid2005; Overstreet & Wegener, Reference Overstreet and Wegener2013). Therefore, we aimed at investigating the behavioural response of WHR to different antidepressants in this protocol. Moreover, since s-ketamine is known to promote a rapid antidepressant effect in protocols that typically requires chronic antidepressant administration, we also investigated the effects induced by a single s-ketamine administration (10 mg/kg).
Independent groups of WHR received one daily injection of imipramine (20 mg/kg/day), fluoxetine (20 mg/kg/day) or vehicle for 7 days (repeated/subchronic group) and were submitted to the OFT and the FST 1 h after the last injection (seventh day). To control for the effects of the single administration, independent groups of WHR received vehicle injection for 6 days and received a single injection of imipramine (20 mg/kg/day), fluoxetine (20 mg/kg/day) or vehicle on the seventh day (acute group). To investigate the rapid effect of s-ketamine, an independent group was treated with the vehicle for 6 days and a single s-ketamine administration on the seventh day, 1 h before the FST.
The animals were randomly assigned to the different treatment conditions, constituting of a block of six groups: vehicle, (vehicle from day 0 to day 7); repeated imipramine (20 mg/kg/day); acute imipramine group (20 mg/kg); repeated fluoxetine group (20 mg/kg/day); acute fluoxetine group (20 mg/kg); and s-ketamine group (10 mg/kg). To investigate the response of WHR to another SSRI, in addition to fluoxetine, we investigated the effects induced by acute and repeated treatment with escitalopram, using the same protocol as described above. Independent groups of WHR were randomised in four experimental groups: acute escitalopram group (10 mg/kg); repeated escitalopram group (10 mg/kg/day); and the vehicle and acute s-ketamine group (10 mg/kg). During the 6 days, the animals were treated from 11 a.m. to 1 p.m., and on day 7, the animals received the injection 1 h before the OFT.
Experiment 3: Effects of fluoxetine in WHR submitted to LH (protocol 1)
The animals were divided into three groups and received the following intraperitoneal treatments for 7 days: non-stressed or habituated: vehicle (sterile isotonic saline, 1 mL/kg); stressed: vehicle (sterile isotonic saline, 1 mL/kg); and stressed: fluoxetine 20 mg/kg/mL dissolved in tween 80 at 2% in sterile isotonic saline. During the 6 days, the animals were treated around 12 a.m and on day 7, 1 h before the test (Joca et al., Reference Joca, Padovan and Guimarães2003, Reference Joca, Zanelati and Guimarães2006; Stanquini et al., Reference Stanquini, Biojone, Guimarães and Joca2017; Ribeiro et al., Reference Ribeiro, Casarotto, Staquini, Silva, Biojone, Wegener and Joca2019).
Experiments 4 and 5: Effects of fluoxetine in WHR submitted to LH (protocol 2)
On the first day of the experiment, the animals were divided into habituated or stressed and subjected to pre-test. The next day, the animals were subjected to T1. In both PT and T1, the habituated group was exposed to the shuttle box without shocks. After T1, the animals were classified as resilient or susceptible and divided into four groups to receive the following treatments via intraperitoneal for 7 days: non-stressed or habituated: vehicle (sterile isotonic saline, 1 mL/kg); resilient-stressed: vehicle (sterile isotonic saline, 1 mL/kg); susceptible-stressed: vehicle (sterile isotonic saline, 1 mL/kg); and susceptible-stressed: fluoxetine 20 mg/kg/mL dissolved in tween 80 at 2% in sterile isotonic saline. The animals were treated around 12 a.m. and 1 h before the test on day 7 for 6 days. In another experiment with the same conditions, we evaluated the effects of imipramine, thus replacing the group treated with fluoxetine previously described.
