




Introduction
Coffee (Coffea spp.) is one of the most popular and consumed beverages in the world and an important export agricultural product in many Asian, African and Latin American countries (Mishra, Reference Mishra, Al-Khayri, Jain and Johnson2019). The two species of greatest economic importance are arabica coffee (C. arabica) and conilon or robusta coffee (C. canephora). The conilon or robusta coffee plants are produced in tropical regions, usually in places with altitudes below 1000 m, and responsible for 36% of world production (Campuzano-Duque et al., Reference Campuzano-Duque, Herrera, Ged and Blair2021). Brazil is the second largest producer of conilon coffee in the world, with the state of Espírito Santo (ES) being the main producer in the country, responsible for about 64% of production, with an expanding market (CONAB, 2022).
The largest part of Brazilian coffee is produced in monoculture crops without an inter-row vegetation cover of the coffee plants. The use of monoculture has led to the depletion of agricultural soils, changing the dynamics of soil organic matter and water availability, reducing agricultural productivity and intensifying soil erosion (Cardoso et al., Reference Cardoso, Silva, de Carvalho, de Freitas and Avanzi2012; Melloni et al., Reference Melloni, Belleze, Pinto, Dias, Silve, Melloni, Alvarenga and de Alcântara2013; Franco Junior et al., Reference Franco Junior, Terra, Teruel, Mantovani and Florentino2018). Most Brazilian coffee growers use herbicides to control weeds in the inter-row of coffee plants (Melloni et al., Reference Melloni, Belleze, Pinto, Dias, Silve, Melloni, Alvarenga and de Alcântara2013). Herbicides applied on exposed soil are easily carried away and can contaminate surface waters (Islam et al., Reference Islam, Wang, Farooq, Khan, Xu, Zhu, Zhao, Muños, Li and Zhou2018). The uptake of herbicides by farmers is growing due to falling herbicide prices coupled with steadily rising wage rates (Haggbladea et al., Reference Haggbladea, Mintenb, Prayc, Reardona and Zilberman2017). However, Carvalho et al. (Reference Carvalho, Souza, França, Ferreira, Franco, Kasuya and Ferreira2014) reported that herbicides caused phytotoxicity in coffee seedlings, delayed vegetative growth, and impaired the development of arbuscular mycorrhizal fungi in the soil, beneficial for nutrient absorption. Recent research reports have also indicated the presence of herbicide residues in leaves and processed coffee grains (Schrübbers et al., Reference Schrübbers, Masís-Mora, Rojas, Valverde, Christensen and Cedergreen2016; John and Liu, Reference John and Liu2018).
Cover crops can minimize the application of herbicides and soil degradation under crops (Cardoso et al., Reference Cardoso, Silva, de Carvalho, de Freitas and Avanzi2012; Melloni et al., Reference Melloni, Belleze, Pinto, Dias, Silve, Melloni, Alvarenga and de Alcântara2013). However, the cover crop must be harvested periodically, avoiding competition for water and nutrients that can alter physiological processes and reduce the productivity of the main crop (Ragassi et al., Reference Ragassi, Pedrosa and Favarin2013; Franco Junior et al., Reference Franco Junior, Terra, Teruel, Mantovani and Florentino2018; Ronchi and DaMatta, Reference Ronchi, DaMatta, Ferrão, da Fonseca, Ferrão and De Muner2019; Medic and Baez Veja, Reference Medic and Baez Vega2021). The choice of the species used as cover crops in the inter-row of coffee plants influences productivity, although it still lacks conclusive scientific information.
Warm-season perennial grasses have the potential to be used as cover crops due to the superior biomass accumulation and adaption to different edaphic and climatic conditions (Ragassi et al., Reference Ragassi, Pedrosa and Favarin2013; Grzyb et al., Reference Grzyb, Wolna-Maruwka and Niewiadomska2020; Medic and Boaz Vega, Reference Medic and Baez Vega2021). In addition, warm-season perennial grasses can be used as forage in crop-livestock systems (Jose and Dollinger, Reference Jose and Dollinger2019). Dávila-Solarte et al. (Reference Dávila-Solarte, Sanginés-García, Amezcua and Solano2019) observed that growing coffee plantations in intercropping with lambs is economically viable and offers agronomic and environmental benefits. However, types of management influence the development and nutritional value of forage cover crops (Terra et al., Reference Terra, Gimenes, Giacomini, Gerdes, Manço, de Mattos and Batista2020; Costa et al., Reference Costa, Jank, Magalhães, Bendahan, Rodrigues and Santos2021), as well as the edaphoclimatic conditions (Garay et al., Reference Garay, Joaquin Cancino, Zárate Fortuna, Ibarra Hinojosa, Martínez González, González Dávila and Cienfuegos Rivas2017; Costa et al., Reference Costa, Jank, Magalhães, Bendahan, Rodrigues and Santos2021), lacking studies on their viability in integrated systems in tropical environments.
