



Introduction
Tropical forests in South-east Asia have high levels of species richness and endemism (Kingston, Reference Kingston, Liat and Akbar2010), with four of the world's 25 biodiversity hotspots found in the region (Myers et al., Reference Kuo, Soisook, Ho, Csorba, Wang and Rossiter2000). However, South-east Asia, including parts of Myanmar, also has one of the highest rates of deforestation, habitat destruction and degradation (Keenan et al., Reference Keenan, Reams, Achard, Freitas, Grainger and Lindquist2015; Curtis et al., Reference Curtis, Slay, Harris, Tyukavina and Hansen2018; Estoque et al., Reference Estoque, Ooba, Avitabile, Hijioka, DasGupta, Togawa and Murayama2019). Approximately 80 million ha of forest were lost during 2005–2015, including nearly 4 million ha in Myanmar (Estoque et al., Reference Estoque, Ooba, Avitabile, Hijioka, DasGupta, Togawa and Murayama2019). In the Philippines and parts of Indonesia, it is projected that 98% of forests will be lost by 2022 (Hughes, Reference Hughes2017). In Viet Nam, forests are becoming more fragmented and degraded, and closed-canopy forests constitute only 4.6% of the total forested area (WorldBank, 2005). Rapid land-use changes mean that much of the region's fauna is threatened (Kingston, Reference Kingston, Liat and Akbar2010) and the density of threatened vertebrates is amongst the highest in the world (Hughes, Reference Hughes2017). It has been estimated that the region, which includes 20 hotspots of extinction risk (Cardillo et al., Reference Cardillo, Mace, Gittleman and Purvis2006), could lose up to 42% of its biodiversity by 2100 (Sodhi et al., Reference Simmons and Cirranello2004).
With an area of 11,280 km2 (Suarez-Rubio et al., Reference Suarez-Rubio, Connette, Kyaw, Meyers, Thaw and Renner2020), the Hkakabo Razi Landscape is one of the largest remaining tracts of mainly intact forest in South-east Asia (Bhagwat et al., Reference Bhagwat, Hess, Horning, Khaing, Thein, Aung and Aung2017). It is located along the south-eastern slopes of the eastern sub-Himalayan mountain range and comprises Hkakabo Razi National Park, the Hponkan Razi Wildlife Sanctuary and the proposed southern extension of Hkakabo Razi National Park (Fig. 1; Suarez-Rubio et al., Reference Struebig, Christy, Pio and Meijaard2020).

Fig. 1 The Hkakabo Razi Landscape in northern Myanmar, including the 11 capture sites of 2016.
The forests of Hkakabo Razi have high structural integrity and experienced low rates of deforestation during 1989–2016 (Suarez-Rubio et al., Reference Struebig, Christy, Pio and Meijaard2020). During 1991–1999, the annual deforestation rate was < 0.2%, concentrated on the plains of Putao and Naung Mung (Renner et al., Reference Rao, Saw, Platt, Tizard, Poole, Than and Watson2007). The forests are situated at the meeting point of three biodiversity hotspots: Indo-Burma, Himalaya and Mountains of South-west China (Myers et al., Reference Kuo, Soisook, Ho, Csorba, Wang and Rossiter2000). Bird species richness is high (Renner & Rappole, Reference Renner, Rappole, Leimgruber, Kelly, Shwe, Aung and Aung2011; Renner et al., Reference Renner and Rappole2015), with 40% of Myanmar's c. 1,100 bird species occurring in the area (Renner et al., Reference Renner and Rappole2015). In 2014, the Hkakabo Razi Landscape was proposed as a World Heritage Site under criteria (ix) and (x) for its high integrity and outstanding ecological values (World Heritage Centre, 2014).
Although the diversity of birds and larger mammals in the Hkakabo Razi Landscape is relatively well known (Rao et al., Reference Myers, Mittermeier, Mittermeier, da Fonseca and Kent2013; Renner et al., Reference Renner and Rappole2015), this is not the case for most other taxa. Only two previous surveys included bats: by the Earl of Cranbrook and Kingdon-Ward in 1931 and by Kaulback in 1933. Seventeen specimens of four species were collected during these surveys (Hill, Reference Hill1962). Elsewhere in Kachin State bat research was mostly conducted during 1931–1945 (Bates et al., Reference Bates, Tin Nwe, Pearch, Swe, Bu and Tun2000) and in the early 21st century (Bates et al., Reference Bates, Struebig, Rossiter, Kingston, Oo and Mya2004; Struebig et al., Reference Struebig, Kingston, Zubaid, Mohd-Adnan and Rossiter2005). None was extensive and only one involved harp-traps and mist-nets (Struebig et al., Reference Struebig, Kingston, Zubaid, Mohd-Adnan and Rossiter2005).
