



Introduction
Gerbera (Gerbera jamesonii Bolus ex Hooker F.) is a perennial ornamental herb that belongs to the family Asteraceae. It is a diploid species with somatic chromosome number 2n = 50 and originates from South Africa, Madagascar and tropical Asia. The modern Gerbera arose from G. jamesonii hybridising with G. viridifolia and possibly other species. It is commonly known as the Transvaal Daisy, Barberton Daisy or African Daisy. It is one of the most important commercial cut flower crops in the global floricultural industry and occupies the 5th position in the international flower trade (Rymbai et al., Reference Rymbai, Jha and Talang2021). In India, it ranks 6th among commercial flowers and occupies an area of 1150.05 ha with a production of 25,554.76 MT (NHB, 2016). The flower of Gerbera, called the head or capitulum, features prominent ray florets in one or two whorls. It is suitable both for export and domestic markets due to its durability for prolonged transportation. Gerbera produces excellent flower heads with exquisite shape, size and vibrant colours. Flowers contain varieties of yellow, orange, cream, white, red, yellow and pink colours as visually indicated by the Royal Horticultural Society Colour Chart (6th edition, 2015) (Rymbai et al., Reference Rymbai, Jha and Talang2021). Cut flowers are in great demand for presentation and interior decoration and are widely used in bouquets and flower arrangements. It is also highly suitable for growing in beds, borders, pots and rock gardens. However, the suitability of gerbera plants for different utility varies with varieties, according to their flower types and growth habits. The durability of cut flowers, their attractive geometrical shape and the wide variety of flower colours are the main morphological traits that attract breeding attention. Gerbera plants are propagated through the division of suckers (clumps) and tissue culture plants for the commercial production of flowers. The propagation method through suckers is cheap, hardy, has a low mortality rate on transplanting, and produces early flowering within two months after planting (Rymbai et al., Reference Rymbai, Jha, Ngachan, Shimray and Kumar2015).
Gerbera is mostly grown under protected conditions for year-round production. However, little is known about commercial Gerbera cultivation under open field conditions (Rymbai et al., Reference Rymbai, Jha, Ngachan, Shimray and Kumar2015). The mild and favourable agro-climatic conditions in the mid-hills of the north-eastern region of India have been found to be appropriate for commercial cultivation of Gerbera under open field conditions. This has the benefit of lower production costs since the expense of production and planting materials under protected conditions is very high (Rymbai et al., Reference Rymbai, Jha, Ngachan, Shimray and Kumar2015, Reference Rymbai, Jha, Talang, Assumi, Deshmukh and Roy2017). There is, however, little information available on cultivars suitable for open cultivation of Gerbera. To address this, a breeding programme was initiated at ICAR Research Complex for NEH Region, Umiam, Meghalaya, to develop cultivars and hybrids suitable for open field conditions with quality flowers and a high propensity for sucker production. The flowers of these gerbera hybrids have a good colour appealing as characterised by their hue, saturation, brightness, structural colours (gloss, polarisation, iridescence), and also by their contrast with surrounding colour (van der Kooi et al., Reference van der Kooi, Elzenga, Staal and Stavenga2016). To breed a new variety with desirable traits, knowledge of heritability, genetic divergence and genetic correlation are crucial genetic parameters for quantitative traits that enable prediction of the response to selection. Mahalanobis’ distance (D2) statistic is an effective tool for estimating genetic divergence and permits precise comparison among all possible pairs of populations in any group before affecting actual crosses (Mahalanobis, Reference Mahalanobis1928). Besides aiding in the selection of divergent parents for hybridisation, D2 statistics also estimate the degree of diversification and determine the relative contribution of each component character to the total divergence. Further, principal component analysis (PCA) is a statistical technique used for the identification of characters that contribute significantly to the total variation and to eliminate duplicate genotypes with similar characteristics (Singh et al., Reference Singh, Prakash, Chakraborty, Wheeler, Agarwal and Ghosh2016). The present investigation was undertaken to study the genetic parameters and diversity among the accessions, the correlation in quantitative traits for further utilisation in a breeding programme, and the identification of suitable genotypes for open conditions.
