


INTRODUCTION
Yersiniosis is an important zoonotic disease in humans in Europe [1]. Most of the reported cases are caused by Y. enterocolitica. Human enteric yersiniosis is thought to be primarily foodborne [Reference Laukkanen-Ninios, Fredriksson-Ahomaa and Fraque2]. Y. enterocolitica has been shown to be transmitted mainly by pork products and Y. pseudotuberculosis by contaminated fresh produce. In a Y. pseudotuberculosis outbreak in Finland, it was likely that iceberg lettuce were contaminated by irrigation water contaminated with roe deer faeces [Reference Nuorti3]. In a small study conducted in Germany, raw game (including meat from roe deer, red deer, and chamois) were frequently (38%) contaminated with potentially pathogenic (ail-positive) Y. enterocolitica when studied by polymerase chain reaction (PCR) [Reference Bucher4].
Wild boars were recently shown to be an important reservoir of enteropathogenic Y. enterocolitica and Y. pseudotuberculosis in Switzerland [Reference Fredriksson-Ahomaa5]. Yersiniosis due to Y. pseudotuberculosis has also been shown to be a disease of major importance in deer [Reference Zhang6, Reference Sanford7]. Moreover, Y. pseudotuberculosis has also been reported to be a common finding in clinically healthy farmed deer weaners in New Zealand [Reference Hodges, Carman and Woods8].
The prevalence of Y. enterocolitica and Y. pseudotuberculosis in wild deer, however, has so far been very rarely studied [Reference Henderson9–Reference Aschfalk12]. In these few studies, both species were isolated from faecal samples of animals free from obvious symptoms of disease. However, all Y. enterocolitica strains were considered non-pathogenic, and Y. pseudotuberculosis was very rarely isolated from faecal samples. The aim of this work was to study the occurrence of Yersinia spp. in wild ruminants in Switzerland and to characterize the strains in order to obtain more information on the epidemiology of enteropathogenic Yersinia in the wildlife.
METHODS
Animals
This study was based on investigations carried out during 3 months (September–November) of the hunting season in 2011. The samples originated from shot red deer (Cervus elaphus), roe deer (Capreolus capreolus), chamois (Rupicapra rupicapra), and ibex (Capra ibex). The sampled animals were hunted in the central and eastern part of Switzerland. In total, 219 faecal samples (red deer, roe deer, chamois, ibex) were examined. The faecal samples originated from 77 red deer, 60 chamois, 55 roe deer and 27 alpine ibex. State gamekeepers and hunters collected the samples in the field immediately after shooting and evisceration of the wild ruminants. After opening the large intestine, faecal matter (at least 10 g) was collected from the colon, placed into sterile tubes and stored under refrigeration. For each hunted animal, sex, age, and location of hunting were recorded.
Yersinia detection and identification
About 1 g faecal material was mixed in 10 ml PMB [Reference Martínez13, Reference Fukushima, Gomyoda and Kaneko14]. After 2 weeks of cold enrichment at 4 °C, 10 μl of the enrichment was plated on cefsulodin-irgasan-novobiosin (CIN) agar (Oxoid AG, Switzerland). The CIN plates were incubated at 30 °C for 24–48 h. Presumptive positive colonies were subcultured on blood agar and then tested for the urease enzyme. Urease-positive colonies were identified with API 20E and matrix-assisted laser desorption/ionization–time of flight (MALDI–TOF) mass spectrometry [Reference Stephan15, Reference Fredriksson-Ahomaa16]. One isolate per sample in a total of 20 strains were biotyped and serotyped. The biotype was determined using pyrazinamidase and Tween activity, esculin hydrolysis, indole production, and salicin, xylose and trehalose fermentation and serotyping was performed with slide agglutination using commercial Y. enterocolitica O:1–O:3, O:5, and O:9 antisera (Denka Seiken, Japan).
Further strain characterization
Eight genes were studied by PCR: two virulence genes (yadA, virF) located on the virulence plasmid of the pathogenic Yersinia spp. (pYV) and five virulence genes (ail, ystA, ystB, myfA, hreP) and rfbC for O:3 serotype located in the chromosome [Reference Bhagat and Virdi17–Reference Weynants20]. The DNA was released from bacterial colonies by heating at 97 °C for 10 min, and 1 μl of this liquid was added to 19 μl of the mastermix (iQ™ SYBR Green Supermix; Bio-Rad, USA). The fluorescence intensity of SYBR Green and the melting curve analysis were studied using the CFX96 system (Bio-Rad). A threshold cycle (Ct) under 30 and a specific melting temperature (T m) indicated a positive result.
