Introduction
Reward processing includes the attribution of ‘motivational salience’, incorporating both approach behaviour towards a rewarding outcome, and the avoidance of an aversive outcome (Berridge, Reference Berridge2012) which has been linked to mesolimbic dopaminergic neurons (Berridge, Reference Berridge2012; Ferguson, Ahrens, Longyear, & Aldridge, Reference Ferguson, Ahrens, Longyear and Aldridge2020). One of the prevailing theories of psychosis suggests that the attribution of motivational significance or salience to contextually irrelevant or neutral stimuli may be associated with the onset of psychotic symptoms (Chapman, Reference Chapman1966; Kapur, Reference Kapur2003; McGhiee & Chapman, Reference McGhiee and Chapman1961), consistent with the idea that this may be induced by elevated dopaminergic release within mesolimbic reward pathways causing aberrant stimulus-reinforcement (Miller, Reference Miller1976).
Traditionally, the aberrant salience hypothesis has focused on the striatum and midbrain, with patients with psychosis presenting with increased dopamine levels (Howes et al., Reference Howes, Kambeitz, Kim, Stahl, Slifstein, Abi-Dargham and Kapur2012) and attenuated striatal activation during the anticipation of reward, indicative of the abnormal processing of reward-based stimuli (Radua et al., Reference Radua, Schmidt, Borgwardt, Heinz, Schlagenhauf, McGuire and Fusar-Poli2015). Recent evidence has implicated a ‘salience’ network, functioning to select internal and externally generated signals for higher-order processing (Seeley, Reference Seeley2019; Seeley et al., Reference Seeley, Menon, Schatzberg, Keller, Glover, Kenna and Greicius2007; Uddin, Reference Uddin2015) and in particular, the insular cortex, a core component within the network for its role in psychosis (Wylie & Tregellas, Reference Wylie and Tregellas2010). The insula functions as a network hub that coordinates information across multiple cognitive domains and processes (Uddin, Nomi, Hébert-Seropian, Ghaziri, & Boucher, Reference Uddin, Nomi, Hébert-Seropian, Ghaziri and Boucher2017). A range of insular abnormalities have been reported in established psychotic disorders including altered activation (Moran et al., Reference Moran, Tagamets, Sampath, O'Donnell, Stein, Kochunov and Hong2013; Palaniyappan, Simmonite, White, Liddle, & Liddle, Reference Palaniyappan, Simmonite, White, Liddle and Liddle2013; Smieskova et al., Reference Smieskova, Roiser, Chaddock, Schmidt, Harrisberger, Bendfeldt and Borgwardt2014), volume (Goodkind et al., Reference Goodkind, Eickhoff, Oathes, Jiang, Chang, Jones-Hagata and Etkin2015; Sheffield et al., Reference Sheffield, Huang, Rogers, Blackford, Heckers and Woodward2021; Shepherd, Matheson, Laurens, Carr, & Green, Reference Shepherd, Matheson, Laurens, Carr and Green2012), and connectivity (Li et al., Reference Li, Wang, Xiong, Bo, He, Li and Wang2019; O'Neill, Mechelli, & Bhattacharyya, Reference O'Neill, Mechelli and Bhattacharyya2019; Sheffield, Rogers, Blackford, Heckers, & Woodward, Reference Sheffield, Rogers, Blackford, Heckers and Woodward2020; Tian, Zalesky, Bousman, Everall, & Pantelis, Reference Tian, Zalesky, Bousman, Everall and Pantelis2019). In the clinical high risk for psychosis state, compared to healthy controls, there have also been reports of altered insula activation (Wilson et al., Reference Wilson, Bossong, Appiah-Kusi, Petros, Brammer, Perez and Bhattacharyya2019), volume (Borgwardt et al., Reference Borgwardt, Riecher-Rössler, Dazzan, Chitnis, Aston, Drewe and McGuire2007; Chan, Di, McAlonan, & Gong, Reference Chan, Di, McAlonan and Gong2011; Ellison-Wright, Glahn, Laird, Thelen, & Bullmore, Reference Ellison-Wright, Glahn, Laird, Thelen and Bullmore2008; Lee et al., Reference Lee, Niznikiewicz, Asami, Otsuka, Salisbury, Shenton and McCarley2016; Takahashi et al., Reference Takahashi, Wood, Yung, Phillips, Soulsby, McGorry and Pantelis2009b), and functional connectivity with other salience network regions (Li et al., Reference Li, Wang, Xiong, Bo, He, Li and Wang2019; Wang et al., Reference Wang, Ji, Hong, Poh, Krishnan, Lee and Zhou2016; Wotruba et al., Reference Wotruba, Michels, Buechler, Metzler, Theodoridou, Gerstenberg and Heekeren2014). Moreover, correlation between symptom severity and degree of insular atrophy (Takahashi et al., Reference Takahashi, Wood, Soulsby, McGorry, Tanino, Suzuki and Pantelis2009a), or activation, indexed using fMRI (Smieskova et al., Reference Smieskova, Roiser, Chaddock, Schmidt, Harrisberger, Bendfeldt and Borgwardt2014; Thusius, Romanowicz, Mlynek, & Sola, Reference Thusius, Romanowicz, Mlynek and Sola2018; Walter et al., Reference Walter, Suenderhauf, Smieskova, Lenz, Harrisberger, Schmidt and Borgwardt2016; Wilson et al., Reference Wilson, Bossong, Appiah-Kusi, Petros, Brammer, Perez and Bhattacharyya2019) have been reported. These findings have consistently highlighted insular pathology across early stages of psychosis and suggest that insular dysfunction may hold a prominent role in the onset of psychotic symptoms.
