



Introduction
The Bismarck Archipelago, off the northeast coast of New Guinea, represents an important region for research on Early human activity. The Sahul landmass, together comprising New Guinea and Australia during a time of lower sea level, was settled 50–60 thousand years ago (Clarkson et al. Reference Clarkson, Smith, Marwick, Fullagar, Wallis, Faulkner, Manne, Hayes, Roberts, Jacobs, Carah, Lowe, Matthews and Florin2015). Along with hunting and animal commensals, this migration, into an area of high faunal diversity and endemism, resulted in mega-faunal extinctions and eventually agriculture. The latter was developed independently (Denham et al. Reference Denham2004) and included both plant and animal husbandry. In the Late Holocene, another migrant wave occurred, involving the Lapita culture, who were the first to colonise Remote Oceania (Kirch Reference Kirch2000).
The commonly accepted account for the initial presence of Lapita occupation in the Western Pacific is that it represented new people who brought with them new ideas, language, and a translocated economy (Kirch Reference Kirch2000). This period is called Early Lapita (Summerhayes Reference Summerhayes2000) and covers the first settlement phase dated from 3300–3000 cal. BP (Summerhayes Reference Summerhayes, Gadu and Lin2010). Examples of economic introductions include pig, dog, and chicken, all of which would have originated from Southeast Asia. The economy of these Early Lapita settlers was seen as involving broad-spectrum foraging, domestic animals, horticulture, and arboriculture. A detailed, comprehensive study of Lapita culture is given in the recent work edited by Bedford and Spriggs (Reference Bedford and Spriggs2019).
Archaeological studies of settlement sites in the Bismarck Archipelago have focused largely on tool, faunal, and botanical records, and more most recently DNA analysis, yielding a varied body of evidence of the nature and timing of cultural activity and development (Bedford & Spriggs Reference Bedford and Spriggs2019). At the time of this writing, there appear to be no confirmed reports of palaeoparasitological analysis in the archipelago, in contrast to the varied literature from the Northern Hemisphere. Beyond Melanesia in wider Oceania, such research is restricted to New Zealand alone (Horrocks et al. Reference Horrocks, Brown, Brown and Presswell2023a; Horrocks & Presswell Reference Horrocks and Presswell2023b; Irwin et al. 2024). This kind of analysis can provide evidence of past diet, agricultural practices, animal domestication, migration routes, climate fluctuations, hygiene practices, cultural interactions, ethnomedicine, and the overall health status of past human societies (https://en.wikipedia.org/wiki/Archaeoparasitology). Microscopic eggs of gastro-intestinal parasites (helminths) can be readily extracted from soils, sediments, and coprolites using density separation or flotation techniques (Anastasiou & Mitchell Reference Anastasiou and Mitchell2013; David & Lindquist Reference David and Lindquist1982; Horrocks Reference Horrocks2020).
During 1997–2001, archaeological excavations were carried out at Kamgot, an Early Lapita settlement site, in the Anir Island Group of the Bismarck Archipelago, to shed light on Early Lapita settlement (Summerhayes Reference Summerhayes2000; Summerhayes et al. Reference Summerhayes, Leavesley and Fairbairn2009). That work included analysis of plant microfossil samples from a habitation layer soil in each of two test pits (Summerhayes et al. Reference Summerhayes, Szabo, Fairbairn, Horrocks, McPherson, Crowther, Bedford and Spriggs2019a, Reference Summerhayes, Szabó, Leavesley, Gaffney, Bedford and Spriggsb). In the present study, we examined the same samples for helminth eggs, with the aim of (1) building on the previous research, providing a new type of evidence for hunted and commensal animals associated with human activity at the site; and (2) introducing this under-utilised, potentially highly informative method to the archaeological study of tropical Oceania.