Statistical Analyses
The statistical analyses were performed using the GraphPad Prism program (version 8.3.1, GraphPad Software, San Diego California, USA). The effect of stress in the vehicle groups, stressed and not stressed, was analysed by Student’s t-test in both OFT and FST. The effect of drugs in the FST was analysed by one-way ANOVA followed by Dunnett’s post-test. In both cases, differences with p < 0.05 were considered significant. For the first LH protocol, the behaviour parameters were analysed by Kruskal–Wallis test followed by Dunn’s test. For the second LH protocol, the behaviour parameters were analysed by the Mann–Whitney test to compare resilient × susceptible animals and the Kruskal–Wallis test, followed by Dunn’s test for T2. In both cases, differences with p < 0.05 were considered significant.
Results
Experiment 1: Imipramine, fluoxetine and escitalopram fail to induce antidepressant-like effects in WHR submitted to the FST with pre-swim (Fig. 1)
Imipramine treatment did not induce significant changes in the immobility time when compared to the vehicle group (n = 7–10, F 4,36 = 1.041, p = 0.3997; Fig. 1b). However, the doses of 10, 15 and 20 mg/kg significantly reduced the distance travelled in the OFT (n = 7-10, F 4,36 = 3.205, p = 0.0238; Fig. 1c). Moreover, neither fluoxetine nor escitalopram changed the immobility time in the FST (F 4,33 = 0.8726, p = 0.4908; Fig. 1d). The distance travelled in the OFT was also not changed by any of the treatments (F 4,32 = 1.195, p = 0.3318; Fig. 1e).
Experiment 2: Imipramine, fluoxetine, escitalopram and s-ketamine effects in WHR submitted to the FST without pre-swim (Figs. 2 and 3)
Since the conventional treatment with different antidepressants (0, 5 and 23 h after PT) was ineffective in the standard FST protocol, we tested the effect of the single and repeated treatments with antidepressants in WHR submitted to an adapted FST protocol, characterised by a single section of forced swimming (10 min). The immobility time was analysed for 10 min or blocks of 5 min.
In 10 min of FST, none of the treatments induced significant changes in the immobility time (n = 8–10, F 5,48 = 2.141, p = 0.0765; Fig. 2b). However, when analysing the immobility time in blocks of 5 min, both imipramine (7 days) and acute s-ketamine significantly reduced the immobility time in the first 5 min (n = 8–10, F 5,47 = 2.551, p = 0.0402; Fig. 2d), an effect that was not observed in the last 5 min (n = 7–10, F 5,47 = 1.859, p = 0.1197; Fig. 2e). In the OFT, acute and repeated imipramine treatment reduced the locomotor activity compared to the vehicle group during the 10 min of analysis (n = 8–10, F 5,47 = 9.746, p < 0.0001; Fig. 2c).
Since fluoxetine failed to change the immobility time, we investigated whether WHR would not respond to another SSRI, escitalopram (Fig. 3). S-ketamine was used as a positive control in these experiments. The acute administration of s-ketamine significantly reduced the immobility time in 10 min (n = 9–10, F 3,34 = 3.495, p = 0.0259; Fig. 3b) and in the first 5 min of the FST (n = 9–10, F 3,34 = 6.527, p = 0.0013; Fig. 3d), but not in the last 5 min (n = 9–10, F 3,34 = 0.3356, p = 0.7997; Fig. 3e). In contrast, the acute and repeated administration of escitalopram did not change immobility time in any of the analyses. None of the drugs changed the locomotor activity in the OFT when compared with the control group (n = 9–10, F 3,34 = 1.301, p = 0.2901; Fig. 3c).
Experiment 3: Fluoxetine failed to induce antidepressant-like effects in WHR submitted to LH protocol 1 (Fig. 4) (n = 6–10)
Fluoxetine did not change the number of failures (H = 3.632, p = 0.1627, Kruskal–Wallis test), intertrial crossings (H = 4.738; p = 0.0936, Kruskal–Wallis test), avoidances (H = 3.275, p = 0.1945, Kruskal–Wallis test) and escapes (H = 2.797, p = 0.2469, Kruskal–Wallis test) presented by animals in the LH compared to vehicle-stressed group. Moreover, there was no significant difference between vehicle-stressed versus non-stressed vehicle groups.