This research evaluated the herbage accumulation and nutritive value of forages used as cover crops and their effect on productivity, grain size and physiology of conilon coffee plants.
Materials and methods
Characterization of the area
The research was conducted in a commercial plantation of conilon coffee in Rio Bananal – ES, Brazil (latitude 19°15′58″ S, longitude 40°19′60″ W, altitude of 75 m) from April 2020 to June 2022. The climate in the region is Aw, according to the Köppen classification (Alvares et al., Reference Alvares, Stape, Sentelhas, Gonçalves and Sparovek2013), with annual precipitation of 1217 mm and an average temperature of 24.2°C with a rainy period between October and March (Fig. 1). The soil has a clayey texture, with 47% of sand, 7% of silt, and 46% of clay.

Figure 1. Average monthly precipitation of the period (1970 to 2020) and of the agricultural year 2020/21 and 2021/22 in Rio Bananal, Brazil, using data from National Water Agency of Brazil (2022).
The coffee seedlings were planted in March 2018 after subsoiling the planting rows at a depth of 0.60 m with a bulldozer. The spacing was 2.5 m × 1.2 m (3333 plants/ha) with localized drip irrigation. A soil sample was collected in January 2020 between the rows of coffee plants for soil fertility diagnosis (Table 1).
Table 1. Chemical attributes of the soil collected in the 0–0.2 m layer on the inter-row of the conilon coffee plants before planting the cover crops

Analysis methodology: Teixeira et al. (Reference Teixeira, Donagemma, Fontana and Teixeira2017). P, phosphorus; Na, sodium; K, potassium; Ca, calcium; Mg, magnesium; Al, aluminium; CEC, cation exchange capacity; OM, organic matter; AS, aluminium saturation; BS, base saturation.
The inter-row management were: 1- Congo grass, 2- Mombaça guineagrass, 3- Marandu palisadegrass, 4- weeds [Andropogon bicornis L., Lantana camara L., Cnidoscolus urens (L.) Arthur, Solanum Americanum Miller, Baccharis dracunculifolia DC., Ipomoea sp., Vernonia tweediana (Baker) H.Rob., Cenchrus echinatus L., Euphorbia heterophylla L., Cyperus rotundus L.], 5- weeding and application of 200 l/ha of glyphosate herbicide, at a concentration of 10 ml/l (1%). Congo grass, Mombaça guineagrass and Marandu palisadegrass were planted in April 2020 without organic and mineral fertilizers to correct soil fertility, which is naturally acidic (Table 1). Before planting, weeds were eliminated with a hoe, and the herbicide glyphosate was applied in the coffee plants inter-row. Plots (9.0 m2 on each side) were seeded with a manual seeder (Krupp Metal 13AZ, Araricá, Brazil) using the row spacing of 0.5 m and plant spacing within the row of 0.25 m for Congo grass and Marandu palisadegrass, and the row spacing of 0.55 m and plant spacing within the row of 0.35 m for Mombaça guineagrass. Weeds emerged naturally in the area.
The experimental plots had dimensions of 4.0 × 6.0 m (width × length), with five coffee plants and the inter-row on both sides. Each line of coffee plants had a single clone (genotype: P1, LB1, A1, P1), and the inter-row management was distributed side by side with every five coffee plants (6.0 m). Each coffee plant line represented a block, in a total of four blocks, and the coffee plants present in the rows between the blocks represented a border plant (coffee plants not evaluated).
Herbage accumulation of cover crops
An area of 1 m2 was harvested manually using a 0.2 m stubble height and used to estimate herbage accumulation, following the method adapted from Leite et al. (Reference Leite, Cardoso, Fonseca, Silva, Tedeschi, Delevatti, Ruggieri and Reis2021). Two samples were collected per plot when the forage reached 0.5 m (Costa et al., Reference Costa, Jank, Magalhães, Bendahan, Rodrigues and Santos2021), totalling seven sampling events from January 2021 to June 2022. Herbage accumulation was measured in the field with a portable digital scale in seven periods (harvests) from January 2021 to June 2022. The samples were dried in an oven with forced ventilation at 60°C until constant mass was measured with a digital bench scale (AOAC, 2012).
Forages nutritive value
Dried samples from three harvests, July 2021, December 2021, and June 2022, were ground in a Thomas-Willey mill and passed through a 1 mm sieve, adapted from Dias et al. (Reference Dias, Costa, Severiano, Bilego, Furtini Neto, Almeida, Brand and Vilela2020) and Leite et al. (Reference Leite, Cardoso, Fonseca, Silva, Tedeschi, Delevatti, Ruggieri and Reis2021). A subsample was dried at 135°C for two hours to determine the definitive dry matter concentration used to calculate dry matter concentration (AOAC, 2012).
Crude protein was analysed by the Kjeldhal method, and the ether extract was analysed by adapting Garay et al. (Reference Garay, Joaquin Cancino, Zárate Fortuna, Ibarra Hinojosa, Martínez González, González Dávila and Cienfuegos Rivas2017). The total carbon was calculated by the muffle method according to do Carmo and Silva (Reference do Carmo and Silva2012) and used to calculate carbon:nitrogen (C:N) ratio.