Materials and methods
Experimental site
The experiment was conducted every year, including all 37 accessions, for a consecutive crop of five years (2013–2018) in ICAR Research Complex for NEH Region, Umiam, Meghalaya. The study area is situated at 250 41' N latitude, 91° 55' E longitude and 1010 m altitude above mean sea level. The experiment site is clay loam to sandy clay loam. Soil is acidic in nature with pH 4.9, P-deficient acid alfisols, SOC (1.77%), exchangeable Al+++ (148.6), Ca++ (240.5), Mg++ (120), K + (66.7), Bray's P2-P (1.2) and available B (0.9 mg kg–1).
Weather characteristics
The climatic conditions at the experimental site are humid and subtropical. A mean maximum temperature of 26.16°C and mean minimum (14.0°C) were recorded during the study period, with the maximum temperature recorded during the month of July (28.57°C) and the minimum in January (5.68°C). Total annual rainfall of 2273.8 mm, with more than 90% falling during April to October; maximum rainfall was received during August (495.8 mm) and minimum during December (7.1 mm). Relative humidity (RH) ranged between 77.18–88.12% and 46.00–75.70% for maximum and minimum RH, respectively.
Experimental design
The study consists of 37 accessions including standard check Alesmera and CSA Collection were evaluated at Horticulture Farm, ICAR Research Complex for NEH Region, Umiam, Meghalaya (Table S1). The experimental design was entirely randomised block design, with 37 treatments (accessions/genotypes) and replicated three time for each accession. Planting was done at a spacing of 30 × 30 cm in a raised bed of 30 cm height and size of 1.5 × 1.5 m under open field conditions. Each replication consisted of 25 uniform sized (4–5 leaves) clumps (suckers) of each genotype were planted during August. Cultivation methods were followed as per standard procedures (Rymbai et al., Reference Rymbai, Jha, Ngachan, Shimray and Kumar2015).
Observations
Vegetative characteristics
The observations for vegetative characters such as number of leaves, leaf length, leaf width and plant spread were recorded from randomly selected ten plants in each replication at full bloom stage (120 days after planting). Plant spread was measured at the maximum width of the plants in two directions, i.e. East-West and North-South, and the average of the two directions was expressed in cm.
Flower characteristics
Flower attributes were recorded at the full bloom stage. The number of days taken from flower bud (pea size) appearance until buds breaking was estimated as days to bud burst and till the opening of the floret as days to flower opening. Flower stalk length was measured from base to neck of the stalk, and diameter was measured in the mid portion of the stalk using a digital vernier calliper (INSIZE SL-A1108-150). The flower head diameter was measured across two extreme points of the fully developed capitulum.
Yield and vase life
The number of flowers per plant was recorded for six months (November – April) and the number of suckers was taken during the month of August at the time of uprooting. Flowers were harvested early in the morning when the outer florets were fully opened and perpendicular to the stalk. The flowers were precooled for 1 h at 5°C, and then the stalks were cut prior to placing them in a container holding tap water at room temperature. Vase life was recorded from the day of harvesting until more than 1/3rd of the outer petals of the flowers turned brown/ wilted.
Data analysis
The data of each year during the five-year experiment (2013–2018) were pooled and subjected to statistical analysis in triplicates, and the data of ten plants from each replication was presented as the mean using one-way ANOVA (p ≤ 0.05). Phenotypic and genotypic variances of the genotypes were estimated as described by Burton and Devane (Reference Burton and Devane1953), heritability as described by Hanson et al. (Reference Hanson, Robinson and Comstock1956), and genetic advance was estimated using the formula suggested by Johnson et al. (Reference Johnson, Robinson and Comstock1955). The genotypic and phenotypic correlation coefficient and path coefficient were estimated as described by Dewey and Lu (Reference Dewey and Lu1959). The genetic distance between the genotypes was worked out using Mahalanobis’ D2 analysis (1928), and the grouping of varieties into clusters was done following the Tochers method suggested by Rao (Reference Rao1952).
Results
Genetic parameters for growth and floral traits
The result indicated a considerable range of differences with respect to phenotypic, genotypic and environmental coefficients of variation (Table 1). The phenotypic coefficients of variation (PCV) were higher than their corresponding genotypic coefficients of variation (GCV) for all the characters under consideration. Amongst all the characters, the highest GCV and PCV were recorded for the total number of leaves per plant (56.54, 55.66) followed by disc diameter (37.44, 36.36) and plant spread (25.90, 23.91). A high estimate of broad-sense heritability (H2) was also obtained for the number of leaves (96.91), flower disc diameter (94.30), number of flowers (90.11), flower stalk length (86.59) and plant spread (85.18). All the traits have shown high heritability (>60%) with high genetic advance (GA, >20%), with the exception of days to flower opening, stalk diameter and flower diameter.