Experiment 4: Fluoxetine failed to induce antidepressant-like effects in WHR submitted to LH protocol 2 (Fig. 5) (n = 6–11)
In the T1, there was a significant difference in the number of failures (U = 0.0, p < 0.0001), avoidances (U = 32.00, p = 0.0433) and escapes (U = 3.000, p < 0.0001) between resilient and susceptible animals (Fig. 5; Mann–Whitney). This effect does not seem to be associated with non-specific locomotor alterations, since there was no difference in the number of intertrial crossing (U = 34.50, p = 0.0692). In the T2, the repeated treatment of susceptible animals with fluoxetine 20 mg/kg did not induce an antidepressant effect when compared to the control group (stressed-susceptible-vehicle), characterised by the number of failures reduction (H = 19.68, p = 0.0002, Kruskal–Wallis test).
Experiment 5: Imipramine failed to induce antidepressant-like effects in WHR submitted to LH protocol 2 (Fig. 6) (n = 10–19)
In the T1, there was a significant difference in the number of failures (U = 0.0, p < 0.0001), avoidances (U = 92.00, p < 0.0001), escapes (U = 6.500, p < 0.0001) and intertrial crossings (U = 150.0, p = 0.0059) between resilient and susceptible animals (Fig. 6, Mann–Whitney test). In the T2, repeated treatment of susceptible animals with imipramine 20 mg/kg did not induce an antidepressant effect (stressed-susceptible-vehicle), characterised by the number of failures reduction (H = 26.86, p < 0.0001, Kruskal–Wallis test).
Discussion
Our work is the first to investigate the effect of different classes of antidepressant drugs in the WHR strain in two animal models of depression, FST and LH. Unexpectedly, the main finding in the present study is that subacute treatment with three different antidepressants (imipramine, fluoxetine and escitalopram, three injections during 24 h) was ineffective in WHR submitted to the traditional FST protocol (pre-test and test). Interestingly, in WHR not exposed to the pre-test, the repeated treatment with imipramine or acute S-ketamine administration induced antidepressant-like effects, but not fluoxetine or escitalopram treatment. Furthermore, in the LH model, both the subchronic treatment with fluoxetine or imipramine failed to modify the helplessness behaviour of WHR in two different protocols.
Considering that the WHR showed higher immobility during the test, compared with the outbred Wistar rats previously used in our lab (Sartim et al., Reference Sartim, Moreira and Joca2017; Diniz et al., Reference Diniz, Casarotto, Fred, Biojone, Castrén and Joca2018), we hypothesised that the lack of response to acute antidepressant administration in the FST could be related to an increased sensitivity of WHR to stress (average 200 s of immobility in a test lasting a total of 300 s). Therefore, we investigated the effect of acute and repeated treatment with antidepressants in WHR that had not been previously exposed to PT. Interestingly, the average immobility of WHR without pre-test was similar to the immobility reported in the literature for WKY and FSL rats, with an average of 200 s in the first swimming session (5 min) (Overstreet, Reference Overstreet1986; Lahmanme & Armario, Reference Lahmanme and Armario1996; Lahmame et al., Reference Lahmame, Del, Pazos, Yritia and Armario1997; Brand & Harvey, Reference Brand and Harvey2017; Sales et al., Reference Sales, Fogaça, Sartim, Pereira, Wegener, Guimarães and Joca2018). Moreover, our results revealed that only repeated imipramine treatment (7 days) and acute ketamine intervention reduced the immobility time in the WHR, whereas both fluoxetine and escitalopram were ineffective after either acute or repeated (7 days) treatment. This data is consistent with the observation that repeated treatment with tricyclics, monoamine oxidase inhibitors or 8-OH-DPAT is required to promote antidepressant effects in congenitally depressive-like rats, such as the FSL and the WKY rats (Overstreet, Reference Overstreet1986; Paré, Reference Paré1994; Marti & Armario, Reference Marti and Armario1996; Lahmame et al., Reference Lahmame, Del, Pazos, Yritia and Armario1997; Will et al., Reference Will, Aird and Redei2003; Overstreet & Wegener, Reference Overstreet and Wegener2013; Brand & Harvey, Reference Brand and Harvey2017), while fluoxetine is ineffective (López-Rubalcava & Lucki, Reference López-Rubalcava and Lucki2000; Will et al., Reference Will, Aird and Redei2003).