Antimicrobial susceptibility testing
Antimicrobial resistance analysis was performed by disk-diffusion test according to Clinical and Laboratory Standards Institute (CLSI, 2009). Fourteen antimicrobials were tested: ampicillin (10 μg), amoxicillin/clavulanic acid (20/10 μg), cefalothin (30 μg), cefoxitin (30 μg), cefpodoxim (10 μg), ceftazidim (30 μg), cefuroxime (30 μg), ciprofloxacin (5 μg), gentamicin (10 μg), kanamycin (30 μg), nalidixic acid (30 μg), streptomycin (10 μg), tetracycline (30 μg) and trimethoprim/sulfamethoxazole (1·25/23·75 μg) [Reference Fredriksson-Ahomaa16]. The reference strain Escherichia coli ATCC 25922 was used as the quality control.
RESUTS AND DISCUSSION
The occurrence of Yersinia spp. varied between 4% and 13% in wild ruminants being highest in roe (13%) and red deer (12%) (Table 1). The most frequently identified species was Y. enterocolitica found in 13%, 10%, 4% and 2% of roe deer, red deer, alpine ibex and chamois, respectively. Surprisingly, no Y. pseudotuberculosis was isolated even though cold enrichment in peptone broth supplemented with 1% mannitol and 0·15% bile salts (PMB), which should be favourable for Y. pseudotuberculosis [Reference Ortiz Martínez21], was used. The prevalence of Y. enterocolitica and Y. pseudotuberculosis in wild deer has so far very rarely been studied (Table 2). In Japan, 4% of the deer were shown to shed Y. pseudotuberculosis in faeces [Reference Fukushima and Gomyoda11]. In Norway, the prevalence of Yersinia in wild red deer was clearly lower [Reference Aschfalk12]. One reason for the higher prevalence of Yersinia in our study could be due to the use of a cold enrichment instead of 2 days enrichment at 21 °C. Y. enterocolitica was also the dominant species in Norwegian deer; however, one Y. pseudotuberculosis strain was detected in Norway. In Italy and New Zealand, the prevalence of Yersinia in red deer was clearly higher (Table 2). In the Italian study, most of the strains isolated were Y. kristensenii. One reason for the low isolation rate of Y. kristensenii in our study could be that we used CIN agar and Y. kristensenii grows very slowly. Y. enterocolitica was the dominant species in wild red deer in New Zealand; however, Y. frederiksenii was also frequently identified [Reference Henderson9]. In the same study, Y. pseudotuberculosis was sporadically isolated from clinically healthy farmed deer but not from wild deer. One reason for the low prevalence of Y. pseudotuberculosis could be that the carriage status cannot be adequately identified by faecal culture due to either sporadic shedding of this pathogen or due to the localization of this pathogen in the mesenteric or ileocecal lymph nodes [Reference Henderson9].
Table 1. Prevalence of Yersinia spp. in faeces of clinically healthy wild ruminants in Switzerland 2011

Table 2. Prevalence of Yersinia spp. in faeces of clinically healthy wild deer

The Yersinia spp. strains were identified with MALDI–TOF, API 20E and biotyped (Table 3). Only one of the 20 strains (strain no. 20) could not be identified at species level by MALDI–TOF. By API 20E this strain was identified as Y. frederiksenii/intermedia with an ID% of 98·5%. The biotype remained unknown for three Y. enterocolitica strains (strain nos. 15–17) by MALDI–TOF. One of the Y. enterocolitica strains (strain no. 17) was regarded as potentially pathogenic because it was pyrazinamidase, esculin and salicin negative. However, it was impossible to clearly differentiate if this strain belongs to biotype 3 or 5. This strain was xylose positive and trehalose negative. A typical strain of biotype 3 should be xylose and trehalose positive, and a typical biotype 5 strain should be xylose and trehalose negative [Reference Wauters, Kandolo and Janssens22]. This strain was also sorbitol negative. Y. enterocolitica strains are typically sorbitol positive and Y. pseudotuberculosis strains sorbitol negative.
Table 3. Identification and characterisation of the Yersinia strains isolated from wild ruminants free from obvious symptoms of disease
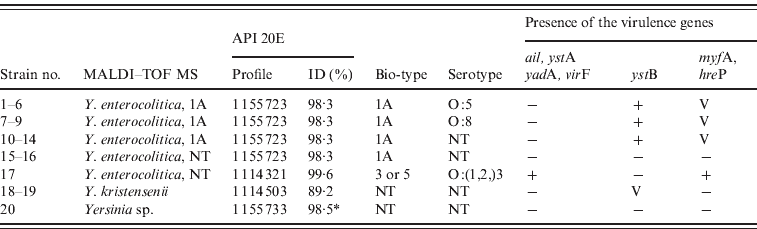
MALDI–TOF MS, Matrix-assisted laser desorption/ionization–time of flight mass spectrometry; NT, biotype not typable; V, the genes were detected in some strains.
* ID for Y. frederiksenii/intermedia.