Bats are recognized as a critical component of South-east Asia's fauna, comprising 30% of the region's mammal species and 50% of the mammal species within the tropical rainforest ecoregions (Kingston, Reference Kingston, Liat and Akbar2010). South-east Asia is home to 27% of global bat diversity (385 of the currently recognized 1,411 bat species; Simmons & Cirranello, Reference Schnitzler and Kalko2020). Nearly half of the region's bat species are predicted to be extinct by the end of the 21st century, and forest-dependent species are particularly vulnerable (Kingston, Reference Kingston, Liat and Akbar2010, Reference Kingston2013; Frick et al., Reference Frick, Kingston and Flanders2019). As Kingston (Reference Kingston, Liat and Akbar2010) noted, this is a catastrophic decline, including the loss of the many ecosystem services that bats provide.
Study area
The mountain ranges of the Hkakabo Razi Landscape encircle the Putao Plains, except to the south, where lowlands give way to the headwaters of the Ayeyarwady River (Fig. 1). The altitude ranges from 450 m in the Putao Plains to 5,881 m at the summit of Mount Hkakabo Razi. Except for dry deserts, all major vegetation types are present. The area is predominantly covered by different forest types, tropical to subtropical (semi-deciduous) from lowlands to alpine zones. Above the alpine vegetation, rock/boulder and glaciers/snow fields occur (Renner et al., Reference Renner and Rappole2015). Settlements and agriculture are restricted mainly to the flood plains of Putao and Naung Mung (Renner et al., Reference Rao, Saw, Platt, Tizard, Poole, Than and Watson2007). In some areas, swidden cultivation has created a patchwork of secondary forest in various stages of regeneration. The Hkakabo Razi Landscape is home to >8,000 people, and except for Putao and Naung Mung townships, virtually no other settlement exceeds 100 inhabitants (Suarez-Rubio et al., Reference Struebig, Christy, Pio and Meijaard2020).
Methods
We undertook fieldwork during 28 nights at 11 sites (Table 1), in the southern margins of the Hkakabo Razi Landscape at altitudes of 450–1,220 m (Fig. 1), using four-bank harp-traps and mist-nets, and conducting roost surveys and acoustic surveys (Supplementary Table 1). Wherever possible, we used a suite of three capturing methods to maximize success. Four-bank harp-traps were set in forests, across trails and over streams, and checked during 18.00–21.00 and again at dawn. This resulted in a total of 66 harp-trap nights, with one trap-night defined as one trap set per night. Mist-nets (70 denier nylon, 30 × 30 mm mesh size; Ecotone, Gdynia, Poland) were also set along and across forest trails and across streams. They were opened during 18.00–21.00 and checked regularly. In addition, we employed some canopy nets; in Tana cave we used mist-nets and hand nets. Mist-net hours, including canopy nets, totalled 718. We conducted transect and point acoustic surveys with a Pettersson D1000x bat detector (Pettersson Elektronik, Uppsala, Sweden) and analysed echolocation calls with BatSoundPro4.1 (Pettersson Elektronik, Uppsala, Sweden). We identified bat calls to species level using the call database of the Princess Maha Chakri Sirindhorn Natural History Museum (P. Soisook, unpubl. data; Hughes et al., Reference Hughes, Satasook, Bates, Soisook, Sritongchuay, Jones and Bumrungsri2010; Hughes et al., Reference Hughes, Satasook, Bates, Soisook, Sritongchuay, Jones and Bumrungsri2011; Francis, Reference Francis2019).
Table 1 Sampling effort by method at 11 capture sites in the Hkakabo Razi Landscape in 2016.

1 Number of households within community borders of capture sites.
We took 62 non-lethal tissue samples from 27 species for molecular analysis, and 19 voucher specimens of 13 taxonomically ambiguous, small vespertilionid species. We estimated the expected species numbers S based on the spatial effort (capture sites) for the three detection methods, treated separately and combined. We included one unidentified Murina species and one unidentified Myotis species in the analysis. In addition to estimating the expected species numbers for 10 capture sites (Tana cave was excluded from this particular analysis), we extrapolated the expected species numbers for a hypothetical 30 capture sites (for all combined methods and for acoustic/sight sampling) in EstimateS 9.1.0 (Colwell, Reference Colwell2013), and for 28 harp-traps and mist-netting capture sites, to examine the difference between observed diversity and theoretically expected diversity. To assess the effort per method, we established a matrix for all detected species with frequency indications per method employed and per sites where they were encountered. The estimator S indicates that our effort would have been maximized with 28 capture sites for harp-trapping and 30 capture sites for all other methods.