S-ketamine is an NMDA receptor antagonist that promotes acute and fast antidepressant effects; and it is effective in depressed patients that fail to respond to monoaminergic drugs (Berman et al., Reference Berman, Cappiello, Anand, Oren, Heninger, Charney and Krystal2000; Murrough et al., Reference Murrough, Iosifescu, Chang, Al Jurdi, Green, Perez, Pillemer, Foulkes and Mathew2013). Considering that acute ketamine intervention was effective in WHR, as described for FSL and WKY rats (Tizabi et al., Reference Tizabi, Bhatti, Manaye, Das and Akinfiresoye2012; Du Jardin et al., Reference Du Jardin, Liebenberg, Cajina, Müller, Elfving, Sanchez and Wegener2018; Sales et al., Reference Sales, Fogaça, Sartim, Pereira, Wegener, Guimarães and Joca2018), our results indicate that the WHR might be a strain with some level of resistance to monoaminergic drugs, especially SSRIs. Previous studies have reported mixed results regarding the effects of antidepressant treatment in WHR submitted to the FST, both in line and in contrast with our findings. For example, Kawai and colleagues (Kawai et al., Reference Kawai, Kodaira, Tanaka, Ishibashi, Kudo, Kawashima and Mitsumoto2018) demonstrated that both single and repeated injection of imipramine (30 mg/kg) promoted an antidepressant-like effect in WHR exposed to the FST. The discrepancy with our findings can result from several factors, ranging from differences in the protocol, apparatus and origin of animals. For instance, in the PT section, Kawai et al. (Reference Kawai, Kodaira, Tanaka, Ishibashi, Kudo, Kawashima and Mitsumoto2018) submitted the animals only to 10 min of swim FST and not 15 min as in the present study. Moreover, despite the temperature and depth of water being similar to ours, the diameter of the tank cylinder was smaller than ours (24 cm), which can also interfere with the behaviour of animals submitted to FST (Calil & Marcondes, Reference Calil and Marcondes2006; Bogdanovaa et al., Reference Bogdanovaa, Kanekara, D’Ancid and Renshawa2013). For example, the initial difficulty to detect the effect of SSRIs was overcome by changing the water depth and cylinder diameter (Detke et al., Reference Detke, Rickels and Lucki1995; Lucki, Reference Lucki1997).
The origin of the WHR might also play an important role in conflicting results between studies, since local colonies can be subject to founder effects and genetic drift, with substantial genetic divergence from other colonies of the same outbred strain (Brekke et al., Reference Brekke, Steele and Mulley2018). Kawai et al. (Reference Kawai, Kodaira, Tanaka, Ishibashi, Kudo, Kawashima and Mitsumoto2018) obtained the animals from CLEA laboratories (Japan), in contrast to our WHR which were purchased from Charles River Laboratories (USA – 2015) and then bred at the University of São Paulo. The WHR strain was originated in 1964, at Hannover Institute (ZentralInstitut für Versuchstierzucht), after selected breeding for certain phenotypes of Wistar rats(Giknis & Clifford, Reference Giknis and Clifford2009). In 1989, the Han Wistar rats were transferred to Biomedical Research Laboratories (BRL; later Research Consulting Company, RCC; Switzerland), posteriorly to GlaxoWellcome (Wood et al., Reference Wood, Nicolas, Choi, Roman, Nylander, Fernandez-Teruel, Kiianmaa, Bienkowski, de Jong, Colombo, Chastagnier, Wafford, Collingridge, Wildt, Conway-Campbell, Robinson and Lodge2017) and to Charles River, the UK, and North America in 1996 and 1997, respectively (Charles River Laboratories International, 2011, 2015). Furthermore, the WHR were also distributed directly from Hannover Institute to Javier labs, and from BRL to Taconic and CLEA laboratories (Wood et al., Reference Wood, Nicolas, Choi, Roman, Nylander, Fernandez-Teruel, Kiianmaa, Bienkowski, de Jong, Colombo, Chastagnier, Wafford, Collingridge, Wildt, Conway-Campbell, Robinson and Lodge2017; CLEA Japan, Inc., 2020). The behavioural differences in WHR according to their origins can result from the prevalence of specific mutations. For instance, Wood et al. (Reference Wood, Nicolas, Choi, Roman, Nylander, Fernandez-Teruel, Kiianmaa, Bienkowski, de Jong, Colombo, Chastagnier, Wafford, Collingridge, Wildt, Conway-Campbell, Robinson and Lodge2017) demonstrated that WHR presented more prevalence of the mutation at cysteine 407 on mGlu2, which is associated with altered emotionality and impulsivity than other Wistar strains. It was also demonstrated that the frequency of this mutation is not the same depending on the origin/vendor (Wood et al., Reference Wood, Nicolas, Choi, Roman, Nylander, Fernandez-Teruel, Kiianmaa, Bienkowski, de Jong, Colombo, Chastagnier, Wafford, Collingridge, Wildt, Conway-Campbell, Robinson and Lodge2017).