Study area and site
The site of Kamgot is located on the northwest coast of Babase Island, Anir Island Group. Babase is mostly volcanic, but its western end comprises raised limestone. Today the site is 100 m inland from the high-tide mark, with the old, raised reef forming a raised beachfront. During the initial Lapita occupation, the settlement was spread over both a sand bar at or just above the high-water mark and a stilt village occupation over a shallow lagoon parallel to an outer reef some 80 m away, with swamp or fresh water available on the landward side. The Summerhayes et al. (Reference Summerhayes, Szabo, Fairbairn, Horrocks, McPherson, Crowther, Bedford and Spriggs2019a) study focused on faunal and botanical remains.
The site of Kamgot provided new insights in the chronology of Early Lapita and the nature of society and interaction across the Bismarck Archipelago. It also provided important data on Early Lapita subsistence and demonstrated the presence of domesticated animals and horticulture. Full details of the site and results of the different types of analyses carried out are given in the original Summerhayes et al. (Reference Summerhayes, Szabo, Fairbairn, Horrocks, McPherson, Crowther, Bedford and Spriggs2019a) report.
Methods
Parasitological analysis of samples from each of the two test pits (TP 1 and TP 23) at Kamgot (Summerhayes et al. Reference Summerhayes, Szabo, Fairbairn, Horrocks, McPherson, Crowther, Bedford and Spriggs2019a) was carried out. Samples were prepared by density separation (1.7–1.8 specific gravity) with sodium polytungstate (David & Linquist Reference David and Lindquist1982; Horrocks Reference Horrocks2020), and presence/absence was noted. Photomicrographs were taken with a Canon EOS 600D camera (Auckland Camera Centre, Auckland, New Zealand) mounted on a Nikon 400E microscope (Olympus New Zealand Ltd, Auckland, New Zealand), with a blue light filter. Measurements were made using a calibrated ocular micrometre.
Results
Six types of parasitic helminth eggs were identified: Trichuris suis, Dipylidium caninum, and Toxocara canis, as well as two unknown trematodes and a possible anoplocephalid cestode.
Trichuris suis
Nematode of family Trichuridae: two specimens found in TP 23. Dark, brown, oval eggs with bipolar plugs (plugs missing in both specimens), a thick shell with a smooth outer surface. These characteristics identify the eggs as belonging to genus Trichuris. The better specimen measures 58 μm x 31 μm (Figure 1a). Given the context, these are most likely to belong to either the human whipworm (T. trichiura), the pig whipworm (T. suis), or the dog whipworm (T. vulpis). However, the egg of T. trichiura (54–60 x 24–29 μm) is consistently smaller than that of T. suis (57–68 x 25–30 μm) (Sondak Reference Sondak1948) or of T. vulpis (72–90 x 32–40μm) (Yevstafieva et al. 2019). As rat remains (Rattus exulans) were collected from the site, we also compared the specimens to T. muris, the eggs of which are also larger (62–68 x 28–32 μm) (Felui et al. 2000). The smaller size of the eggs places the specimens found here in T. suis.