Productivity of conilon coffee plants and grain size
Conilon coffee clones productivity was measured in May 2021 and May 2022, analysing the three central plants of the plots containing five coffee plants each, when 80% of the fruits were ripe, i.e. with a reddish colouration of the exocarp (Ronchi and DaMatta, Reference Ronchi, DaMatta, Ferrão, da Fonseca, Ferrão and De Muner2019; Mulindwa et al., Reference Mulindwa, Kaaya, Muganga, Paga, Musoli, Sseremba, Wagoire and Bitalo2021). The fruits were harvested manually and measured volumetrically with a 20-l graduated bucket.
Samples with 2.0 l of coffee fruits were placed in perforated bags and dried to 12% humidity on a suspended and covered terrace. The dried fruits were peeled, and the grains were weighed to calculate the grain/fruit ratio. The grain/fruit ratio was obtained by the ratio between the mass of processed grains and the initial volume of ripe fruits and used to calculate the mass productivity of grains per area (Franco Junior et al., Reference Franco Junior, Terra, Teruel, Mantovani and Florentino2018; Pereira et al., Reference Pereira, Sera, Sera, Shigueoka, Carducci, Dias da Silva and Telles2022), i.e. in bags (60 kg) per hectare. The size of the processed grains was measured using a set of sieves with holes ranging from 16/64″ (6.35 mm) to 8/64″ (3.175 mm), according to the Official Brazilian Classification (Brasil, 2003) and grouped into three classes, being ⩾15/64″, 13–14/64″ and ⩽12/64″.
Evaluation of the vegetative vigour of coffee plants
The vegetative vigour of coffee plants was measured in July 2021 and June 2022 using the normalized difference vegetation index (NDVI), acquired by the GreenSeeker portable meter (Trimble, Sunnyvale, USA), adapted from Martello et al. (Reference Martello, Molin, Bazame, Tavares and Maldaner2022). The apparatus was positioned at a height of 1.5 m above and parallel to the ground. The reading was performed at 0.5 m of linear distance from the coffee plants. Readings were taken on the three central plants of the plot on both sides of the planting row between 9:00 and 10:00 h (GMT, BRS, 3 h day), adapted from Quartezani et al. (Reference Quartezani, Sales, Berilli, Pletsch, Rodrigues, Campostrini and Mantoanelli2018). After activating the sensor, the reading occurs every 0.1 s, generating between 30 and 40 measurements per reading.
Physiological indices of coffee plants
Fluorescence indices of coffee plant leaves were collected in July 2021 and June 2022 with a Multiplex 330 optical fluorescence sensor (Force-A, Orsay, France), with six sources of light excitation, being ultraviolet (UV, 375 nm), blue (B, 450 nm), green (G, 515 nm) and red (R, 635 nm). The detection of fluorescence emitted by chlorophyll occurs in the red (RF) and far red (FRF) spectrum and was used to estimate the nitrogen balance index (NBI-G and NBI-R), chlorophyll (SFR-G and SFR-R), anthocyanin (ANT-RG and ANT-RB) and flavonoids (FLAV), adapted from Agati et al. (Reference Agati, D'Onofrio, Ducci, Cuzzola, Remorini, Tuccio, Lazzini and Mattii2013) and Abdallah et al. (Reference Abdallah, Philippe and Goffart2018).
Readings were taken on the three central plants of the plot on both sides of the planting row, between 9:00 and 10:00 h (GMT, BRS, 3 h day), adapted from Quartezani et al. (Reference Quartezani, Sales, Berilli, Pletsch, Rodrigues, Campostrini and Mantoanelli2018). The device was positioned at a height of 1.5 m above and parallel to the ground, directed towards the top of the coffee plants, from top to bottom, at an angle of 45° and a linear distance of 0.5 m.
Statistical design
The experiment was installed in a split-plot design (inter-row management [n = 5] and periods [n = 7, 3 or 2]) with four randomized blocks (replications), according to Eqn (1):

where Υijk is the response variable, μ is a mean, αi is the effect of the whole-plot, ηk(i) is the whole-plot error, βj is the effect of the split-plot, (αβ)ij is the interaction between factors, and ɛk (ij) is the split-plot error.
Data were submitted for analysis of variance using the F test (P > 0.05) and comparison of means using the Tukey test (P > 0.05) in the R Core Team (2017) software. The interaction between the factors of inter-row management and period was considered significant for the productivity of conilon coffee plants with P < 0.1 due to the seasonality of the data in both periods, which agreed with field observations and with Megerssa et al. (Reference Megerssa, Welde Michael and Teshome2012) and Tanimonure and Naziri (Reference Tanimonure and Naziri2021).