Table 1. Genetic parameters for growth and floral traits of 37 genotypes of gerbera
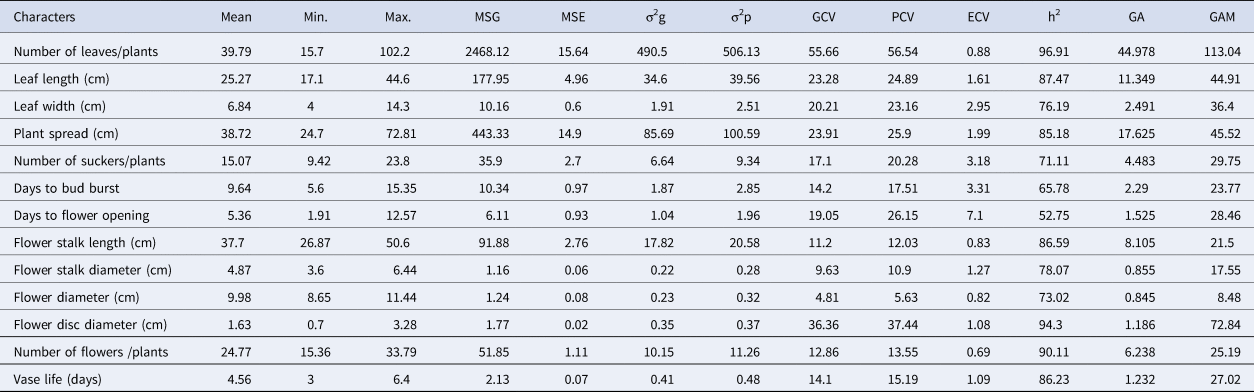
MSG, mean square of genotypes; MSE, error mean square; σ2g,=genotypic variance; σ2p,=phenotypic variance; PCV, phenotypic coefficients of variation; GCV, genotypic coefficients of variation; ECV, genotypic coefficients of variation; h2, Broad sense heritability; GA, Genetic advance; GAM, genetic advance as percentage of the mean.
Cluster (D2) analysis
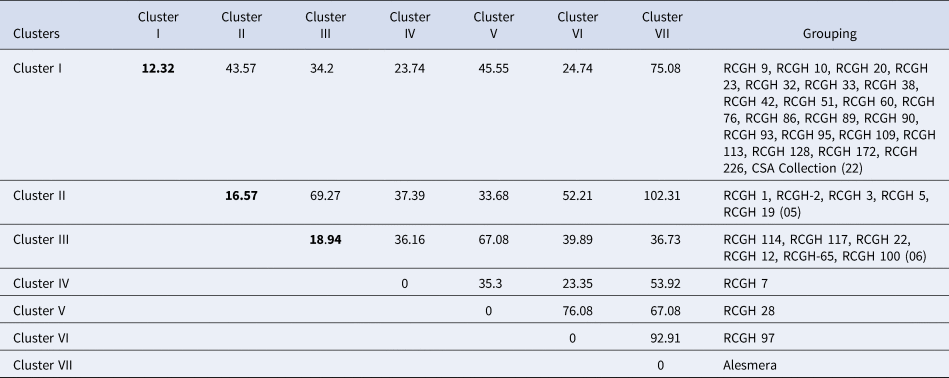
Analysis as per cluster D2 resulted in the grouping of all 37 accessions into seven clusters (Table 2). The maximum number of genotypes (22) were grouped into cluster I, followed by cluster II (05 genotypes) and cluster III (06 genotypes), whereas clusters IV, V, VI and VII were each monogenotypic. The inter-cluster D2 values ranged from 23.35 to 102.31, wherein the lowest inter-cluster D2 value was recorded between clusters IV and VI and the highest was recorded between clusters II and VII. The monogenetic cluster V (RCGH 28) had the unique trait of dwarfing flower stalks in nature (28.33 cm).