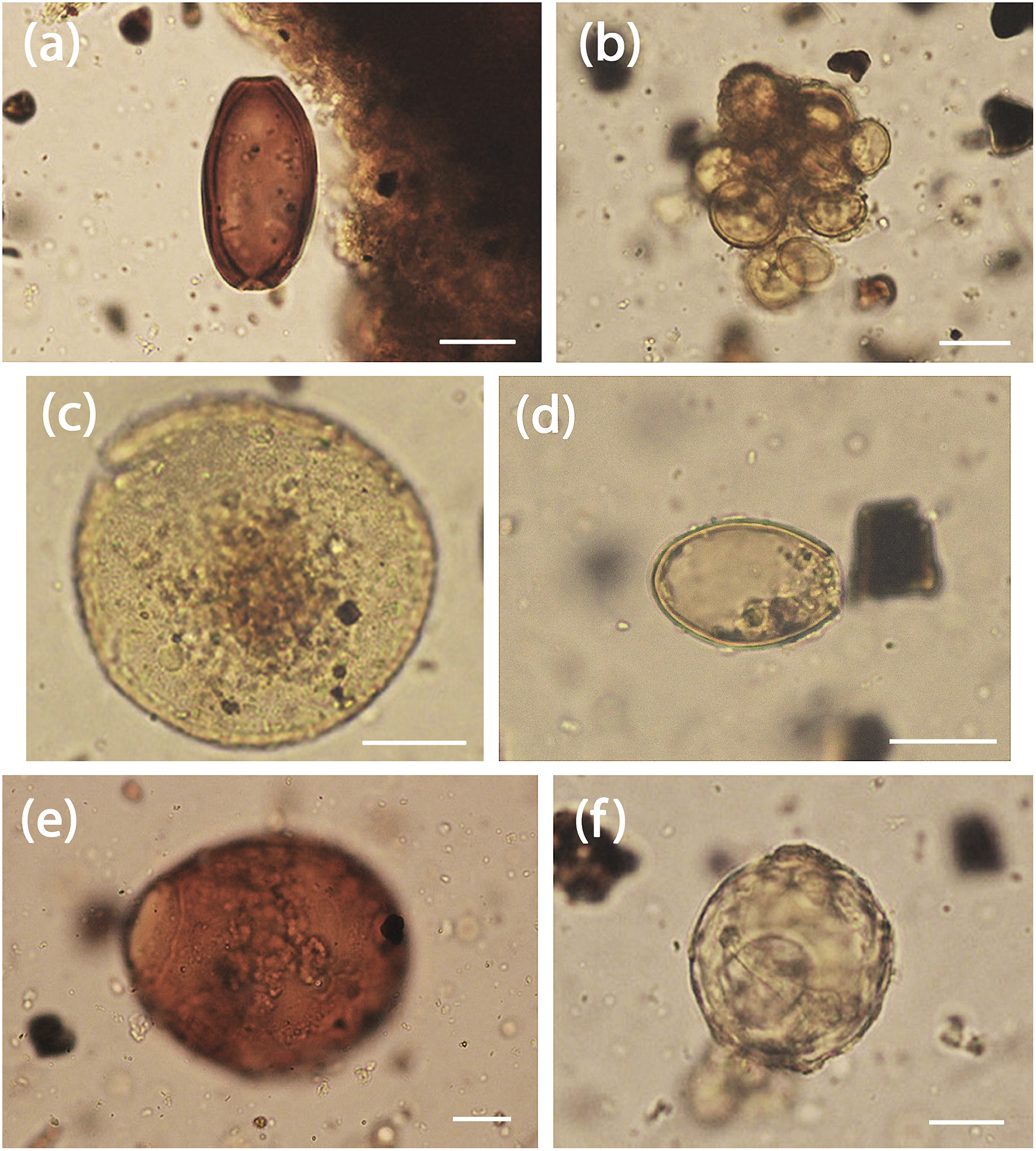
Figure 1. Helminth eggs from Kamgot Early Lapita archaeological site, Bismarck Archipelago (all from TP 23, mounted in glycerol jelly, scale bars 20 μm). (a) Trichuris suis egg, bipolar plugs missing; (b) Dipylidium caninum egg packet containing >10 eggs; (c) Toxocara canis egg showing pitted surface; (d) unknown trematode egg #1, showing single opercular opening; (e) unknown trematode egg #2, showing thick brown shell and single opercular opening; (f) unknown cestode egg, possibly attributable to Anoplocephalidae.
Dipylidium caninum
Cestode of family Dipylidiidae: one specimen found in TP 23. Packet or agglomeration (60–70 μm longest axis) of >10 round eggs (23–27 x 19–22 μm), colourless to light brown (Figure 1b). The outer envelope missing or not visible. The egg packet presents the distinctive appearance of those belonging to Dipylidium caninum.