Results
Herbage accumulation and nutritive value of cover crops
Herbage accumulation showed a significant interaction between inter-row management and periods (P < 0.001). The herbage accumulation of Congo grass, Mombaça guineagrass and Marandu palisadegrass (1.12 to 3.81 t/ha) were higher than the weeds (0.18 to 1.95 t/ha) in the evaluated periods, except for Congo grass on 13 July 2021 and 18 June 2022, not statistically different from the weeds (Table 2). Mombaça guineagrass presented an average herbage accumulation 29.5% higher than the other forages cover crops (1.47 to 3.81 t/ha), differing statistically from at least one of them on 28 January 2021, 13 July 2021, 24 November 2021, 17 December 2021, and 18 June 2022. The herbage accumulation of cover crops (2.32 t/ha) was 89% higher in the rainy season (October to March) compared to the dry season (1.23 t/ha).
Table 2. Herbage accumulation (t/ha) of cover crops in the inter-row of conilon coffee plants in the evaluated periods

Mean ± standard error. HSD, honestly significant difference (Tukey's test, P = 0.05).
Dry matter (P = 0.969), crude protein (P = 0.931), and C:N ratio (P = 0.427) showed no significant interaction between inter-row management and periods that were evaluated separately. Congo grass, Mombaça guineagrass and Marandu palisadegrass did not differ statistically from each other for dry matter (P = 0.437), crude protein (P = 0.615), and C:N ratio (P = 0.419) in the three evaluations performed (Table 3). However, dry matter concentration (P < 0.001), crude protein (P < 0.001), and C:N ratio (P < 0.001) differed statistically between periods. Dry matter concentration and crude protein contents decreased with the increase in the number of cuts, from 13 July 2021 (29.0 and 11.7%) to 18 June 2022 (21.5 and 10.5%), while the C:N ratio increased (24:1 to 61:1).
Table 3. Dry matter concentration (DM), crude protein (CP) and carbon:nitrogen ratio (C:N) of cover crops in the inter-row of conilon coffee plants
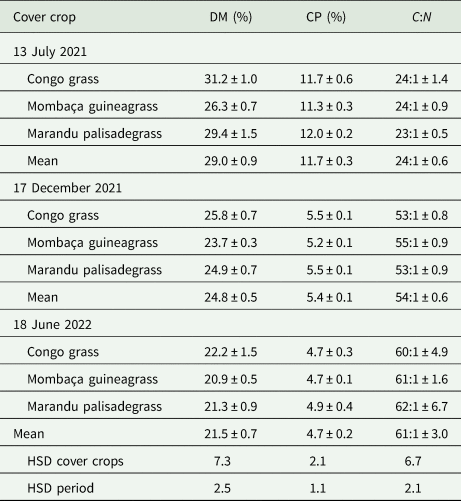
Mean ± standard error. HSD, honestly significant difference (Tukey's test, P = 0.05).
Productivity of conilon coffee plants and grain quality
Productivity (P = 0.092), grain/fruit ratio (P = 0.855), and grain size classes ⩾15/64″ (P = 0.560), 13–14/64″ (P = 0.827) and ⩽12/64″ (P = 0.119) showed no significant interaction between inter-row management and periods. Inter-row management differs statistically for conilon coffee productivity for the two years average (P = 0.002), with weeding and herbicide application showing higher productivity with 125.7 bags/ha (7545 kg/ha). The inter-row management with Congo grass, Mombaça guineagrass and Marandu palisadegrass obtained the lowest values, ranging from 96.0 to 82.0 bags/ha (5761 to 4922 kg/ha). However, the results demonstrated a variation in productivity between 2021 and 2022, indicating a tendency towards statistical significance of the interaction between inter-row management and period (P < 0.1). In 2021, inter-row management did not differ statistically for conilon coffee productivity, ranging from 102.0 to 115.2 bags/ha (6120 to 6912 kg/ha). Nevertheless, in 2022, the inter-row management with weeding and herbicide application showed higher productivity with 149.5 bags/ha (8970 kg/ha), while the inter-row management with Congo grass, Mombaça guineagrass and Marandu palisadegrass obtained the lowest values, ranging from 56.3 to 84 bags/ha (3378 to 5040 kg/ha). The inter-row management with weeding and herbicide application showed a 47% increase in the productivity of conilon coffee plants in 2022 compared to 2021, while the inter-row management with Congo grass, Marandu palisadegrass and weeds productivity did not vary statistically, and the inter-row management with Mombaça guineagrass showed a 48% reduction in productivity.
Inter-row management did not differ statistically (P = 0.358) for grain/fruit ratio in 2021 and 2022, ranging from 0.14 to 0.15 and from 0.17 to 0.20 kg/l, respectively (Table 4). The same also occurred for the grain size classes ⩾15/64″, 13–14/64″, and ⩽12/64″, ranging from 71.4 to 84.9%, 13.9 to 24.8%, and 1.2 to 7.0%, respectively. The grain/fruit ratio increased by 29% in 2022 (0.18 kg/l) compared to 2021 (0.14 kg/l). The grain size classes of conilon coffee ⩾15/64″ (P = 0.270), 13–14/64″ (P = 0.162), and ⩽12/64″ (P = 0.296) did not differ statistically between 2021 and 2022.