There is growing interest in the antipsychotic potential of cannabidiol (CBD), a non-addictive substance present in the extract of Cannabis sativa, following evidence that it may oppose the psychotomimetic and neurophysiological effects of delta 9-tetrahydrocannabinol (THC) in healthy individuals (Bhattacharyya et al., Reference Bhattacharyya, Morrison, Fusar-Poli, Martin-Santos, Borgwardt, Winton-Brown and McGuire2010; Englund et al., Reference Englund, Morrison, Nottage, Hague, Kane, Bonaccorso and Kapur2013; Gunasekera, Davies, Martin-Santos, & Bhattacharyya, Reference Gunasekera, Davies, Martin-Santos and Bhattacharyya2021) complemented by preliminary evidence of antipsychotic efficacy in patients with psychosis in some (Leweke et al., Reference Leweke, Piomelli, Pahlisch, Muhl, Gerth, Hoyer and Koethe2012; McGuire et al., Reference McGuire, Robson, Cubala, Vasile, Morrison, Barron and Wright2018) but not all (Boggs et al., Reference Boggs, Surti, Gupta, Gupta, Niciu, Pittman and Ranganathan2018) clinical trials, as well as an excellent tolerability profile across different age groups (Chesney et al., Reference Chesney, Oliver, Green, Sovi, Wilson, Englund and McGuire2020; Velayudhan, McGoohan, & Bhattacharyya, Reference Velayudhan, McGoohan and Bhattacharyya2021).
Consistent with this, there is a growing body of work investigating mechanisms that may underlie the antipsychotic potential of CBD. We have recently shown that CBD may normalise impaired activation of the medial temporal lobe, midbrain, striatum, and insula in the clinical high-risk state for psychosis (Bhattacharyya et al., Reference Bhattacharyya, Wilson, Appiah-Kusi, O'Neill, Brammer, Perez and McGuire2018; Davies et al., Reference Davies, Wilson, Appiah-Kusi, Blest-Hopley, Brammer, Perez and Bhattacharyya2020; Wilson et al., Reference Wilson, Bossong, Appiah-Kusi, Petros, Brammer, Perez and Bhattacharyya2019) and established psychosis (O'Neill et al., Reference O'Neill, Annibale, Blest-Hopley, Wilson, Giampietro and Bhattacharyya2021a, Reference O'Neill, Wilson, Blest-Hopley, Annibale, Colizzi, Brammer and Bhattacharyya2021b). However, the precise neurocognitive mechanisms underlying the therapeutic potential of CBD remain unclear (Bonaccorso, Ricciardi, Zangani, Chiappini, & Schifano, Reference Bonaccorso, Ricciardi, Zangani, Chiappini and Schifano2019; Davies & Bhattacharyya, Reference Davies and Bhattacharyya2019; Velayudhan et al., Reference Velayudhan, Diepen, Marudkar, Hands, Suribhatla, Prettyman and Bhattacharyya2014), particularly when considering its effect on motivational salience (Gunasekera, Diederen, & Bhattacharyya, Reference Gunasekera, Diederen and Bhattacharyya2022). In a previous study in people at clinical high risk for psychosis, we have reported that CBD attenuated insular hyperactivation while processing motivationally salient stimuli during the anticipation phase of the monetary incentive delay task (MIDT) (Wilson et al., Reference Wilson, Bossong, Appiah-Kusi, Petros, Brammer, Perez and Bhattacharyya2019), a reward processing paradigm adapted for fMRI (Knutson, Westdorp, Kaiser, & Hommer, Reference Knutson, Westdorp, Kaiser and Hommer2000). This study investigated people at clinical high risk for psychosis, as opposed to those with established psychosis, and employed a between-group design. Hence, one cannot completely rule out the possibility that between-group (placebo-treated and CBD-treated patient groups) differences observed in the study were a result of between-subject variability rather than truly an effect of treatment with CBD. These limitations prevented direct examination of the effects of CBD, compared to placebo, with brain activation and psychopathology change in the same participants.