Table 2. Inter - and intra- (bold) cluster distances (D2) and distribution of 37 genotypes of gerbera to different clusters
Contribution and mean clusters of individual traits
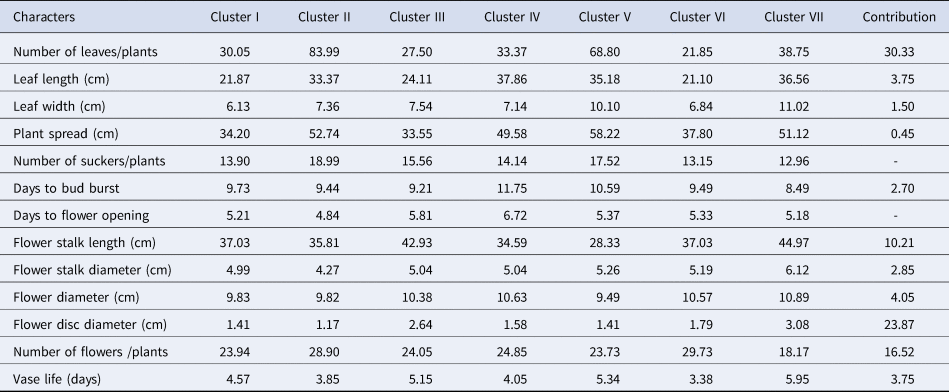
Table 3 revealed the divergence due to the contributions and mean clusters of specific traits. The results showed that among various traits, number of leaves contributed the most towards genetic divergence (30.33), followed by flower disc diameter (23.87 cm), number of flowers (16.52) and flower stalk length (10.21). The mean values of cluster VII ranked first for stalk length (44.97 cm), stalk diameter (6.12 mm), flower diameter (10.89 cm) and vase life (5.95 days). The mean values of cluster VI recorded the highest yield in terms of number of flowers (29.73 per plant). Cluster IV ranked first for plant spread (49.58 cm). Cluster II recorded the highest mean values for number of leaves (83.99) and number of suckers (18.99). The mean values of clusters II and III showed an overall medium in the case of yield and quality contributing characters.
Table 3. Cluster mean value for different characters and their contribution (%) in diversity of 37 genotypes of gerbera
Genotypic and phenotypic correlation for growth and floral traits
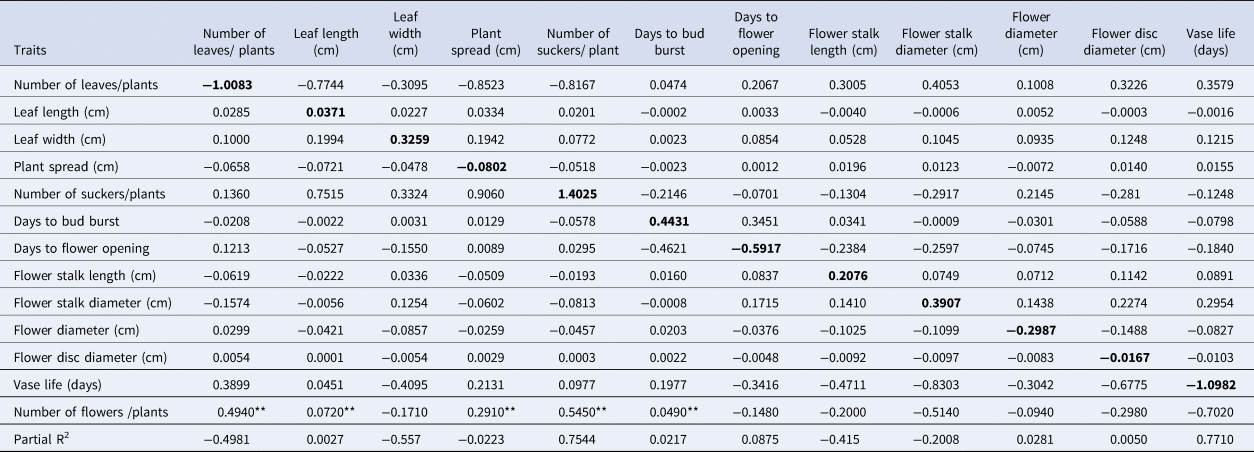
The results showed that the genotypic correlation coefficients for flower yield and quality contributing traits were higher than the phenotypic correlation coefficients (Table 4). Among the traits at genotypic and phenotypic levels, the number of leaves was positively correlated with leaf length, leaf width, plant spread and the number of suckers per plant. Similarly, flower yield showed a significant and positive correlation with the number of leaves (0.494**), plant spread (0.291**) and number of suckers (0.545**). While flower yield was negatively correlated with flower stalk length (−0.200**), stalk diameter (−0.514**) and disc diameter (−0.298**). Stalk length and stalk diameter had a significant positive genotypic correlation with days to flower opening, flower diameter and flower head disc diameter. Flower head diameter had a positive correlation with leaf dimension, plant spread, number of suckers, stalk length, stalk diameter, disc diameter and vase life. Vase life had a significant positive genotypic correlation with leaf width, flower stalk length, flower stalk diameter, flower head diameter and flower disc diameter but had a negative relationship with other vegetative traits.