Results
We recorded a total of 203 bats of 35 species in six families in the Hkakabo Razi Landscape (Supplementary Table 1), of which two have been described as new species (Murina hkakaboraziensis and Kerivoula furva) and five are new species records for Kachin State, including two that are also new for Myanmar (Kuo et al., Reference Kingston, Francis, Akbar and Kunz2017; Soisook et al., Reference Sodhi, Koh, Brook and Ng2017). The Vespertilionidae with eight genera and 19 species were the most diverse, with 54.3% of all species recorded (Supplementary Table 1). However, with 64 individuals (31.5% of captures) their abundance was relatively low. Fruit bats (Pteropodidae) accounted for only 11.4% of total diversity but 28.6% of captures. With 38 captures, Cynopterus sphinx was the most commonly captured species. The rank-abundance curves indicate a typical pattern of a few species with many individuals and many species with few individuals (Fig. 2).

Fig. 2 Species rank-abundance curve based on all capture methods combined, for different habitat types and elevation (median elevation) for bats surveyed in the Hkakabo Razi Landscape in 2016.
The rarefaction curve suggests that if we had tripled our effort and increased the number of capture sites from 11 to a hypothetical 30, we could have expected 48.8 ± 7.8 species, an increase of c. one-third. This is based on the three detection methods combined (audio recording, harp-traps and mist-nets; Fig. 3).
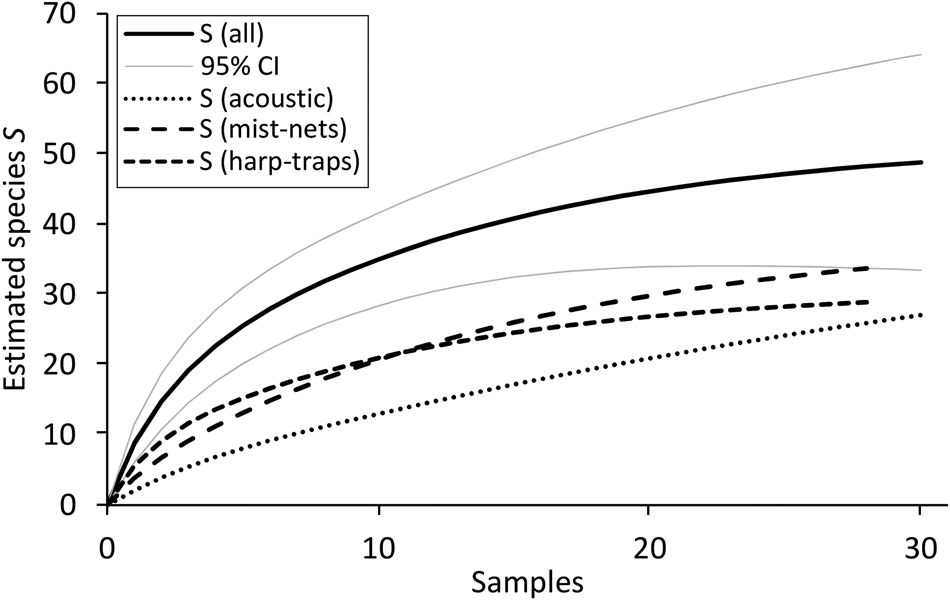
Fig. 3 Species rarefaction curve for all capture methods with 95% confidence intervals and for 30 hypothetical capture sites, additionally separated for the three capture methods (base: ten capture sites used for all methods and acoustic survey; eight sites for all mist-nets and harp-traps).
Of the 27 species captured in harp-traps and mist-nets, nine were only captured in harp-traps and eight were only captured in mist-nets (Supplementary Table 1). Of the 13 species detected by acoustic/sight surveys, eight were not captured in mist-nets or harp-traps. We recorded six species in Tana cave, the only limestone cave encountered; four of these were captured in mist-nets; two species (Taphozous theobaldi and Hipposideros armiger) were found nowhere else (Supplementary Table 1).