Therefore, we investigated the effects of CBD on brain activation, indexed using the blood-oxygen-level-dependent (BOLD) haemodynamic response measured using functional resonance imaging (fMRI) during motivational salience processing in patients with early psychosis (PSY). Here, we defined early psychosis as a psychotic mental disorder diagnosis within 5 years of presentation to clinical services (Malla et al., Reference Malla, Joober, Iyer, Norman, Schmitz, Brown and Abadi2017). Using a within-subject design, we investigated whether acute CBD administration would attenuate abnormal brain activation within PSY, compared to placebo. We also compared PSY patients under placebo to a group of healthy controls studied under identical conditions but not receiving any treatment. This was conducted to help understand whether the acute effects of CBD, compared to placebo, on brain activation in psychosis patients overlapped with brain regions that were differentially activated in placebo-treated psychosis patients compared to healthy controls (i.e. regions affected by psychosis) and to examine whether the direction of any CBD-related change in activation in patients was consistent with a likely therapeutic benefit. We used the MIDT paradigm which comprises of ‘anticipation’ and ‘feedback’ conditions. Here we have focused on the anticipation condition, as in our previous study (Wilson et al., Reference Wilson, Bossong, Appiah-Kusi, Petros, Brammer, Perez and Bhattacharyya2019). Meta-analytic findings have identified robust insular engagement during the anticipation of both reward and loss (Wilson et al., Reference Wilson, Colizzi, Bossong, Allen, Kempton, Abe and Bhattacharyya2018). Therefore, consistent with our previous study (Wilson et al., Reference Wilson, Bossong, Appiah-Kusi, Petros, Brammer, Perez and Bhattacharyya2019) and other work (Nielsen et al., Reference Nielsen, Rostrup, Wulff, Bak, Broberg, Lublin and Glenthoj2012a, Reference Nielsen, Rostrup, Wulff, Bak, Lublin, Kapur and Glenthøj2012b), we examined all motivationally salient (reward and loss avoidance) conditions of the MIDT. Based on results in our previous study in patients at clinical high-risk psychosis, we predicted that PSY participants would express greater levels of activation in the insula, compared with healthy controls, and that a single dose of CBD would attenuate this abnormal brain activity. Exploratory analyses examined effects at the whole-brain level and within a hippocampus-midbrain-striatum mask.
Methods
The methods are reported in full in the online Supplementary Methods and have been summarised below in the interest of brevity.
The study protocol was approved by the appropriate research ethics committee (reference: 14/LO/1861). All participants took part after providing written informed consent. Participants with early psychosis were recruited from mental health services in South London, United Kingdom. Fifteen participants attended for 2 study days, and 14 completed two fMRI scanning sessions [see (O'Neill et al., Reference O'Neill, Wilson, Blest-Hopley, Annibale, Colizzi, Brammer and Bhattacharyya2021b)]. Patients were included if they had a psychotic mental disorder diagnosis (meeting criteria for schizophrenia, schizophreniform, or brief psychotic disorder, but no other Axis I diagnoses) within 5 years of illness onset and did not have a diagnosis of alcohol or a substance use disorder (excluding cannabis). Nineteen healthy control (HC) participants, who were not administered CBD or placebo were also included. Additional inclusion/ exclusion criteria as well as advice regarding caffeine, alcohol and other drugs use and urine drug screening and smokerlyzer tests on study days for both participant groups are reported in online Supplementary Methods.
We employed a within-subject, crossover, double-blind randomised placebo-controlled design, in patients, over 2 sessions with a 1-week interval to allow for the washout of CBD. Psychosis participants were administered either a 600 mg CBD (approx. 99.9% pure) or an identical gelatine placebo (PLB) capsule. One hundred and eighty minutes after drug administration participants underwent fMRI scanning and performed the MIDT. Blood samples were obtained at three-time points: (T1) 60 min before drug administration, (T2) 60 min post drug administration, (T3) 270 min post-drug administration. The Positive and Negative Syndrome Scale (Kay, Fiszbein, & Opler, Reference Kay, Fiszbein and Opler1987) was used to assess psychopathology at timepoints T1 and T3.
The data presented in this study is part of a larger study which utilised a number of neuroimaging approaches (O'Neill et al., Reference O'Neill, Annibale, Blest-Hopley, Wilson, Giampietro and Bhattacharyya2021a, Reference O'Neill, Wilson, Blest-Hopley, Annibale, Colizzi, Brammer and Bhattacharyya2021b). For the overall study, an initial power calculation was conducted (please see online Supplementary Methods), however, this focused on brain activation during a verbal learning task (O'Neill et al., Reference O'Neill, Wilson, Blest-Hopley, Annibale, Colizzi, Brammer and Bhattacharyya2021b) rather than specifically for the MIDT that we report here.