Table 4. Genotypic (above diagonal) and phenotypic (below diagonal) correlation for growth and floral traits of 37 genotypes of gerbera
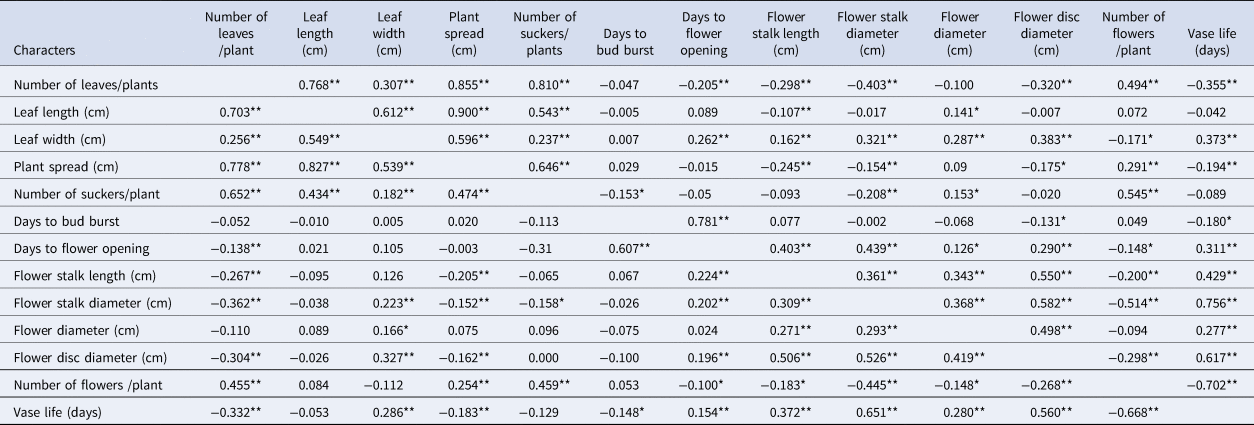
*, ** Significant at 5% and 1%, respectively.
Principal component analysis
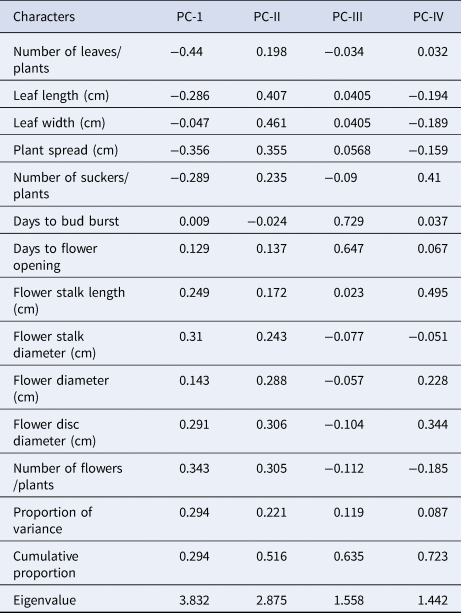
The results of the PCA analysis also revealed the presence of variability for different traits (Table 5). The first four components had an extracted eigenvalue of >1 (Figure S1), which contributed 72.3% of the total variation among the genotypes. Principal component 1 (PC1) contributed 29.4% of the total variability. PC2 contributed 22.1%, PC3 (11.9%) and PC4 (8.7%) to the total variability.
Table 5. Principal component analysis for growth and floral traits of 37 genotypes of gerbera
Path coefficient analysis
The path analysis at the genotypic level revealed that a number of suckers per plant (1.403), days to bud burst (0.443), flower stalk diameter (0.391) and flower stalk length (0.208) contribute major positive direct effects (Table 6). Similarly, other characters that had a positive direct effect are leaf dimension, which is reinforced by the positive and highly significant genotypic correlation of the number of suckers with flower yield.