Table 4. Productivity, grain/fruit ratio and grain size classes of conilon coffee plants in the inter-row management with cover crops and with weeding and herbicide application

Mean ± standard error. HSD, honestly significant difference (Tukey's test, P = 0.05). Bags = 60 kg of processed grains (12% moisture and peeled).
Vegetative vigour and physiological indices of coffee plants
The vegetation index per normalized difference (P = 0.280), chlorophyll index (SFR-G [P = 0.359] and SFR-R [P = 0.468]), nitrogen balance index (NBI-G [P = 0.870] and NBI-R [P = 0.922]), anthocyanin (ANTH-RB [P = 0.221]) and flavonoids (P = 0.187) showed no significant interaction between inter-row management and periods while the anthocyanin (ANTH-RG [P = 0.014]) showed the opposite.
The vegetation index per normalized difference (P = 0.509 and P < 0.001), chlorophyll index (SFR-G [P = 0.113 and P < 0.001] and SFR-R [P = 0.109 and P = 0.085]), nitrogen balance index (NBI-G [P = 0.675 and P < 0.001] and NBI-R [P = 0.423 and P < 0.001]), anthocyanin (ANTH-RB [P = 0.415 and P < 0.001]) and flavonoids (P = 0.724 and P < 0.001) showed no significant interaction for inter-row management but showed a significant effect for periods (Table 5). The inter-row management did not differ statistically for the vegetation index per normalized difference and physiological indices in 2021 and 2022, except for the anthocyanin index (ANT-RG) in 2022, with the highest values for inter-row management with cover crops (0.03 to 0.06) and the lowest for inter-row management with weeding and herbicide application (0.02).
Table 5. Normalized difference vegetation index (NDVI), chlorophyll (SFR-G and SFR-R), nitrogen balance (NBI-G and NBI-R), anthocyanin (ANT-RG and ANT-RB) and flavonoids (FLAV) of conilon coffee plants in the inter-row management with cover crops and with weeding and herbicide application

Mean ± standard error. HSD, honestly significant difference (Tukey's test, P = 0.05).
In 2022, the inter-row management presented a vegetation index per normalized difference lower than in 2021 (0.79 vs. 0.83). The same occurred for the chlorophyll index (SFR-G [0.91 vs. 3.20]) and flavonoids (FLAV [0.79 vs. 1.20]), with the opposite occurring for the nitrogen balance index (NBI-G [0.18 vs. 0.10] and NBI-R [0.59 vs. 0.19]) and anthocyanin (ANTH-RG [0.04 vs. −0.33] and ANTH-RB [1.79 vs. −0. 64]).
Discussion
Herbage accumulation and nutritive value of cover crops
Congo grass, Mombaça guineagrass and Marandu palisadegrass produced greater herbage accumulation than the weeds. The lower herbage accumulation of weeds was due to the residual effect of two years of herbicides use in the soil, applied before the installation of the experiment, eliminating weeds originating from seeds in the soil, reducing the potential of the soil seeds bank (Beckie et al., Reference Beckie, Ashworth and Flower2019). Mombaça guineagrass was the cover crop with higher herbage accumulation, related to genetic factors and its adaptability to tropical climate regions (Dias et al., Reference Dias, Costa, Severiano, Bilego, Furtini Neto, Almeida, Brand and Vilela2020; Costa et al., Reference Costa, Jank, Magalhães, Bendahan, Rodrigues and Santos2021). A greater Mombaça guineagrass forage accumulation can occur in conditions of greater soil fertility and irrigation, while species of the genus Urochloa are more adapted to soils with low fertility and periods of water deficit (Costa et al., Reference Costa, Jank, Magalhães, Bendahan, Rodrigues and Santos2021).
The herbage accumulation of cover crops was higher in the rainy season compared to the dry season. It occurs due to climatic oscillations that occur in a year (Fig. 1), as the reduction of precipitation, temperature and photoperiod in the dry season, fundamental for photosynthesis, which decreases the growth of forage plants, resulting in the seasonality of production (Xia et al., Reference Xia, Yuan, Wang and Zhang2017; Terra et al., Reference Terra, Gimenes, Giacomini, Gerdes, Manço, de Mattos and Batista2020). In periods with lower water availability, plants assign more carbon to the root system than to the shoots as an adaptive response, reducing the production of new tillers (Maia et al., Reference Maia, Pinho Costa, Costa Severiano, Epifanio, Neto, Ribeiro, Fernandes, Guimarães Silva and Gonçalves2014; Xia et al., Reference Xia, Yuan, Wang and Zhang2017). Furthermore, weeds have a lower range of herbage accumulation between periods, indicating phenotypic plasticity in the tropical environment (Terra et al., Reference Terra, Gimenes, Giacomini, Gerdes, Manço, de Mattos and Batista2020).