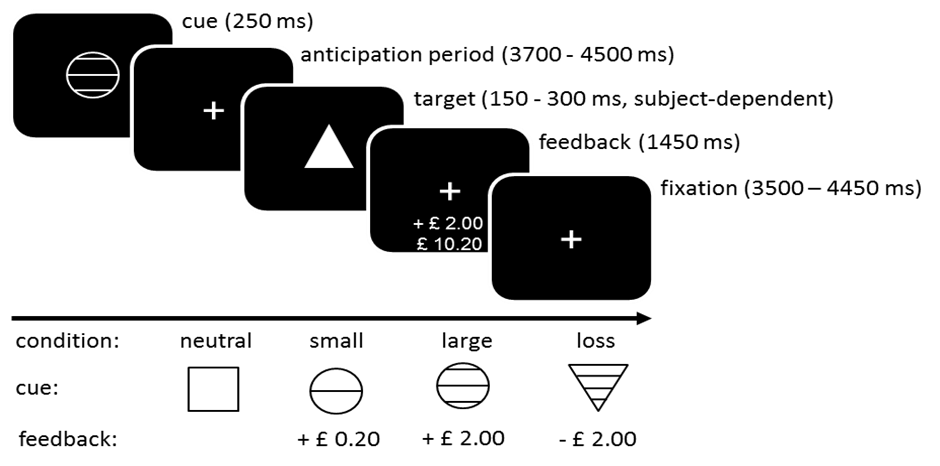
Monetary incentive delay task
Participants completed two runs of the MIDT (see online Supplementary Fig. S1 for task schematic) each consisting of 48 individual trials (approximately 16 min in two consecutive 8-min runs). The MIDT was comprised of four reward valence conditions signalled by learned visual cues: neutral (£0.00), small reward (£0.20), large reward (£2.00), and loss (£2.00). A total of 12 trials were used for each. All participants started with £10.00 and were provided with payment based on the actual monetary reward they earned over the two runs of the task on each study day. Participants underwent standardised training prior to entering the scanner.
Data analysis
Imaging analysis
Functional MRI data were pre-processed and analysed using SPM8. Data pre-processing steps consisted of functional image realignment, anatomical scan co-registration, spatial normalisation into a standard MNI space via unified segmentation, and smoothing via Gaussian filter (FWHM = 8 mm). Using general linear model regression with factors time-locked to task events convolved with a canonical hemodynamic response function, the regression coefficient (beta weight) for each condition at each voxel was determined. Twelve regressors were included in the task design: 4 anticipatory conditions (win small, win large, loss, neutral), 7 feedback conditions (feedback was displayed for neutral, and for the hit or miss of win small, win large and loss conditions), and 1 regressor that modelled all 4 anticipation conditions collapsed together. As we were interested in the effect of group on salience processing, we examined this across all reward valence conditions (win small, win large, and loss, each contrasted with neutral) during anticipation.
Group comparisons of HC-v.-PSY-PLB and PSY-PLB-v.-PSY-CBD were analysed using a flexible factorial model in SPM8. The between-group model was created by specifying the factors ‘Group’ and ‘Reward Valence Conditions’. The within-group model was created in the same way with the additional ‘Subject’ factor specified in the design matrix. Using F contrasts (1 1 1 −1 −1 −1) the group by reward anticipation effect was determined. Contrast weights were extracted to determine contrast directions.
Brain activation was examined using region of interest (ROI) analysis. Two ROI masks were created using Pick Atlas in SPM8: a primary bilateral insula mask and a secondary mask comprising the hippocampus, midbrain and striatum (composed of bilateral medial hippocampi, subicula, caudate, putamen, pallidum and midbrain). Effects at the whole-brain level were also investigated. All neuroimaging results are reported using a family-wise error threshold.
Linear regression analysis was then conducted to determine if a relationship between mean contrast estimates during motivational salience processing (across anticipation win small, win large, and avoid lose conditions; extracted from the insula anatomical ROI) and PANSS total score change on study day (T1 minus T3) was significantly different under CBD compared with the placebo condition.
Performance analysis
Two group comparisons (HC-v.-PSY-PLB and PSY-PLB-v.-PSY-CBD) were carried out using Python 3.8.8 (Python Software Foundation, 2016) (code available at https://github.com/bgunasekera/Hemp/blob/main/megafile_analysis.py). Pairwise comparisons were applied on mean monetary reward (£), accuracy (correct responses %), reaction time (ms) and false-starts [a response < 100 ms was considered a false start (Brosnan, Hayes, & Harrison, Reference Brosnan, Hayes and Harrison2017)]. t tests were conducted for mean monetary reward, analysis of variance (ANOVA) for mean reaction time, and binary logistic regression for accuracy, and false-starts. Repeated measures were specified for PSY-PLB-v.-PSY-CBD during t test and anova comparisons.
Results
Socio-demographic and substance use history of the participant groups have been reported before (O'Neill et al., Reference O'Neill, Wilson, Blest-Hopley, Annibale, Colizzi, Brammer and Bhattacharyya2021b) and are summarised again in Table 1. Although, one patient tested positive on urine drug screen for phencyclidine (PCP) on both study days, this was disregarded as the person was receiving venlafaxine, which has been reported to induce false PCP positive results (Landy & Kripalani, Reference Landy and Kripalani2015). Antipsychotic medication was being used by all participants, except for one who had discontinued use of their prescribed olanzapine medication.