Our surveys up to 645 m (the median elevation) resulted in 22 species and 111 records; at altitudes of 645–1,220 m, we recorded 92 individuals of 31 species (Supplementary Table 1). Of the 203 individual bat records, 130 (64.0%), representing 25 species, were captured within or near the edge of intact forest. All other species were from secondary forest, settlements or shrub.
Discussion
Our survey of bats in the Hkakabo Razi Landscape provides a baseline species list for an area nominated as a World Heritage Site. It increases the number of species known from this area from four (Hill, Reference Hill1962) to 36, the number known from Myanmar from 100 to 104 species (Bates et al., Reference Bates, Tun, Aung, Lu, Lum and Sein2015; Dar et al., Reference Dar, Kamalakannan, Venkatraman and Chandra2019; Francis, Reference Francis2019; Simmons & Cirranello, Reference Schnitzler and Kalko2020), and the number known from Kachin State (89,041 km2) from 38 to 56 (Fig. 4), which is 14.5% of South-east Asia's 385 bat species. In a global context, this exceeds the 47 species recorded from North America (Simmons & Cirranello, Reference Schnitzler and Kalko2020), with an area of 24.71 million km2.
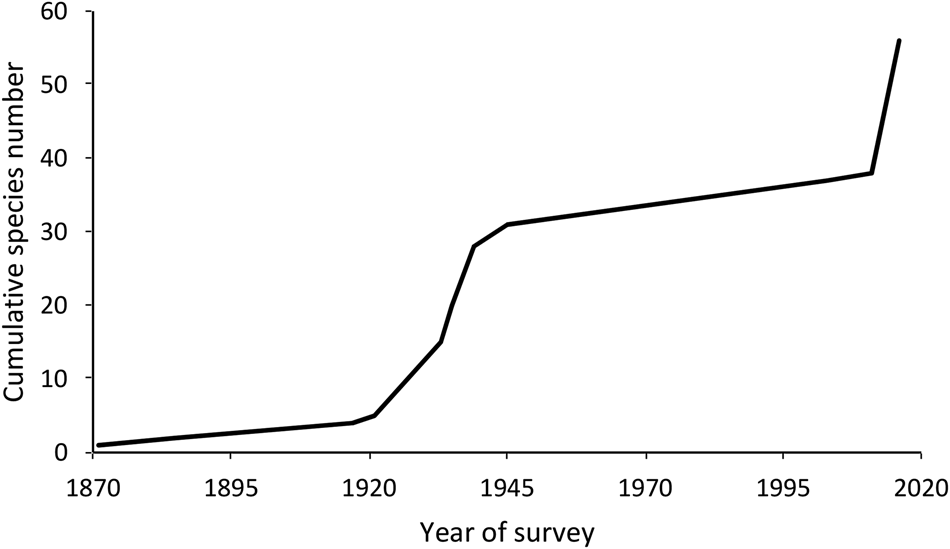
Fig. 4 Cumulative number of bat species recorded from Kachin State, Myanmar (1871–2016), including the number of new species for Kachin from each survey period.
However, it is not only the number of species recorded in the Hkakabo Razi Landscape that is remarkable but also the species composition, which indicates that the area is an important refuge for forest-interior specialist bats. We recorded 12 species, belonging to two subfamilies, the Kerivoulinae (Kerivoula, Phoniscus) and Murininae (Murina). Many of these taxa are little-known and have been discovered and described only recently. Two are categorized on the IUCN Red List (IUCN, 2020) as Data Deficient, five as Least Concern, four have yet to be assessed, and one is only identified to genus (Supplementary Table 1). Forest-interior specialist bats are a priority for chiropteran conservation, being most at risk from habitat change. They have strong site fidelity, are less geographically mobile than nomadic generalist bats such as Cynopterus sphinx, and have smaller home ranges that do not extend beyond forest boundaries (Struebig et al., Reference Son, O'Shea, Gore, Csorba, Tu, Oshida, Endo and Motokowa2008; Huang et al., Reference Huang, Rustiati, Nusalawo and Kingston2019). Compared to cave-roosting bats, they are more susceptible to microclimate and habitat changes and more vulnerable to loss of roosting sites, which include hollows and cavities of standing and fallen trees (Struebig et al., Reference Son, O'Shea, Gore, Csorba, Tu, Oshida, Endo and Motokowa2008). Furthermore, they have eco-morphological adaptations, which constrain their ecological flexibility (Furey et al., Reference Furey, Mackie and Racey2010). These include specialized wing morphology (low wing loading, low aspect ratios), which restricts them to hunting in environments with dense vegetation (Schnitzler & Kalko, Reference Renner, Rappole, Milensky, Aung, Shwe and Aung2001), and specialized acoustic characteristics (short duration, low intensity but very high frequency calls), which allow them to glean their arthropod prey in narrow spaces but is ill-suited for prey detection in more open habitats (Kingston et al., Reference Kingston, Adams and Pedersen2003).