Table 6. Path coefficient showing direct (bold) and indirect effect of observed characters on number of flowers
R Square = 0.8609 Residual effect = 0.3730.
Discussion
The phenotypic coefficients were slightly higher than the genotypic coefficient of variation for all the characters, with majority of the traits having high heritability (>60%) and genetic advance (>20%). The cluster analysis (D2) grouped the accessions into 7 clusters and showed wider genetic diversity within and between the clusters. The number of leaves, number of flowers, flower disc diameter and flower stalk length contributed the most to genetic divergence. Flower yield showed a significant positive genotypic correlation with the number of leaves, plant spread and suckers. The path analysis (genotypic) revealed that the number of suckers, leaf width, days to bud burst, flower stalk diameter and flower stalk length contribute a strong and positive direct effect on the number of flowers. The difference between PCV and GCV was very low for number of leaves, leaf length, plant spread, flower stalk length and diameter, number of flowers and vase life. In these characters, the expression of traits may not be solely due to genotypes but also reflect a little influence of the environment, as indicated by the ECV, which ranged between 0.69 and 3.31 in number of flowers and days to bud burst, respectively (Table 1). Hence, a lower sensitivity of genotypes to the environment and a greater role of genetic control govern the characters, as explained by Kumari et al. (Reference Kumari, Patel and Choudhary2011), Sravan et al. (Reference Sravan, Rangare, Suresh and Kumar2012) and Tiwari et al. (Reference Tiwari, Tripathi, Tripathi, Khatri and Bastola2019).
The assessment of the heritable portion of variation through heritability estimates and genetic gain showed that the number of flowers, number of suckers, stalk length, vase life, days to bud burst, flower disc diameter, leaf dimension, plant spread and number of leaves had high h2 (>60%) with high GA (>20%). The previous studies by Kumari et al. (Reference Kumari, Patel and Choudhary2011) and Ghimiray and Sarkar (Reference Ghimiray and Sarkar2015) also showed similar results for the number of flowers, number of suckers, stalk length and vase life in gerbera. Such traits are governed by additive gene action; thus, a selection based on phenotypic performance would be more effective in breeding programmes (Nair and Shiva, Reference Nair and Shiva2003; Kolte, Reference Kolte2008). The magnitude of heritable variability is the most significant feature of the genetic make-up of the selection programme in Gerbera (Kannan and Ramdas, Reference Kannan and Ramdas1990). The high h2 and low to moderate GA observed for flower stalk diameter and flower diameter indicate the effect of non-additive gene action, and the effect of environment on the expression of these characters was higher than the other traits. Therefore, selection may not be effective for this character (Panse and Sukhatme, Reference Panse and Sukhatme1967).
The magnitude of correlation is higher at the genotypic level for flower yield and quality-related traits as compared with the corresponding phenotypic level. It indicates that there is a strong inherent association between these traits. Among the traits at the genotypic and phenotypic levels, the number of leaves per plant showed a positive correlation with plant spread, number of suckers, number of flowers and flower diameter. The selection of genotypes on the basis of the number of leaves may be reliable for higher yields of both flowers and suckers. The higher leaf number and plant spread resulted in maximum accumulation of photosynthates and translocation with proper uptake of nutrients towards higher production of suckers and the number of flowers (Rymbai et al., Reference Rymbai, Jha and Talang2021). The number of leaves in plants reflects their good vigour and their suitability to the environment; hence, the exploitation of plant vigour in gerbera may be utilised for crop improvement. Our studies further showed that flower stalk diameter had a positive correlation with flower head diameter and vase life. This might mean that a thicker stalk could supply higher nutrient uptake to the flower head, as Khalaj et al. (Reference Khalaj, Kumar and Roosta2014) found that gerbera flowers with strong stems were less likely to fold in the vase. The stalk parameters are easy to observe in the field and are the most important traits related to shelf-life longevity and floral traits in gerbera. Hence, selection of genotypes that have a thicker stalk would be critical to increasing the vase life of flowers.