Table 1. Sociodemographic and clinical measures (previously also reported in O'Neill et al. (Reference O'Neill, Annibale, Blest-Hopley, Wilson, Giampietro and Bhattacharyya2021a, Reference O'Neill, Wilson, Blest-Hopley, Annibale, Colizzi, Brammer and Bhattacharyya2021b) *patient was prescribed olanzapine but was not taking it)

All HC individuals had a lifetime cannabis use of less than 10 times.
Clinical characteristics, and the change in symptoms as a result of treatment of the participants taking part in this study have been reported before (O'Neill et al., Reference O'Neill, Annibale, Blest-Hopley, Wilson, Giampietro and Bhattacharyya2021a) and are summarised here again (Table 2). On each study day, prior to drug administration, there was no significant difference in total PANSS between PSY-CBD and PSY-PLB groups (z = −1.07, p = 0.14). Following drug administration, compared to baseline, patients reported an improvement in total PANSS score which was significantly greater under CBD than under PLB (z = −2.14; p = 0.02).
Table 2. Symptom scores for HC and PSY patients at baseline, and post-drug for both study days [previously reported in (O'Neill et al., Reference O'Neill, Annibale, Blest-Hopley, Wilson, Giampietro and Bhattacharyya2021a)]

PSY-PLB, psychosis participants with placebo; PSY-CBD, psychosis participants with cannabidiol; PANSS, Positive and Negative syndrome scale; STAI-S, State-Trait Anxiety Inventory state subscale. T1 = 60 min prior to drug administration, T3 = 270 min post-drug administration. Non-parametric Sign test was used to compare paired medians.
Behavioural performance
Mean monetary reward
At the end of the MIDT, the HC group won a significantly higher amount of monetary reward relative to PSY-PLB (Table 3; p = 0.03). No significant differences were observed when comparing PSY-PLB with PSY-CBD.
Table 3. Behavioural performance

HC, healthy controls; PSY-CBD, early psychosis cannabidiol group; PSY-PLB, early psychosis placebo group.
a Independent t test.
b Paired t test.
c Binary logistic regression.
d Analysis of variance.
e Repeated measures analysis of variance.
In the calculation of accuracy, responses <100 ms after target presentation were considered as an inaccurate response, and subsequently excluded from reaction time analysis, in accordance with previous work (Wilson et al., Reference Wilson, Bossong, Appiah-Kusi, Petros, Brammer, Perez and Bhattacharyya2019).
Accuracy
Across both sets of comparisons (HC v. PSY-PLB and PSY-PLB v. PSY-CBD), there was a main effect of stimulus type (salient v. neutral) on task accuracy, such that accuracy percentage was higher for salient compared to neutral stimuli (p < 0.001 and p = 0.007 respectively). In separate pairwise comparisons, there was also a main effect of group (HC v. PSY and PSY-CBD v. PSY-PLB) on task accuracy (Table 3), such that the PSY-PLB group were less accurate than HC (p < 0.001) and PSY-CBD (p = 0.005) across all types of stimuli. No significant interactions between group and salience were observed in either of the comparisons (HC v. PSY or PSY-CBD v. PSY-PLB) (Table 3).
Reaction time
Across both sets of comparisons (HC v. PSY-PLB and PSY-PLB v. PSY-CBD), there was a main effect of stimulus type (salient v. neutral) on RT such that RT was lower for salient compared to neutral stimuli (p < 0.001 and p = 0.03). In separate pairwise comparisons, there was also a main effect of group in HC v. PSY on RT, such that the PSY-PLB group were slower than HC (p < 0.001) across all types of stimuli but this was not present in PSY-PLB v. PSY-CBD. No significant interactions between group and salience were observed in either of the comparisons (HC v. PSY or PSY-CBD v. PSY-PLB) (Table 3).
False-starts (response⩽100 ms)
Across both sets of comparisons (HC v. PSY-PLB and PSY-PLB v. PSY-CBD), there was no main effect of stimulus type (salient v. neutral) on the percentage difference of false starts. In separate pairwise comparisons, there was a main effect of group (HC v. PSY) such that the PSY-PLB group had a higher percentage of false starts (p < 0.001) across all types of stimuli. This relationship was not seen in the PSY-PLB v. PSY-CBD pairwise comparison. No significant interactions between group and salience were observed in either of the comparisons (HC v. PSY or PSY-CBD v. PSY-PLB) (Table 3).
Imaging
Hc-v.- PSY-PLB
No differences were observed when comparing PSY-PLB with HC during the salience v. neutral contrast (PSY-PLB > HC) within the bilateral insula or the bilateral hippocampus-midbrain-striatum ROI masks or at the whole-brain level.
Within the PSY group, nine participants were current cannabis users. Therefore, sensitivity analyses were conducted to control for the confounding effect of previous cannabis exposure by including this as a covariate within the analysis. The result remained the same after controlling for current cannabis use.