Accordingly, Theilmann et al. (Reference Theilmann, Kleimann, Rhein, Bleich, Frieling, Löscher and Brandt2016) demonstrated that, depending on the breeder, the WHR submitted to CMS could present different mRNA levels of S100 protein p11 (S100A10), a protein involved with the stress susceptibility. In male WHR from Javier labs, CMS did not change behaviour in the open field, elevated plus-maze or sucrose consumption tests. In contrast, WHR from Charles River showed higher anhedonia and anxiolytic-like phenotypes (Theilmann et al., Reference Theilmann, Kleimann, Rhein, Bleich, Frieling, Löscher and Brandt2016). Interestingly, females WHR from Charles River and Javier presented an anxiogenic behaviour when compared to females Wistar rats from Harlan-Winkelmann in the OFT. On elevated plus-maze, the females from Charles River lab presented low locomotor activity and more anxious behaviour than from Javier lab (Honndorf et al., Reference Honndorf, Lindemann, Töllner and Gernert2011). Other suppliers demonstrated different behaviour in the elevated plus-maze, light/dark emergence test, the object recognition memory, prepulse inhibition, and the ethanol intake and preference of male WHR (Goepfrich et al., Reference Goepfrich, Gluch, Friemel and Schneider2013; Momeni et al., Reference Momeni, Segerström and Roman2015). These differences are not exclusive for WHR, since similar behavioural differences have been reported for WKY and in SD depending on their origin (Paré & Kluczynski, Reference Paré and Kluczynski1997; Pecoraro et al., Reference Pecoraro, Ginsberg, Warne, Gomez, la and Dallman2006).
Different suppliers can also be a source of variability in detecting the effect of antidepressant drugs. Porsolt and colleagues (Porsolt et al., Reference Porsolt, Bertin and Jalfre1978b) demonstrated that three injections of imipramine promoted an antidepressant-like effect in Wistar rats from Charles River on FST; however, no effect was observed in the same strain from Iffa Credo. Similarly, Bupropion reduced the immobility time of WKY rats obtained from Charles River and Harlan laboratories, but not from Taconic (Browne et al., Reference Browne, van and Lucki2015). Altogether, differences in the strain, the origin and suppliers of the Wistar rats can interfere in behavioural responses to antidepressants in the FST, thus compromising the reproducibility of the data. Nevertheless, our results indicates that WHR purchased from Charles River Laboratories (USA – 2015) are more resistant to monoaminergic drugs when testesd in the FST, since several studies have described that these drugs reduce the immobility time of different rat strains when exposed to similar protocols, at similar doses (Porsolt et al., Reference Porsolt, Anton, Blavet and Jalfre1978a; Borsini et al., Reference Borsini, Lecci, Sessarego, Frassine and Meli1989; Slattery et al., Reference Slattery, Desrayaud and Cryan2005; France et al., Reference France, Li, Owens, Koek, Toney and Daws2009; Fernández-Guasti et al., Reference Fernández-guasti, Olivares-nazario, Reyes and Martínez-mota2017).