Most (2/17) of the Y. enterocolitica strains from wild ruminants belonged to biotype 1A. The majority of the Y. enterocolitica strains isolated from food and the environment belong to this biotype and these strains are generally regarded as non-pathogenic because the prerequisite virulence genes are missing [Reference Zhang6, Reference Bhagat and Virdi23]. Further, in this study, the most important virulence genes (ail, yadA, virF) are missing in biotype 1A strains (Table 3). All the 14 strains identified as Y. enterocolitica 1A by MALDI–TOF carried the ystB gene. Some evidence indicates that YstB plays a role in the pathogenesis caused by Y. enterocolitica 1A [Reference Bhagat and Virdi23]. Five of the ystB-positive strains also carried hreP. Two ystB-positive strains were also positive for myfA. Both hreP and myfA have sporadically been identified in ystB-positive Y. enterocolitica 1A strains. However, the impact of hreP and myfA in virulence of biotype 1A strains remains unclear [Reference Bhagat and Virdi23]. Some of the 1A strains were identified as serotype O:5 or O:8, which are both associated with human disease; however, the role of these O antigens in virulence of this biotype also remains unclear [Reference Bhagat and Virdi23].
One Y. enterocolitica strain (strain no. 17) that harboured all the important virulence genes was isolated from faeces of a clinically healthy wild alpine ibex (Capra ibex) (Table 3). This strain carries the virulence genes yadA and virF located on the pYV, and ail, ystA, hreP and myfA located in the chromosome. It was identified as serotype O:3 strain with commercial antiserum and PCR targeting the rfbC. Furthermore, it agglutinated very weakly with O:1 and O:2 antisera. This pathogenic Y. enterocolitica belongs either to biotype 3 or biotype 5. Similar to goats, ibex belong to the genus Capra and Y. enterocoliticabelonging to biotype 5 and serotype O:2,3 has already been isolated from goats in New Zealand [Reference Lanada24]. This bioserotype has frequently been associated with Y. enterocolitica infections in goat flocks. Young animals, in particular, have been shown to be susceptible to this infection. Y. enterocolitica 5/O:2,3 has also been isolated from young emaciated goat and sheep with diarrhoea in Australia [Reference Slee and Button25]. In Europe, bioserotype 5/O:2,3 is reported to be host restricted to hares and thus is known as ‘hare type’ [Reference Wuthe and Aleksić26]. Interestingly, Y. enterocolitica belonging to biotype 3 and serotype O:1,2,3 has been isolated from chinchillas with lesions associated with pseudotuberculosis in Europe and North America [Reference Wuthe and Aleksić27]. This type has been assigned as ‘chinchilla type’.
All strains were susceptible to ceftazidim, ciprofloxacin, gentamicin, nalidixic acid, tetracycline and trimethoprim/sulfamethoxazole. They were resistant to ampicillin, amoxicillin/clavulanic acid and cefalothin due to the β-lactamase. Intermediate sensitivity occurred sporadically to cefoxitin, cefpodoxim, cefuroxime, kanamycin and streptomycin (Table 4). No multidrug-resistant strain was detected. The resistance patterns of biotype 1A strains of wild ruminants differed slightly from the patterns of human strains belonging to biotype 1A in Switzerland. The human strains were more frequently resistant to cefoxitin and cefpodoxim and some of them were resistant to kanamycin and nalidixic acid [Reference Fredriksson-Ahomaa16].
Table 4. Antimicrobial resistance patterns in Yersinia strains isolated from wild game
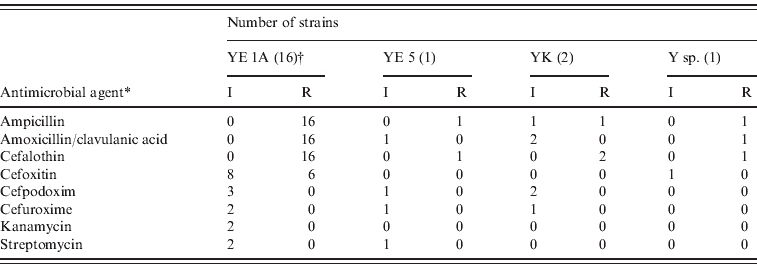
YE, Y. Enterocolitica; YK, Y. kristensenii; Y sp., Yersinia species; I, intermediate; R, resistant.
* only antibiotics where intermediate and resistant strains were found are listed.
† Number of strains studied.
To summarize, clinically healthy wild ruminants are shedding Y. enterocolitica biotype 1A in their faeces. An untypical Y. enterocolitica O:3 strain carrying the most important virulence genes was isolated from a clinically healthy alpine ibex. More studies are needed to clarify the importance of alpine ibex as a reservoir of pathogenic Y. enterocolitica and the significance of this untypical strain in human and animal infections.
ACKNOWLEDGEMENTS
The authors thank D. Ziegler and V. Pflüger, Mabritec AG, Riehen, Switzerland for their assistance with the MALDI TOF experiments and the hunters for their help with collecting the samples.
DECLARATION OF INTEREST
None.