PSY-PLB-v.- PSY-CBD
Within the bilateral insula ROI mask (Fig. 1a and b), activation was significantly attenuated in the left insula following CBD compared with placebo during the salience v. neutral contrast (PSY-PLB > PSY-CBD) (p-FWE = 0.047, x = −36, y = 12, x = 12, k = 33).

Fig. 1. (a) PSY-PLB > PSY-CBD comparison, (b) Box plot of extracted contrast weight estimates showing attenuated activation in CBD group compared with the placebo group. p < 0.05 FWE-corrected, L = left, A = anterior.
The result remained unchanged after controlling for current cannabis use (p-FWE = 0.045; x = −36, y = 12, x = 12; k = 34).
There was no significant difference in the relationship between the contrast estimate extracted from the bilateral insula anatomical ROI and change (T1 minus T3) in PANSS total score on study day between the CBD and placebo treatment conditions (estimate = 5.87, s.e. = 11.20, p = 0.61, CI −15.2 to 27.0).
No areas of significant activation were observed within the hippocampus-midbrain-striatum ROI or at the whole-brain level.
Discussion
In this study we compared brain activation differences between healthy controls and those with early psychosis while processing motivationally salient stimuli and examined the acute effects of CBD, relative to placebo, in psychosis, focusing primarily on changes within the insula cortex, based on previous work. Contrary to our first prediction, we failed to identify differences in brain activation while processing motivationally salient stimuli in early psychosis compared to healthy controls. In accordance with our second hypothesis, we found an attenuation of insula activity in participants with early psychosis following a single dose of CBD, compared with placebo. In the psychosis group, these effects were accompanied by a lower mean monetary reward won during the task, as well as poorer performance accuracy, faster reaction time and premature action initiation across all salience conditions compared to healthy controls. An attenuating effect of CBD on insular activation in psychosis patients was accompanied by a concomitant improvement in performance accuracy across all salience conditions compared with placebo. Exploratory whole-brain analyses, as well as those focusing on the striatum-hippocampus-midbrain ROI, did not reveal any significant difference in pairwise comparisons across the combined salience conditions.
The neuroimaging results comparing healthy controls with early psychosis are inconsistent with our previous report that investigated the neural substrates involved in the antipsychotic effect of CBD in the context of reward processing in people at clinical high risk for psychosis (Wilson et al., Reference Wilson, Bossong, Appiah-Kusi, Petros, Brammer, Perez and Bhattacharyya2019). Using the MIDT, Wilson et al. (Reference Wilson, Bossong, Appiah-Kusi, Petros, Brammer, Perez and Bhattacharyya2019) identified increased activation in the left insula/parietal operculum in clinical high risk for psychosis participants under placebo compared to healthy controls not receiving any study drug. One explanation for not seeing a similar effect in the present study may be due to the confounding effects of dopamine antagonism in our early psychosis cohort who were taking antipsychotic medication at the time of study, unlike in the report by Wilson and colleagues where participants were antipsychotic-naïve. Another study investigating early psychosis participants reported a negative correlation between left insular activation during salience processing and cumulative antipsychotic medication dose (Walter et al., Reference Walter, Suenderhauf, Smieskova, Lenz, Harrisberger, Schmidt and Borgwardt2016). A further study, also using a salience processing task, identified reduced insula–anterior cingulate connectivity in early psychosis, relative to healthy controls, observed only in untreated patients and not in antipsychotic treated patients (Schmidt et al., Reference Schmidt, Palaniyappan, Smieskova, Simon, Riecher-Rössler, Lang and Borgwardt2016). These reports complement meta-analytic findings that antipsychotic medicated early psychosis patients are more likely to show structural abnormalities of grey matter volume in the left insula (Radua et al., Reference Radua, Borgwardt, Crescini, Mataix-Cols, Meyer-Lindenberg, McGuire and Fusar-Poli2012). Therefore, antipsychotic treatment may partly ameliorate insular activation abnormalities in patients with psychosis, such that they may no longer be detectable in comparison with healthy controls. Nevertheless, by employing a within-subject design in the present study, as opposed to a between-group design by Wilson et al. (Reference Wilson, Bossong, Appiah-Kusi, Petros, Brammer, Perez and Bhattacharyya2019), we were able to extend those results to clearly demonstrate that a single dose of CBD can attenuate salience processing-related insular activation in patients with established psychosis. In accordance with previous results (Wilson et al., Reference Wilson, Bossong, Appiah-Kusi, Petros, Brammer, Perez and Bhattacharyya2019), we also found a shorter RT to salient compared to neutral stimuli across all participant groups, consistent with the idea that faster responding during MIDT may indicate greater salience perception (Mir et al., Reference Mir, Trender-Gerhard, Edwards, Schneider, Bhatia and Jahanshahi2011).