Baseline data for the Hkakabo Razi Landscape contribute to our understanding of the ecology, diversity, composition, assemblages, and the natural spatio-temporal changes of the bat fauna in an intact eastern Himalayan forest. This will help to devise effective conservation strategies. With further, more temporally and spatially widespread research within Hkakabo Razi, these data will complement those obtained elsewhere in the Indo-Chinese subregion and from South-east Asia's Sundaic lowland forest bats, such as from Krau Wildlife Reserve in peninsular Malaysia. Krau is considered home to ‘the highest diversity of bats recorded anywhere in the Old World tropics’ (Kingston et al., Reference Kingston, Liat and Akbar2009, p. 11), with > 70 bat species, including 14 species of Kerivoulinae and Murininae, recorded during 1980–2008 (Struebig et al., Reference Son, O'Shea, Gore, Csorba, Tu, Oshida, Endo and Motokowa2008).
Apart from Krau, the Hkakabo Razi forests have one of the highest recorded diversities of forest-interior specialist bat species in the Murininae and Kerivoulinae. In Bukit Barisan Selatan National Park, one of the last refuges of intact forest in Sumatra, of a total of 60 bat species, 11 were forest-interior specialists in these subfamilies (Huang et al., Reference Huang, Jazdzyk, Nusalawo, Maryanto, Maharadatunkamsi, Wiantoro and Kingston2014). In the disturbed forests of Kim Hy Nature Reserve, Viet Nam, which includes extensive limestone karst, 42 species were recorded, but the diversity of forest-interior bats is considerably lower, with five Murininae and two Kerivoulinae (Furey et al., Reference Furey, Mackie and Racey2010). In the watershed protection forest in the south-eastern Truong Son (Annamite Mountains), 20 species were recorded, including six forest-interior bats, one Kerivoulinae and five Murininae (Son et al., Reference Soisook, Thaw, Kyaw, Lin Oo, Pimsai, Suarez-Rubio and Renner2016). Although other areas in Viet Nam have a bat diversity comparable to that of the Hkakabo Razi Landscape (Put Mat with 39 species, Cuc Phuong with 38 species and Phong Nha with 32 species), the number of forest specialists is lower. However, most of the surveys there were undertaken prior to the use of harp-traps (Hendrichsen et al., Reference Hendrichsen, Walston, Bates and Hayes2001). Our findings from Hkakabo Razi reinforce the view that large forest tracts should be conservation priorities in landscape-level planning because they support rare, forest specialist species (Struebig et al., Reference Struebig, Rossiter, Bates, Kingston, Oo, Nwe and Aung2010).
Although our survey was short and geographically restricted, it indicates that the Hkakabo Razi Landscape has a highly diverse bat fauna and is a conservation priority for bats in South-east Asia. As one of the last remaining extensive tracts of mainly intact forest in Asia, it fully deserves the protection that could be provided through a listing as a World Heritage Site.
Acknowledgements
We thank the Director General of the Forest Department in the Ministry of Nature Conservation and Environmental Protection, U Nyi Nyi Kyaw and Deputy Director U Naing Zaw Htun for permission to study; UNESCO for financial support (grants 4500291033, 7833010728); Koen Myers for discussion on World Heritage; the staff of Hkakabo Razi National Park, particularly U San Naing Dee for logistical support; U Aung Kyaw for organizing the trip in 2016; and U San Lwin Oo, Dee Shin, Htin, Hdoa Dee and numerous porters, cooks and assistants for their help during the expeditions.
Author contributions
Study design: SCR, PJJB; fieldwork: PS, SSLO, AP; data analysis: MSR, AD, PS, SSLO, AP; writing: PJJB; revisions: all authors.
Conflicts of interest
None.
Ethical standards
This work abided by the Oryx guidelines on ethical standards. The Nature and Wildlife Conservation Division of the Forestry Department (NWCD) endorsed the study and granted permission to capture bats and access the protected areas under a contract with UNESCO (Phase II 504MYA4001). All voucher specimens were taken in accordance with national Myanmar law and the animal protection laws of the EU. All procedures were approved by the Nagoya Protocol process, represented by MoNREC NR-2/2-2017.








 Open access
Open access