Toxocara canis
Nematodes of family Ascarididae; several specimens found in each of TP 1 and TP 23. Spherical, 69–89 μm in diameter, with a pitted or granular surface, and usually yellow/amber/brown in colour, these eggs are consistent with those of Toxocara canis (Figure 1c). A similar species, T. cati, which has smaller eggs (65–75μm diameter), could be eliminated from comparison as there are no native felids in Melanesia.
Unknown trematode 1
One specimen found in TP 23. Roundly oval, 36 μm x 24.7 μm, golden brown, opercular (operculum missing), apparently without opercular shoulders or anopercular knob, or visible surface structure (Figure 1d).
Unknown trematode 2
One specimen found in TP 23. Large, roundly oval, 97 x 82 μm, dark brown, opercular (operculum missing) without opercular shoulders (Figure 1e).
Unknown cestode
Three specimens found in TP 23, round, 53–59 μm in diameter, with a dimpled or punctate outer surface and a neat round hole in one aspect (Figure 1f).
Discussion and Conclusions
Trichuris suis (pig whipworm) was found in TP 23, which also contained pig bones (Summerhayes et al. Reference Summerhayes, Szabo, Fairbairn, Horrocks, McPherson, Crowther, Bedford and Spriggs2019a) (Figure 1a). Pigs (Sus scrofa) were almost certainly introduced to the Anir Islands from Southeast Asia with Lapita (O’Connor et al. Reference O’Connor, Barham, Aplin, Dobney, Fairbairn and Richards2011). Whipworm is a nematode parasite of wild and domestic pigs worldwide, but it is most prevalent in warm, humid climates (Pittman et al. Reference Pittman, Shepherd, Thacker and Myers2010). The life cycle is direct, with no intermediate host, and infection is via the faecal-oral route. Eggs develop over 3–8 weeks in the environment and, once infective, may remain viable for several years. When ingested by a pig, the larva hatches and penetrates the lining of the intestine. Development continues until the adult stage when the thicker posterior section emerges from the intestinal lining and reproduction takes place, the eggs being shed into the intestine for release into the environment via the pig’s faeces. In heavy infections, and when associated with secondary bacterial infections, symptoms may include gastrointestinal problems, weight loss, and death (Pittman et al. Reference Pittman, Shepherd, Thacker and Myers2010). The species is potentially zoonotic, humans becoming infected by ingesting eggs from soil or water. Because there is no migratory stage in the life cycle, the worms remain in the gut and cause little harm to humans.
Dipylidium caninum eggs were found in TP 23 (Figure 1b). A canid tooth was found in TP 23, remains of dogs were found in pits throughout the site, and apparently dogs were locally present from Early Lapita settlement (Summerhayes Reference Summerhayes, Szabo, Fairbairn, Horrocks, McPherson, Crowther, Bedford and Spriggs2019a). At the time of writing, no evidence of dogs has been found in Lapita archaeological sites east of the Bismarck Archipelago (Greig et al. Reference Greig, Gosling, Collins, Boocock, McDonald, Addison, Allen, David, Gibbs, Higham and Liu2018). The eggs are slightly smaller than expected for this cestode (the usual range is 31–50 μm), which we assume to be a factor of preservation conditions. We recognise other possibilities for this egg type, noting that several families of cyclophyllidean cestodes (Anoplocephalidae, Paruterinidae, Nematotaeniidae, and Mesocestoididae) have paruterine organs into which eggs move from the uterus, causing some clustering, similar to that seen in Dipylidium. It is unlikely that any anoplocephalid or mesocestoidid with such features occurred on the archipelago, but the local bird and reptile cestode fauna is not well known and could harbour paruterinids and nematotaeniids. Dipylidium caninum is principally a cestode of dogs and other canids, but the species has zoonotic potential if infected fleas are ingested by humans, modern cases nearly always being reported in young children (Sapp & Bradbury Reference Sapp and Bradbury2020). Dipylidium caninum adults live in the small intestine of the definitive host and release gravid segments into the environment via the host faeces. The segments are full of egg packets, up to c. 100 μm or more in size, which may contain more than 30 eggs, and which are ingested by flea larvae and develop within a cyst in the adult flea. When an infected flea is eaten by a dog, the infective stage is released into the intestine of the host and becomes an adult. This parasite has been recorded from archaeological sites in New Zealand, in a Māori midden on Auckland Isthmus, North Island, and in Māori stone garden systems at Palliser Bay in southern North Island (Horrocks et al. Reference Horrocks, Brown, Brown and Presswell2023a; Horrocks et al. Reference Horrock, Dodd, Bickler, Carley and Presswellin review).