Evidence suggests that atypical insula engagement, within the salience network, is a feature of psychotic disorders (Palaniyappan & Liddle, Reference Palaniyappan and Liddle2012; Walter et al., Reference Walter, Suenderhauf, Smieskova, Lenz, Harrisberger, Schmidt and Borgwardt2016). Upon detection of a salient stimulus, the insula facilitates task-related information processing by initiating appropriate transient control signals to brain areas mediating attentional, working memory, and higher-order cognitive processes, while disengaging the default mode network to facilitate goal-directed behaviour (Uddin, Reference Uddin2015). Significantly, previous studies have associated aberrant left insular activation with psychosis (Raij, Mäntylä, Mantere, Kieseppä, & Suvisaari, Reference Raij, Mäntylä, Mantere, Kieseppä and Suvisaari2016; Thusius et al., Reference Thusius, Romanowicz, Mlynek and Sola2018; Walter et al., Reference Walter, Suenderhauf, Smieskova, Lenz, Harrisberger, Schmidt and Borgwardt2016; Wilson et al., Reference Wilson, Bossong, Appiah-Kusi, Petros, Brammer, Perez and Bhattacharyya2019).
Finally, we found no activation within the striatum, hippocampus, or midbrain in our exploratory analyses. This is in line with our previous report in people at clinical high risk of psychosis (Wilson et al., Reference Wilson, Bossong, Appiah-Kusi, Petros, Brammer, Perez and Bhattacharyya2019). In other reports from the same clinical high-risk cohort as that in Wilson and colleagues, we have identified altered striatal and parahippocampal gyrus activation in psychosis compared to healthy participants and their modulation by CBD following verbal learning and fear paradigms (Bhattacharyya et al., Reference Bhattacharyya, Wilson, Appiah-Kusi, O'Neill, Brammer, Perez and McGuire2018; Davies et al., Reference Davies, Wilson, Appiah-Kusi, Blest-Hopley, Brammer, Perez and Bhattacharyya2020). Therefore, as we have indicated before (Wilson et al., Reference Wilson, Bossong, Appiah-Kusi, Petros, Brammer, Perez and Bhattacharyya2019), these results are likely reflective of the different cognitive paradigms employed in these studies, e.g. MIDT in the present study and in Wilson et al. (Reference Wilson, Bossong, Appiah-Kusi, Petros, Brammer, Perez and Bhattacharyya2019), verbal memory in Bhattacharyya et al. (Reference Bhattacharyya, Wilson, Appiah-Kusi, O'Neill, Brammer, Perez and McGuire2018) and fear processing in Davies et al. (Reference Davies, Wilson, Appiah-Kusi, Blest-Hopley, Brammer, Perez and Bhattacharyya2020). In particular, these differences may arise from the distinct roles of the insula and striatum in the processing of salient stimuli. While the striatum may be involved in the attribution of motivational salience to stimuli (Kapur, Reference Kapur2003), the insula has been suggested to be involved in proximal salience which occurs during the evaluation of stimuli (Palaniyappan & Liddle, Reference Palaniyappan and Liddle2012). The concept of proximal salience should be considered as an extension of aberrant salience to include the disruption of cognition and volition in psychosis (Palaniyappan & Liddle, Reference Palaniyappan and Liddle2012). It has been suggested that insular dysconnectivity induces the inappropriate assignment of proximal salience. Specifically, this is when external stimuli generate activity within the salience network that updates expectations which then lead to initiation or modification of action. This, in turn, is thought to contribute to the onset of perceptual and cognitive distortions, disorganisation, and psychomotor slowing (Palaniyappan & Liddle, Reference Palaniyappan and Liddle2012).
While the precise molecular mechanisms that may underlie the effects of CBD reported here remain unclear, a number of potential candidate mechanisms have been suggested, such as negative allosteric modulation of CB1 receptors (Laprairie, Bagher, Kelly, & Denovan-Wright, Reference Laprairie, Bagher, Kelly and Denovan-Wright2015), weak antagonism of CB2 receptors (Thomas et al., Reference Thomas, Baillie, Phillips, Razdan, Ross and Pertwee2007), partial agonism of D2 receptors (a mechanism also shown by aripiprazole) (Tuplin & Holahan, Reference Tuplin and Holahan2017), inhibition of anandamide hydrolysis (Bisogno et al., Reference Bisogno, Hanuš, De Petrocellis, Tchilibon, Ponde, Brandi and Di Marzo2001), and stimulation of vanilloid receptor type 1 (Bisogno et al., Reference Bisogno, Hanuš, De Petrocellis, Tchilibon, Ponde, Brandi and Di Marzo2001) and 5-HT1A receptors (Sartim, Guimarães, & Joca, Reference Sartim, Guimarães and Joca2016). In any case, direct or indirect modulation of endogenous anandamide signalling by CBD may be a potential mechanism of antipsychotic action that is consistent with independent evidence of altered CB1 receptor levels across a number of brain regions including in the insula of schizophrenia patients (Ceccarini et al., Reference Ceccarini, De Hert, Van Winkel, Peuskens, Bormans, Kranaster and Van Laere2013; Ranganathan et al., Reference Ranganathan, Cortes-Briones, Radhakrishnan, Thurnauer, Planeta, Skosnik and D'Souza2016).