The low natural soil fertility (Table 1) resulted in no statistical difference in dry matter concentration, crude protein and C:N ratio between forage cover crops, preventing mainly the Mombaça guineagrass from expressing its nutritional potential (Costa et al., Reference Costa, Jank, Magalhães, Bendahan, Rodrigues and Santos2021). Another factor that minimized differences between forage cover crops was the rest period between cuts that, when increased, reduces the nutritional value of forages (Terra et al., Reference Terra, Gimenes, Giacomini, Gerdes, Manço, de Mattos and Batista2020; Costa et al., Reference Costa, Jank, Magalhães, Bendahan, Rodrigues and Santos2021). For the coffee producer, increasing the number of forage plants represents an increase in the cost of production. However, for crop-livestock integration systems, this adjustment is necessary and may improve economic viability (Dávila-Solarte et al., Reference Dávila-Solarte, Sanginés-García, Amezcua and Solano2019). This nutritional limitation of cover crops stimulates root growth towards the coffee rows, generating competition for water and nutrients with the coffee plants (Partelli et al., Reference Partelli, Duarte, Freitas and Espindola2010; Ronchi and DaMatta, Reference Ronchi, DaMatta, Ferrão, da Fonseca, Ferrão and De Muner2019).
Climatic conditions throughout the year and plant age affect the nutritive value of forage cover crops (Garay et al., Reference Garay, Joaquin Cancino, Zárate Fortuna, Ibarra Hinojosa, Martínez González, González Dávila and Cienfuegos Rivas2017; Maia et al., Reference Maia, Pinho Costa, Costa Severiano, Epifanio, Neto, Ribeiro, Fernandes, Guimarães Silva and Gonçalves2014). Dry matter concentration decreased over time due to higher water content in the leaf tissue, a result of rainfall above the historical average, with 1345 and 1375 mm/year in 2020/21 and 2021/22, respectively (Fig. 1), in agreement with Serrano et al. (Reference Serrano, Shahidian and Marques da Silva2016). Crude protein values found in Congo grass, Mombaça guineagrass and Marandu palisadegrass (11.3 to 12.0%) on 17 March 2021 corroborate the value obtained by Oliveira et al. (Reference Oliveira, Costa, Severiano, da Silva, Dias, Oliveira and Costa2020) for forages of the genus Urochloa, ranging from 9.2 to 12.2%. However, the values found on 17 December 2021 and 18 June 2022 were lower, ranging from 4.7 to 5.4%. Garay et al. (Reference Garay, Joaquin Cancino, Zárate Fortuna, Ibarra Hinojosa, Martínez González, González Dávila and Cienfuegos Rivas2017) also found a reduction in crude protein values with increasing regrowth age for different forages and fluctuations throughout the year, from 14.1% in the rainy season to 7.6% in the dry season. The lowest crude protein values are also related to the low availability of nutrients in the soil (Table 1), in agreement with Leite et al. (Reference Leite, Cardoso, Fonseca, Silva, Tedeschi, Delevatti, Ruggieri and Reis2021). Increasing forage age decreased the crude protein concentration due to the reduction in the leaf:stem ratio, in agreement with Maia et al. (Reference Maia, Pinho Costa, Costa Severiano, Epifanio, Neto, Ribeiro, Fernandes, Guimarães Silva and Gonçalves2014), reaching concentrations lower than the minimum considered (7%) when intake by ruminants could be suppressed (Lazzarini et al., Reference Lazzarini, Detmann, Sampaio, Paulino, Valadares Filho, Souza and Oliveira2009).
The C:N ratio increased from 13 July 2021 to 18 June 2022. Cavalli et al. (Reference Cavalli, Lange, Cavalli and Behling2018) found a C:N ratio for Brachiaria (syn. Urochloa) residues ranging from 32:1 to 56:1. Dias et al. (Reference Dias, Costa, Severiano, Bilego, Furtini Neto, Almeida, Brand and Vilela2020) found C:N ratio values of Mombaça guineagrass ranging from 29:1 to 37:1. The C:N ratio of residues added to the soil influences decomposition and the relationship between mineralization and nitrogen immobilization by microbial biomass (Grzyb et al., Reference Grzyb, Wolna-Maruwka and Niewiadomska2020). Straw with a low C:N ratio decompose quickly, reducing the mulch persistence time on the soil surface (Cavalli et al., Reference Cavalli, Lange, Cavalli and Behling2018). However, if the C:N ratio of plant residues is greater than 25:1, the amount of mineralized nitrogen is not enough to meet the demand of microorganisms, which start to immobilize the mineral nitrogen available in the soil, compromising nitrogen nutrition of crops (Grzyb et al., Reference Grzyb, Wolna-Maruwka and Niewiadomska2020; Watthier et al., Reference Watthier, Antonio, Gomes, Rocha and Santos2020). In tropical regions, decomposition is favoured by environmental conditions, and therefore covers with a C:N ratio greater than 25:1 is favourable, that is, more stable and with a longer time of permanence in the soil (Cavalli et al., Reference Cavalli, Lange, Cavalli and Behling2018).