To further confirm our results, we used the LH model, which is one of the most relevant models for the study of depression neurobiology and new drugs with antidepressant activity (Pryce et al., Reference Pryce, Azzinnari, Spinelli, Seifritz, Tegethoff and Meinlschmidt2011; Ribeiro et al., Reference Ribeiro, Casarotto, Staquini, Silva, Biojone, Wegener and Joca2019). Among the neurochemical changes that lead to helplessness, those associated with the noradrenergic and serotoninergic systems stand out (Anisman & Sklar, Reference Anisman and Sklar1979; Sherman & Petty, Reference Sherman and Petty1980; Petty et al., Reference Petty, Kramer and Wilson1992). Regarding the latter, it has been suggested that LH is related to low serotonin levels in specific areas in the brain (Sherman & Petty, Reference Sherman and Petty1980; Petty et al., Reference Petty, Kramer and Wilson1992), which would be restored by repeated antidepressant treatment (Sherman et al., Reference Sherman, Sacquitne and Petty1982). Accordingly, we had previously demonstrated that 7 days treatment with desipramine, imipramine (15 mg/kg) and fluoxetine (20 mg/kg) were able to attenuate LH development in Wistar rats (Sales et al., Reference Sales, Maciel, Suavinha and Joca2021), exposed to a protocol similar to the one used in the present study (Stanquini et al., Reference Stanquini, Biojone, Guimarães and Joca2017; Ribeiro et al., Reference Ribeiro, Casarotto, Staquini, Silva, Biojone, Wegener and Joca2019). However, unlike other studies that reported the reversal of helplessness behaviour with fluoxetine (Zazpe et al., Reference Zazpe, Artaiz, Labeaga, Lucero and Orjales2007; Reinés et al., Reference Reinés, Cereseto, Ferrero, Sifonios, Podestá and Wikinski2008; Macedo et al., Reference Macedo, Cladouchos, Sifonios, Cassaneli and Wikinski2013), we failed to see any antidepressant effect in WHR submitted to the protocol 1.
In this protocol, we had previously demonstrated that the antidepressant effect in a different protocol, using only the susceptible animals, as previously described (Reinés et al., Reference Reinés, Cereseto, Ferrero, Bonavita and Wikinski2004; Russo et al., Reference Russo, Murrough, Han, Charney and Nestler2012; Drugan et al., Reference Drugan, Christianson, Warner and Kent2013; Wood & Bhatnagar, Reference Wood and Bhatnagar2015; Febbraro et al., Reference Febbraro, Svenningsen, Tran and Wiborg2017). The protocol’s change brings our experiment closer to the clinical setting, in which antidepressant pharmacological treatment is usually performed when behavioural/emotional impairment of individuals is detected (Macedo et al., Reference Macedo, Cladouchos, Sifonios, Cassaneli and Wikinski2013) and is supported by the literature (Reinés et al., Reference Reinés, Cereseto, Ferrero, Bonavita and Wikinski2004; Zazpe et al., Reference Zazpe, Artaiz, Labeaga, Lucero and Orjales2007; Reinés et al., Reference Reinés, Cereseto, Ferrero, Sifonios, Podestá and Wikinski2008). Stress-susceptible animals receiving repeated treatment with fluoxetine (20 mg/kg) did not show improved response when compared to the control group (stressed-susceptible-vehicle).
Previous studies also considered the vulnerability factor to stress when evaluating Wistar rats behaviour in the LH model. In their study, the rodents were exposed on day 1 to a training session (60 inescapable shocks, 0.6 mA, 15 s duration, for 1 h) and 4 days later, they were submitted to a test session (15 shocks, 20 s duration, for 15 min) (Reinés et al., Reference Reinés, Cereseto, Ferrero, Sifonios, Podestá and Wikinski2008). The animals were classified as helpless based on their latency to escape/avoid shocks in T1, which were treated with the drug of interest and re-exposed to the apparatus in test 2. Macedo et al. (Reference Macedo, Cladouchos, Sifonios, Cassaneli and Wikinski2013), Unlike these studies that discarded resilient animals in their analysis, this group was included in our study. Interestingly, we observed that these animals maintained their resilience behaviour at the end of the tests, demonstrating the effectiveness of our model.
