


Introduction
Exceptional fossil preservation consists of the preservation of non-biomineralized soft parts (for example, digestive and nervous systems of animals) in the geological record. Fossiliferous localities showing this type of preservation are called Konservat-Lagerstätten. The late Tremadocian Fezouata Shale in the Anti-Atlas of Morroco, deposited approximately 480 million years ago, is the only Ordovician period Konservat-Lagerstätte to yield a fully marine diverse exceptionally preserved fauna [Reference Van Roy1]. The ~900 meter thick siltstones of the Fezouata Shale have yielded over 200 taxa of marine invertebrates [Reference Van Roy2]. Most of them belong to arthropods and echinoderms [Reference Van Roy2,Reference Lefebvre3]. In these deposits, exceptionally preserved soft parts (for example, guts) are well-documented in various groups of arthropods (for example, anomalocaridids, trilobites) [Reference Van Roy1,Reference Van Roy2,Reference Van Roy and Briggs4,Reference Gutiérrez-Marco5], annelids [Reference Vinther6], hyolithids [Reference Marti Mus7], molluscs [Reference Vinther8], and palaeoscolecid worms [Reference Martin9,Reference Kouraïss10].
In marked contrast, very few occurrences of exceptionally preserved soft parts have been reported from Fezouata echinoderms: a putative gut was mentioned in one specimen of solutan [Reference Lefebvre11], and more abundant remains (water-vascular system, gut) were described in stylophorans [Reference Lefebvre12]. The water-vascular (or ambulacral) system is a complex, non-biomineralized, coelomic, hydraulic structure, which is unique to echinoderms. It consists of ambulacral canals leading to lateral sets of smaller tube feet, which are used for feeding, locomotion, and respiration.
Such a rarity of exceptionally preserved soft parts in echinoderms from the Fezouata Shale is not surprising. In spite of some recent spectacular reports, for example, in the Silurian Herefordshire Lagerstätte, UK [Reference Sutton13,Reference Briggs14,Reference Rahman15] or the Devonian Hunsrück Slate, Germany [Reference Glass and Blake16], very few examples of soft parts have been described in fossil echinoderms. This situation mainly results from the fast post-mortem disarticulation of their multi-element calcite skeleton and, hence, their low preservation potential, [Reference Lefebvre3,Reference Brett17]. Experimental taphonomy, the study of how organisms decay and become fossilized, on modern echinoderms suggests that both skeletal elements and soft parts disarticulate and degrade within a few days after death [Reference Gorzelak and Salamon18]. Consequently, due to taphonomic biases [Reference Saleh19], the preservation of complete echinoderm specimens yielding soft tissues is extremely rare even within Lagerstätten. Because of these biases, understanding the preservational pathway of an organism is vital prior to any paleontological description especially for animal groups with no current representatives. In this study, we aim at understanding the preservation of eocrinoids and somasteroids from the Fezouata Shale [Reference Lefebvre20,Reference Nohejlová21] and critically evaluate the recent reports of putative soft tissues (water-vascular system) in these two groups.
Material and Methods
The taphonomy of soft parts was analyzed in the two best-preserved specimens of eocrinoid and somasteroid echinoderms from the Fezouata Shale [Reference Lefebvre20,Reference Nohejlová21]. This material is deposited in the paleontological collections of the Musée des Confluences, Lyon, France (acronym “ML”), and Claude Bernard Lyon 1 University, Villeurbanne, France (acronym “UCBL-FSL”), respectively. The eocrinoid specimen (ML20-269159, Reboul collection) was collected at Bou Izargane, about 18 km north of Zagora, Morocco. This locality is a well-known fossiliferous site, where abundant and diverse late Tremadocian exceptionally preserved fossils have been collected (for example, anomalocaridids, marrellomorphs, palaeoscolecids), including trilobites and stylophoran echinoderms with soft parts [Reference Gutiérrez-Marco5,Reference Lefebvre12,Reference Martin22]. The somasteroid specimen (UCBL-FSL 424962, Vizcaïno collection) is from an unknown locality, probably late Tremadocian in age, from the Ternata plain, north of Zagora, Morocco.
The two specimens were characterized using a FEI Quanta 250 scanning electron microscope (SEM) equipped with a backscattered electron detector in addition to an energy-dispersive X-ray analyzer (EDX) at accelerating voltages that varied from 5 to 15 kV. The backscattered electron detector allowed the acquisition of images with dark pixels corresponding to light elements (that is, small atomic number Z) and white pixels corresponding to spots with heavy elements (that is, high atomic number Z). The EDX allowed, at low voltages, optimized acquisition of spectra revealing the distribution of light elements (that is, carbon). At high voltages, heavy elements, such as iron, were detected. In addition, X-ray fluorescence (XRF) elemental maps of these two specimens were obtained using a Bruker M4 Tornado micro-XRF instrument operating under vacuum at 50 kV and 600 µA.
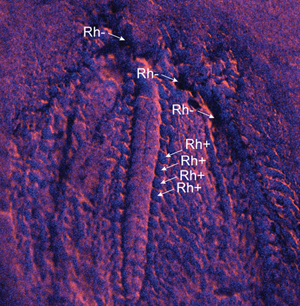
As both specimens were not completely flattened it was difficult to assess if negative elemental results were due to the actual absence of elements in the analyzed spot, or if the negative results were due to a topographic effect that inhibited the beam from reaching the spot. For this reason general elemental maps of rhodium Rh, the source of the XRF machine, were made. When Rh signal was absent it indicated that topographical effects prevented the beam from reaching the spot. Presence of Rh implied that the beam did reach the surface and the absence of an element was due to actual absence of this element from the chemical composition of the analyzed area.
Results
The eocrinoid specimen (Figure 1A) is entirely preserved in iron (Fe), which is shown by the green fluorescence in Figure 1C and the SEM EDX map in Figure 1E. This contrasts with the matrix, which is rich in silicon (Si) as shown by the red fluorescence in Figure 1B and the SEM EDX map in Figure 1E. Some Fe-rich small and repetitive structures are present within the brachioles (skeletonized arm-like feeding appendages) (Figure 1D). Most of the Fe-rich areas are preserved in small iron crystals that do not have any specific shape (that is, star-like minerals that are not euhedral nor framboidal; Figures 2A–D).