Limitations
The results presented must be considered in light of the modest sample size highlighted by the power calculation reported. The data presented in this study is a subset of a larger study which utilised a number of neuroimaging approaches where an initial power calculation was conducted, however, this focused on medial temporal activation during a verbal learning task (O'Neill et al., Reference O'Neill, Wilson, Blest-Hopley, Annibale, Colizzi, Brammer and Bhattacharyya2021b) rather than estimating power specifically for the MIDT that we employed here. Therefore, although the sample size utilised in this report is within the range of suitable power, the absence of a power calculation specific for the MIDT should be considered as a limitation. While differences in task performance (accuracy and reaction time) were detected, brain activation differences between psychosis patients under placebo and healthy controls were not detected. Although group differences in antipsychotic exposure and cannabis use may also explain our inability to detect differences in this comparison, we cannot rule out that limited power affected the results reported here. This underscores the need for future studies to use larger samples. Future studies may also consider investigation of the acute effects of CBD relative to placebo in healthy controls in parallel with the design employed in patients, to aid interpretation of the specificity of the effects of CBD in the psychosis patients.
Another limitation of this study was the confounding effects of dopamine antagonism that likely precluded our ability to detect a difference in insular activation in psychosis patients, relative to healthy control participants. Moreover, the healthy participants in this study had a lifetime cannabis use history of fewer than 10 instances. This is in contrast to the participants within the psychosis group, who were all lifetime cannabis users with 9 of them being current regular users. Although results remained unchanged following sensitivity analysis that attempted to control for the potential confounding effect of difference in cannabis exposure between the two groups, we cannot be certain that this did not affect our ability to detect significant differences in brain activation between the healthy and psychosis group. Nevertheless, we were able to detect an attenuating effect of CBD on insular activation that was over and above any potential effect of antipsychotic medications on insular activation using a within-subject design. While the use of a healthy control group as a comparator with placebo-treated psychosis patients was intended to help examine whether the CBD effects in psychosis patients were observed in the same regions that were differentially affected in psychosis patients under placebo, compared to healthy controls, and to contextualise the direction of any CBD-related change in activation in patients, group differences in cannabis and antipsychotic exposure may have precluded our ability to detect disease-related change as originally intended. Therefore, the results presented here indicate that future studies may also need to consider strategies to better match participant groups in terms of potential confounding factors such as cannabis exposure and antipsychotic exposure.
It is also important to consider that the MIDT recruits cognitive processes other than motivational salience, as evident from meta-analyses examining the anticipation phase of the MIDT in healthy participants. These studies have highlighted the engagement of a number of brain regions outside of the salience network that may serve a range of processes including executive function, psychomotor control and impulsivity (Jauhar et al., Reference Jauhar, Fortea, Solanes, Albajes-Eizagirre, McKenna and Radua2021; Oldham et al., Reference Oldham, Murawski, Fornito, Youssef, Yücel and Lorenzetti2018; Wilson et al., Reference Wilson, Colizzi, Bossong, Allen, Kempton, Abe and Bhattacharyya2018). Further, the insula is known to be engaged in a range of cognitive processes other than salience processing (Uddin et al., Reference Uddin, Nomi, Hébert-Seropian, Ghaziri and Boucher2017). Therefore, we cannot be completely certain that engagement of some of these processes may have influenced the results presented here, although, lack of group differences in brain activation outside of the insula indicate that this may be less likely.
Finally, although the primary aim of this study was to complement clinical investigations of CBD in people with established psychosis by investigating potential mechanisms which may underlie its putative antipsychotic action, it remains to be seen whether similar effects on brain substrates, as those reported here, would be observed following sustained dosing. In particular, it would be important to demonstrate longitudinal changes in brain activation following sustained CBD dosing that occurs in parallel with the improvement in the severity of psychotic symptoms.
Conclusion
This study highlighted an attenuating effect of CBD on insular activation, in participants with an established psychotic disorder during salience processing that is consistent with independent evidence. Whether these effects of CBD on insular activation persist following sustained treatment and whether they are related to any symptomatic benefits remains to be seen in larger studies.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0033291722001672.
Acknowledgements
This work was supported by grants from the Medical Research Council (MRC), UK (MR/J012149/1 and MC_PC_14105 v.2 to SB). SB. has also received support from the National Institute for Health Research (NIHR) (NIHR Clinician Scientist Award; NIHR CS-11-001), the NIHR Mental Health Biomedical Research Centre at South London and Maudsley National Health Service (NHS) Foundation Trust and King's College London, and Parkinson's UK.
Author contributions
The views expressed are those of the author(s), and not necessarily those of the NHS, the NIHR, or the Department of Health. All the authors contributed in a substantial way to the study and approved the manuscript content. The authors have no other biomedical financial interests or potential conflicts of interest related to this publication.
Conflict of interest
The authors report no conflicts of interest.