In both studies mentioned above, intraperitoneal treatment with fluoxetine 10 mg/kg for 21 days was able to produce an antidepressant effect in Wistar rats submitted to LH, characterised by a decrease in latency time and the number of failures to escape or avoid the shocks (Reinés et al., Reference Reinés, Cereseto, Ferrero, Sifonios, Podestá and Wikinski2008; Macedo et al., Reference Macedo, Cladouchos, Sifonios, Cassaneli and Wikinski2013). In the present work, even with an adapted LH protocol, in which only stress-susceptible rats were treated with fluoxetine 20 mg/kg for 7 days, they continued not responding to the treatment. Once again, our results were contradictory to those in the literature, which could be related to protocol differences (number of shocks, intensity, duration, apparatus and days of treatment) but mainly to the strain used (Reinés et al., Reference Reinés, Cereseto, Ferrero, Bonavita and Wikinski2004, Reference Reinés, Cereseto, Ferrero, Sifonios, Podestá and Wikinski2008; Zazpe et al., Reference Zazpe, Artaiz, Labeaga, Lucero and Orjales2007; Macedo et al., Reference Macedo, Cladouchos, Sifonios, Cassaneli and Wikinski2013; Stanquini et al., Reference Stanquini, Biojone, Guimarães and Joca2017; Ribeiro et al., Reference Ribeiro, Casarotto, Staquini, Silva, Biojone, Wegener and Joca2019). In another attempt, we assessed whether the WHR would be responsive to treatment with imipramine 20 mg/kg for 7 days in the adapted LH protocol 2. Once again, there were two populations of animals, 39.5% of resilient and 60.5% of the susceptible. Similar to what we observed in the experiment with fluoxetine (adapted LH protocol), the resilient animals maintained their resilience until the end of the experiment. Regarding treatment, imipramine in WHR did not induce an antidepressant response. Contrasting with these data, previous works from our group demonstrated that intraperitoneal treatment with imipramine 15 mg/kg for 7 days induced an antidepressant effect in Wistar rats submitted to LH (Stanquini et al., Reference Stanquini, Biojone, Guimarães and Joca2017; Ribeiro et al., Reference Ribeiro, Casarotto, Staquini, Silva, Biojone, Wegener and Joca2019).
Compiling our data, the WHR did not respond to different antidepressants known to reverse helpless behaviour, even when tested in an adapted protocol to bring it closer to the clinical setting. Wistar rats comprise several strains derived from a common strain, namely Wistar, Wistar Hannover, Wistar Unielever, Wistar Kyoto and Wistar Furth (McCormick, Reference McCormick2017). Genetic drift and the fact that they grew up in different environments present different behavioural responses, making it difficult to compare studies with different strains (Palm et al., Reference Palm, Hävermark, Meyerson, Nylander and Roman2011). However, a fact that caught our attention is that WKY rats are a valid model of stress susceptibility, exhibiting behaviour, neurochemical and endocrine alterations parallel to clinical depression, being considered resistant to classical antidepressants (Aleksandrova et al., Reference Aleksandrova, Wang and Phillips2019). Corroborating these data, Willner et al. (Reference Willner, Gruca, Lason, Tota-Glowczyk, Litwa, Niemczyk and Papp2019) observed that after chronic stress, WKY did not respond to treatment with different classes of antidepressants (tricyclic, SSRI and serotonin and noradrenaline reuptake inhibitor) in the elevated plus-maze, object recognition test and sucrose intaket. Among the differences that could justify the resistance to antidepressant treatment in WKY are the greater hypothalamic–pituitary–adrenal axis responsiveness (Pardon et al., Reference Pardon, Gould, Garcia, Phillips, Cook, Miller, Mason and Morilak2002; Rittenhouse et al., Reference Rittenhouse, López-Rubalcava, Stanwood and Lucki2002; Morilak et al., Reference Morilak, Barrera, Echevarria, Garcia, Hernandez, Ma and Petre2005), abnormalities in monoaminergic transmission (Tejani-Butt et al., Reference Tejani-Butt, Paré and Yang1994; Durand et al., Reference Durand, Berton, Aguerre, Edno, Combourieu, Mormède and Chaouloff1999; Pollier et al., Reference Pollier, Sarre, Aguerre, Ebinger, Mormède, Michotte and Chaouloff2000; De La Garza & Mahoney, Reference La Garza and Mahoney2004), reduced hippocampus volume relative to Wistar (Tizabi et al., Reference Tizabi, Hauser, Tyler, Getachew, Madani, Sharma and Manaye2010), and lower levels of neurogenesis and cell proliferation in the hippocampus (Kin et al., Reference Kin, Yasuhara, Kameda, Agari, Sasaki, Morimoto, Okazaki, Umakoshi, Kuwahara, Kin, Tajiri and Date2017). Since the WHR is derived from a common lineage to WKY, we speculate that these animals may share similarities in their profile of resistance to antidepressant treatment.