Figure 1: Analyzed specimen of eocrinoid echinoderm, late Tremadocian, Fezouata Shale, Bou Izargane, Zagora area, Morocco; ML20-269159. (A) Photograph of the specimen; (B) X-ray fluorescence image of inset shown in Figure 1A. Red indicates a high concentration of silicon distribution; (C) X-ray fluorescence of iron distribution (green) from the inset region of Figure 1A; (D) tube feet-like structures of the putative water vascular system showing iron distribution; (E) SEM-EDX elemental distribution in the matrix and the fossil.

Figure 2: SEM-EDX images showing amorphous shape of particles and iron-rich areas (white regions).
The somasteroid specimen (Figure 3A) is preserved as an imprint in the rock, and it shows the same elemental signature as the matrix, which is rich in Si (Figure 3B). However, some areas of the fossil appear to be depleted in Si (Figure 3B). Rh distribution shows that some of these anomalies are due to the actual absence of Si (that is, when Rh is present Rh+; Figure 3C), while the absence of Si in other regions is simply due to a hidden zone that was not analyzed by the beam (that is, Rh-; Figure 3C). The Rh+ zones are rich in carbon that is mostly detected when analyzing the specimen at 5 kV (Figure 3D). The carbon shows a repetitive pattern of small identical structures along the arms of the somasteroid (Figures 4A–B). However, carbon is also present elsewhere in the specimen (Figures 4C–F).

Figure 3: Analyzed specimen of somasteroid, late Tremadocian, Fezouata Shale, unknown locality, Zagora area, Morocco; UCBL-FSL424962. (A) Photograph of the specimen; (B) XRF, Si distribution; (C) XRF, superposition of Rh (blue) with K; (D) SEM-EDX elemental distribution in the tube feet-like structures.