The presence of Toxocara canis, a nematode parasite of dogs and other canids, in TP 23, supports the evidence for commensal dogs in this part of the Bismarck Archipelago (Figure 1c). Toxocara canis inhabits the small intestine of the definitive host and is usually asymptomatic, except in the case of massive infection in pups, which may be fatal. Eggs are deposited in dog faeces and the worm develops within the egg, and once embryonated it becomes infectious. Dogs are infected when they ingest embryonated eggs from contaminated soil, or the worm larvae are directly acquired by a pup trans-placentally. In young dogs the larvae hatch out and migrate to the lungs where they are coughed up and swallowed, bringing them again to the intestine where they mature to adulthood. In dogs over three months old, the larval worm either remains in the gut to mature and produce eggs for dispersal or enters the bloodstream and migrates to any organ of the body, where it will encyst in the tissues (Harris-Linton Reference Harris-Linton2001). Toxocariasis is a zoonotic disease, usually acquired by children ingesting eggs from soil. Infective larvae hatch in the intestine, but the juvenile stages do not mature. Instead, they wander throughout the body for months or up to several years, damaging where they settle, potentially causing blindness or death in the case of ocular or neurological infections. There are currently two palaeoparasitological records of this parasite from New Zealand (Irwin et al. Reference Irwin, Horrocks, Williams, Hall, McGlone, Nichol and Irwin2004; Horrocks et al. Reference Horrock, Dodd, Bickler, Carley and Presswellin review) but none we are aware of from any other part of the southern Pacific region.
Regarding unknown trematode 1 (Figure 1d), this trematode egg belongs to a species from one of the families that produce very small eggs, in superfamilies Opisthorchioidea and Microphalloidea. It does not match the description of any trematodes that are regularly reported to infect humans or those with pig or dog associations (Conlan et al. Reference Conlan, Sripa, Attwood and Newton2011; Esteban et al. Reference Esteban, Muñoz-Antoli, Toledo and Ash2014). The most likely affiliation for this specimen is family Heterophyidae, which uses fishes as intermediate hosts and infects fish-eating birds and mammals, including humans, as adults. The host may have been any bird or mammal that lived or was butchered on the site.
Regarding unknown trematode 2 (Figure 1e), this is a large, operculated trematode egg with a dark, almost opaque shell, possibly embryonated. The size and very round shape (length:width = 1:18) are unusual and do not appear to correspond with expected parasites of host species known from the context.
The unknown cestode egg (Figure 1f) bears a very strong resemblance to those of Stringopotaenia psittacea, found in pre-European contexts in subfossil kākāpō parrot coprolites in New Zealand (Horrocks & Presswell Reference Horrocks and Presswell2023b), and may similarly be from a species of Anoplocephalidae. Species of this family infect herbivorous mammals including marsupials (Beveridge Reference Beveridge1976) and, oddly, parrots (Beveridge Reference Beveridge, Khalil, Jones and Bray1994). Both herbivorous mammals and parrots were represented by bone fragments at Kamgot: a padmelon, reported as Thylogale brunii, and an unnamed psittaciform. According to Flannery (Reference Flannery1995), T. brunii is limited to New Guinea and the Aru Islands, but T. brownii is present in New Ireland (Flannery Reference Flannery1994). The bones found at Kamgot may indicate subsequent local extinction of T. brunii from the Anir Islands or may be a result of past taxonomic confusion with T. brownii (Flannery Reference Flannery1992). The cuscus species Phalanger orientalis and Spilocuscus kraemaeri occur on these islands (Flannery Reference Flannery1994) and both genera are known to harbour anoplocephalid cestodes (Bertiella spp.) (Spratt & Beveridge Reference Spratt and Beveridge2016). Schmidt (Reference Schmidt1972) asserted that Paronia trichoglossi is “known from several kinds of parrot in … New Guinea”. Further speculation on the identification of this egg is not possible as little is known about the parasites of Bismarck Archipelago fauna.