Productivity of conilon coffee plants and grain quality
The initial growth of cover crops was in 2021 with the superficial and localized root system, not causing competition for water and nutrients with the coffee plants, so coffee productivity did not differ statistically between the inter-row management with cover crops in relation to weeding and herbicide application. Partelli et al. (Reference Partelli, Duarte, Freitas and Espindola2010) also observed that the cover crops did not disturb the vegetative growth and nutrient concentrations in the leaves of the conilon coffee plants. In 2022, coffee productivity in inter-row management with weeding and herbicide application increased by 47% compared to 2021, while the inter-row management with cover crops stagnated or reduced by up to 48%.
Increased productivity of conilon coffee was expected in 2022 compared to 2021 due to greater rooting and vegetative development of plants, with elongation of orthotropic and plagiotropic branches, with a substantial number of internodes and flower buds, resulting in a higher production of coffee fruits (Ronchi and DaMatta, Reference Ronchi, DaMatta, Ferrão, da Fonseca, Ferrão and De Muner2019; Colodetti et al., Reference Colodetti, Rodrigues, Brinate, Martins, Cavatte and Tomaz2020). This stagnation or reduction in coffee productivity in inter-row management with cover crops in 2022 was due to root development and ability to compete for water and nutrients with coffee plants, in agreement with Franco Junior et al. (Reference Franco Junior, Terra, Teruel, Mantovani and Florentino2018). Less competition occurred in inter-row management with weeds, while inter-row management with forages resulted in lower productivity due to herbage accumulation in the period. It is important to emphasize that the productivity of the coffee plants found for the inter-row management in the two years was higher than the highest national average productivity of conilon coffee in Brazil reached in 2022 with 46.2 bags/ha (2772 kg/ha) (CONAB, 2022).
Coffee spp. are perennial plants sensitive to competition for water and nutrients exerted by weeds (Ragassi et al., Reference Ragassi, Pedrosa and Favarin2013). This competition have negative effects on coffee plant growth and crop productivity (Ragassi et al., Reference Ragassi, Pedrosa and Favarin2013; Franco Junior et al., Reference Franco Junior, Terra, Teruel, Mantovani and Florentino2018). The critical period of competition takes place from flowering to fruiting, which comprises in southeastern Brazil from September to March, corresponding to the rainy season (Ronchi and DaMatta, Reference Ronchi, DaMatta, Ferrão, da Fonseca, Ferrão and De Muner2019). This period was the phase of greatest growth of cover crops (Table 2), which favoured competition with coffee plants (Franco Junior et al., Reference Franco Junior, Terra, Teruel, Mantovani and Florentino2018; Ronchi and DaMatta, Reference Ronchi, DaMatta, Ferrão, da Fonseca, Ferrão and De Muner2019). The effect of the competition is conditioned by the vigour of cover crops, which can be minimized with a higher frequency of harvesting, especially in the rainy season. Although cover crops reduce the presence of weeds, in case of abundant growth, this positive effect will be eliminated by the competition generated by the cover crops themselves (Melloni et al., Reference Melloni, Belleze, Pinto, Dias, Silve, Melloni, Alvarenga and de Alcântara2013; Beckie et al., Reference Beckie, Ashworth and Flower2019; Grzyb et al., Reference Grzyb, Wolna-Maruwka and Niewiadomska2020). In this case, cover crops can play the role of weeds. Therefore, it is necessary, especially in the period from flowering to fruiting, the proper management of cover crops, such as reducing the strip (width) with cover crops, increasing planting spacing and frequency of mowing or using a greater intensity of grazing in agropastoral systems (Ragassi et al., Reference Ragassi, Pedrosa and Favarin2013; Dávila-Solarte et al., Reference Dávila-Solarte, Sanginés-García, Amezcua and Solano2019). An agropastoral system allows the addition of value, diversification of economic activity, and better use of natural resources and inputs (Jose and Dollinger, Reference Jose and Dollinger2019).
The inter-rows management did not influence the grain/fruit ratio and grain size in 2021 and 2022. Mulindwa et al. (Reference Mulindwa, Kaaya, Muganga, Paga, Musoli, Sseremba, Wagoire and Bitalo2021) also did not observe the effect of environmental variations on the grain size of Coffea canephora. Although the inter-row management did not differ statistically for grain size between 2021 and 2022, there was an increase in the average grain/fruit ratio in 2022 compared to 2021 (0.18 vs. 0.14 kg/l). These results demonstrated the absence of the effect of the cover crops competition, not influencing the grain/fruit ratio and grain size of the conilon coffee plants, which would be more related to climate and genotype characteristics (Fialho et al., Reference Fialho, da Fonseca, Ferrão, Ferrão, Olivoto, Nardino, dos Reis and Sakiyamo2022).