Figure 4: (A–B) Carbon particles (black) in tube feet-like structures of the putative water vascular system, (C–F) carbon particles found in other random structures showing the same carbon distribution as tube feet-like structures.
Interpretation and Discussion
The structures found in both the eocrinoid (Figure 1D) and somasteroid (Figures 4B–C) specimens resemble tube feet of the water vascular system, as, for example, those evidenced in stylophoran echinoderms from the Fezouata Shale [Reference Lefebvre12]. However, both the elemental and mineralogical signatures are different in these three occurrences.
Soft parts in exceptionally preserved fossils from the Fezouata Shale are preserved in both euhedral and framboidal pyrite [Reference Saleh23]. These mineralogical morphologies are indicative of an early authigenic pyritization that occurred under anoxic conditions at time of burial [Reference Vaucher24], replicating soft tissues that are the most prone to decay [Reference Gabbott25]. After that, pyrite was transformed due to recent weathering to iron oxides [Reference Lefebvre12,Reference Saleh23]. This situation was observed in the soft parts reported in the stylophoran material from Bou Izargane in Morocco [Reference Lefebvre12].
In contrast, the near absence of both euhedral and framboid minerals in the eocrinoid specimen indicates that iron-rich minerals that are found in this fossil are not the result of the combined activity of authigenic mineralization and recent weathering. In this fossil, the iron oxides are simply related to the activity of recent weathering and, thus, they do not replicate any original anatomy. The distribution of these Fe-rich minerals in all anatomical parts of the specimen, including skeletal plates (Figure 1C), validates this hypothesis.
The somasteroid specimen shows a totally different elemental distribution in the tube feet-like structures, with the total absence of Fe and the presence of carbon. Carbonaceous films are the main mode of preservation of soft tissues, especially in the Paleozoic era [Reference Gaines26]. However, in surface sediments of the Fezouata Shale, carbon is absent from all fossils that have been analyzed [Reference Martin9,Reference Kouraïss10,Reference Lefebvre12]. The porous aspect of C-rich deposits in the analyzed specimen is strikingly different from the texture of carbon films in exceptionally preserved biotas [Reference Gaines26]. Moreover, in the somasteroid specimen, carbon is not limited to any specific region but appears to fill all micro-depressions of the specimen (Figures 4D–G). Thus, it is very likely that this carbon is the consequence of preparation artifacts: numerous latex casts were made to study the detailed morphology of this specimen. Apparently, latex was not completely removed from small cavities located between skeletal plates, thus mimicking regularly branching tube feet along the ambulacral canal, in the arm region of the analyzed specimen.
Conclusion
This study confirms the validity of the interpretation as soft parts of the structures identified in stylophoran echinoderms from the Fezouata Shale [Reference Lefebvre12]. However, it also shows that it is probable that reports of putative soft parts in eocrinoids and somasteroids from the Fezouata Shale [Reference Lefebvre20,Reference Nohejlová21] are either preparation artifacts or the result of recent weathering. In addition, it shows that understanding the history of a specific fossil, from the excavation to preparation, is essential for proper paleontological interpretation, especially when it comes to the description of soft tissues in extinct taxa. These results should be taken into consideration when studying any new fossil, whether it is purchased from a private collector or comes from a museum collection.
Acknowledgements
This paper is a contribution to the TelluS-INTERRVIE projects “Mécanismes de préservation exceptionnelle dans la Formation des Fezouata” (2018), and “Géochimie d'un Lagerstätte de l'Ordovicien inférieur du Maroc” (2019), all funded by the INSU (Institut National des Sciences de l'Univers, France), CNRS. This paper is also a contribution to the International Geoscience Programme (IGCP) Project 653 – The Onset of the Great Ordovician Biodiversification Event. The authors are grateful to the LABEX Lyon Institute of Origins (ANR-10-LABX-0066) of the Université de Lyon for its financial support within the program “Investissements d'Avenir” (ANR-11-IDEX-0007) of the French government operated by the National Research Agency (ANR). The authors thank Stefan Lalonde and Pierre Sansjofre (Brest University) for assistance during SEM and XRF analyses. The Editor-in-Chief, Bob Price, is thanked for his helpful and constructive remarks.