A literature search yields very few records of helminth parasites from the Bismarck Archipelago, precluding meaningful comparison of the parasite fauna found in this study with living species. Our findings of parasites from the Kamgot Early Lapita site support the presence of both pigs and dogs alongside the human population. Summerhayes et al. (Reference Summerhayes, Szabo, Fairbairn, Horrocks, McPherson, Crowther, Bedford and Spriggs2019a) found five definite dog remains and two possible canid remains in the Early Lapita levels at Kamgot, including bones in TP 2, TP 23, and TP 2C, and two teeth in TP 22 and TP 23. DNA analysis and accelerator mass spectrometry on two samples were unsuccessful. Bone sample recovered from above a later Lapita deposit (TP 10) was in a shallow deposit just below the topsoil, and DNA and dating found it to be modern, and therefore from perturbed deposits. These results have left room for ambiguity, and the evidence for the presence of dogs and pigs in Early Lapita contexts and the timing of their arrival in the Bismarck Archipelago has been argued (e.g. Specht et al. Reference Specht, Denham, Goff and Terrell2014; O’Connor et al. Reference O’Connor, Barham, Aplin, Dobney, Fairbairn and Richards2011; Piper Reference Piper, Piper, Matsumura and Bulbeck2017). For example, Specht et al. (Reference Specht, Denham, Goff and Terrell2014) described evidence for pigs during the Early Lapita phase as “ambiguous” because of a lack of direct dating for early remains, including those from Kamgot. (The Kamgot bones were submitted for 14C dating but as they contained no collagen they could not be directly dated.) Specht et al. (Reference Specht, Denham, Goff and Terrell2014) also suggested that contexts assigned to Early Lapita may have accumulated over several hundred years, whereas Summerhayes et al. (Reference Summerhayes, Szabo, Fairbairn, Horrocks, McPherson, Crowther, Bedford and Spriggs2019a) argued that the Kamgot material may have accumulated over a much shorter timescale. A similar position was taken by Specht et al. (Reference Specht, Denham, Goff and Terrell2014) on the evidence for dogs in Early Lapita contexts, which they thought to be “arguable” because of the scarcity of finds. The evidence for the introduction of pigs and dogs was summarised by Matisoo-Smith (Reference Matisoo-Smith and Friedlaender2007) and Anderson (Reference Anderson2009), revealing a complex picture for which dating and sampling questions are critical. Currently, the earliest known dog evidence from mainland New Guinea is from the south-east coast, observed in a recent study where a dog mandible was found in association with Late Lapita material, the latter of which was dated to 2573–2702 cal. BP (Manne et al. Reference Manne, David, Petchey, Leavesley, Roberts, Szabó, Urwin, McNiven and Richards2020).
The origins and arrival of the earliest domestic animals in parts of Oceania has thus been a contested subject that is currently not fully resolved by dating techniques. However, the presence of both dog and pig parasites in this study, found in Early Lapita contexts at Kamgot, confirms that these animals were indeed present at the early stage of Lapita settlement.
Acknowledgements
Summerhayes thanks the communities of Anir Island and the landowners of Kamgot for their permission and support. Thanks also to the National Research Institute of Papua New Guinea (PNG), the National Museum and Art Gallery (NMAG) of PNG, and the University of PNG for their support. Special thanks to the late Herman Mandui, Baiva Ivuyo, and Robert Mondol, all from the NMAG. We thank two anonymous reviewers for improving the paper.