Vegetative vigour and physiological indices of coffee plants
The absence of statistical difference for the vegetation index per normalized difference and physiological indices in 2021 can be explained by the high vigour and productivity of coffee plants of the inter-row management in relation to the national average (CONAB, 2022), indicating a lack of competition with cover crops in 2021. In 2022, inter-row management with weeding and herbicide application had the lowest anthocyanin content (ANT-RB), differing statistically from inter-row management with Mombaça guineagrass, which indicated a result of cover crop competition for water and nutrients. More stressed plants, whether due to toxicity, competition for water, nutrients or light, tend to produce higher levels of secondary compounds, such as anthocyanin (Quartezani et al., Reference Quartezani, Sales, Berilli, Pletsch, Rodrigues, Campostrini and Mantoanelli2018). The effect of competition was evident in the productivity data of coffee plants in 2022 (Table 4), which could be economically compensated in an agropastoral system (Dávila-Solarte et al., Reference Dávila-Solarte, Sanginés-García, Amezcua and Solano2019).
Anthocyanins have functions related to antioxidants, acting in protection against the effects of sunlight, defence mechanisms and biological functions (Agati et al., Reference Agati, D'Onofrio, Ducci, Cuzzola, Remorini, Tuccio, Lazzini and Mattii2013; Quartezani et al., Reference Quartezani, Sales, Berilli, Pletsch, Rodrigues, Campostrini and Mantoanelli2018). This protection against photoinhibition is important for regions with a tropical climate, as in the case of this work. Changes in vegetation during growth result in a differentiation of fluorescence, which allows the use of spectral sensors for monitoring vegetation and the detection of physiological changes in plants (Abdallah et al., Reference Abdallah, Philippe and Goffart2018). Quartezani et al. (Reference Quartezani, Sales, Berilli, Pletsch, Rodrigues, Campostrini and Mantoanelli2018) also observed differences in the physiological indices measured with a Multiplex sensor in conilon coffee seedlings associated with different types of organic matter used in the substrate to produce seedlings.
The reduction in the average for vegetation and chlorophyll indices in 2022 compared to 2021 is related to the lower capacity to produce photoassimilates by coffee plants intercropped with cover crops (Colodetti et al., Reference Colodetti, Rodrigues, Brinate, Martins, Cavatte and Tomaz2020), also resulting in lower average productivity (Table 4). Martello et al. (Reference Martello, Molin, Bazame, Tavares and Maldaner2022) observed relationships between NDVI and coffee productivity. Inter-row management with weeding and herbicide application reduced of chlorophyll in 2022 due to the depletion of coffee plants when they reach high levels of productivity (Pereira et al., Reference Pereira, Sera, Sera, Shigueoka, Carducci, Dias da Silva and Telles2022), that is, reducing the pigments of the leaves and redirecting the metabolites to the formation of grains (Colodetti et al., Reference Colodetti, Rodrigues, Brinate, Martins, Cavatte and Tomaz2020). Often, plant depletion is observed only in the following year's productivity, creating biennial productivity cycles (Ronchi and DaMatta, Reference Ronchi, DaMatta, Ferrão, da Fonseca, Ferrão and De Muner2019; Pereira et al., Reference Pereira, Sera, Sera, Shigueoka, Carducci, Dias da Silva and Telles2022). The increase in nitrogen balance indices in 2022 was related to the effect of coffee competition for water and nutrients in inter-row management with cover crops, with the need for coffee plants to translocate nitrogen to leaf tissues (Colodetti et al., Reference Colodetti, Rodrigues, Brinate, Martins, Cavatte and Tomaz2020) and the greater need for microorganisms to decompose residues with a higher C:N ratio (Dias et al., Reference Dias, Costa, Severiano, Bilego, Furtini Neto, Almeida, Brand and Vilela2020; Watthier et al., Reference Watthier, Antonio, Gomes, Rocha and Santos2020), making coffee plants more efficient.
Conclusions
The productivity of forage plants was higher weeds in the evaluated periods, being a viable option for conservation management in coffee plantations. Among the forages, Mombaça guineagrass presented the highest average productivity, being more advantageous for soil cover and use in the crop-livestock system. Cover crops did not cause a reduction in productivity, grain/fruit ratio, grain size, vegetative vigour, and physiological indices of the coffee plants in 2021. However, in 2022, there was a reduction of 16 to 49% in productivity, with changes in vegetative vigour and physiological indices, indicating the need for adjustments in management for the development of a competitive and sustainable coffee culture.
Authors’ contributions
G. S. de Souza and I. R. Pretti conceived and designed the study. L.C.L. Coelho, G.R. Sarnaglia, and J. Elias conducted data gathering. L.C.L. Coelho and E. C. de Oliveira performed statistical analyses. G. S. de Souza, L. L. Bitencourt, L. L. Pereira wrote the article. R. F. de Almeida and S. da S. Berilli revised it critically for intellectual content.
Funding statement
This work was supported by the PIBIC/IFES (G.R. Sarnaglia, grant numbers 10108/21), (J. Elias, grant number 10107/21), (L.C.L. Coelho, grant number 9084/20).
Competing interests
None.
Ethical standards
Not applicable.