Hybrids of cluster III such as RCGH-12 (Deep Yellowish Pink, 46D), RCGH-22 (Vivid Reddish Orange, 32 A), RCGH-114 (Vivid Reddish Orange, 40 A) and RCGH-117 (Strong Orange, 24 A) had superior flower quality, particularly in flower head colour and yield (number of flowers). The monogenetic cluster V (RCGH 28) had the unique trait of dwarfing flower stalks in nature (28.33 cm), and the VII (Alesmera) had the maximum stalk length and diameter, flower diameter and vase life. The inter-cluster distance was higher than the intra-cluster distance, which suggested the existence of wide divergences among the parents. Hence, accessions in these clusters (III, V and VII) could be utilised as parents or genetic material sources in the hybridisation programme for getting desirable transgressive segregants for colour (ray floret and disc floret), flower stalk length and diameter, flower head diameter, number of flowers, number of suckers and vase life. PCA analysis indicates the existence of variability among the genotypes for different traits (Table 5). The first four components with a higher extracted Eigenvalue (>1) contributed 72.3% of the total variation. PCA revealed that the first principal component strongly correlated with the number of flowers, flower disc diameter, flower stalk length and flower diameter, indicating that these traits vary together in the same direction. Therefore, the first component can be varied as a measure of the flower quality and yield, and thus, it can be inferred that yielding traits of gerbera flowers are linked with stalk and flower dimension. Similarly, the second component showed the relationship between leaf characteristics and flower yield. The third and fourth components varied as a measure of plant vegetative traits and flower traits, respectively. The genotypes RCGH-12, RCGH-22, RCGH-114, RCGH-117 and Alesmera had very high values for the first component, and it is expected that this germplasm would have high values for the characters for which they are strongly correlated. Further, these genotypes (RCGH-12, RCGH-22, RCGH-114 and RCGH-117) are also grouped together in cluster III and have superior flower quality with attractive colours and a higher number of flowers. The monogenic cluster V (RCGH-28) had the unique trait of a dwarf flower stalk (28.33 cm), and cluster VII (Alesmera) was known for other biological characters such as a thick stalk, a large flower diameter and a long vase life. The inter-cluster distance was higher than the intra-distance distance, which suggested the existence of wide divergences among the genotypes of different groups (Titumeer et al., Reference Titumeer, Rahim and Zeba2014). Hence, such genotypes from such a distinct cluster could be utilised as parents or genetic material sources in the breeding programme to generate desirable transgressive segregants for colour (ray floret and disc floret), flower stalk length and diameter, flower head diameter, number of flowers, number of suckers and vase life. The number of flowers per plant had a strongly positive direct effect on the number of suckers per plant, days to bud burst, flower stalk diameter, leaf width and flower stalk length. Consequently, these characters may be considered the main components for selection in a breeding programme for higher flower yield and quality in Gerbera. Paradoxically, while the number of leaves per plant, vase life, days to flower opening and flower diameter had a strong and negative direct effect on the number of flowers. This negative direct effect was counter balanced by the positive indirect influences through suckers per plant, leaf width, days to bud burst, flower stalk diameter and flower stalk length. An overall analysis of the path coefficient suggested that selection should be made with a high number of suckers followed by days to bud burst, leaf width, flower stalk diameter and flower stalk length to increase the yield of Gerbera flowers. Similarly, it has been reported that leaf area, plant spread and number of leaves per plant had the highest indirect effect on the number of flowers per plant (Magar et al., Reference Magar, Warade, Nalge and Nimbalkar2010).
Conclusions
The above findings have shown the presence of high variation with higher GCV and PCV for quantitative traits. Economic traits such as number of flowers per plant, stalk diameter and vase life show high heritability and genetic advance; hence, they are highly responsive to selection. Cluster (D2) analysis also indicated the presence of a wide range of diversity within and between the accessions. PCA has also shown the presence of variability for different traits and their contributions to total variability. Flower yield was significantly correlated with the number of suckers, number of leaves and plant spread at genotypic and phenotypic levels, with positive effects. Moreover, superior lines (RCGH-12, RCGH-22, RCGH-114 and RCGH-117) with higher yield (>23 flowers per plant), appealing colour and vase life (>5 days) may be recommended for commercial production. While RCGH 28, a dwarfing flower stalk, can be promoted for pots and home decoration. These accessions with superior traits can be used for a future breeding programme as well as commercial production.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S1479262123000679.
Acknowledgements
The authors acknowledged the ICAR - All India Coordinated Research Project on Floriculture and Director, ICAR Research Complex for NEH Region, Umiam, Meghalaya for the support provided throughout the entire research work.
Competing interest
None.