Thus, the absence of acute response to antidepressants in WHR is similar to the delayed antidepressant response observed in patients. Moreover, the responsivity to the tricyclic antidepressant and to ketamine, but not to SSRI class, is another similarity with treatment-resistant patients (Oswald et al., Reference Oswald, Brezinova and Dunleavy1972; Posternak & Zimmerman, Reference Posternak and Zimmerman2005; Bennabi et al., Reference Bennabi, Charpeaud, Yrondi, Genty, Destouches, Lancrenon, Alaïli, Bellivier, Bougerol, Camus, Dorey, Doumy, Haesebaert, Holtzmann, Lançon, Lefebvre, Moliere, Nieto, Rabu, Richieri, Schmitt, Stephan, Vaiva, Walter, Leboyer, El-Hage, Llorca, Courtet and Aouizer2019). However, this conclusion is limited by the lack of investigation with other classes of antidepressants, such as monoamine oxidase inhibitors and serotonin and norepinephrine reuptake inhibitors, or even the combination of different classes of antidepressants. Moreover, we have not investigated more extended treatment periods,the drug effect in female rats and different drug administration routes. Although we have not compared the WHR with other rat strains or from different vendors, our results agree with Theilmann et al. (Reference Theilmann, Kleimann, Rhein, Bleich, Frieling, Löscher and Brandt2016), who demonstrated that WHR from Charles River are more susceptible to stress than WHR from other vendors. Therefore, further studies investigating how different antidepressants affect WHR behaviour in comparison with other rat strains (e.g. Wistar, SD and WKY), along with the evaluation of drug metabolism, neurochemical changes induced by stress, and genomic and proteomic profiles in the brain under stressful and non-stressful conditions, could clarify about the mechanisms subsiding the impaired response to antidepressants in WHR.
Acknowledgements
We are thankful to Flavia Salata and Miriam Melo for the helpful technical assistance.
KMS receives a PhD fellowship from CNPq (141900/2018-0), LV receives a PhD fellowship from FAPESP (2019/04616-2), SFL receives a Young Investigator fellowship from FAPESP (2017/19731-6) and SRLJ receives productivity a fellowship from CNPq (304780/20189). This study was financed in part by CAPES – the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brazil (Finance Code 001), FAPESP (2019/04616-2 and 2017/24304-0) and the Aarhus University Research Foundation.
The main idea of this work was from Samia Joca, Ariandra Sartim, Kennia Silveira and Letícia Vieira. Kennia Silveira, Ariandra Sartim, Letícia Vieira, Sabrina Lisboa, Samia Joca and Gregers Wegener designed this protocol together. Data collection has been done by Kennia Silveira, Ariandra Sartim and Letícia Vieira. Writing of the first drafts of the manuscript have been done by Kennia Silveira and Letícia Vieira. All authors contributed to and have approved the final manuscript.
Conflict of interest
Gregers Wegener is the Editor-in-Chief of Acta Neuropsychiatrica but was not involved and actively withdrew during the review and decision process of this manuscript.






