


INTRODUCTION
The marine reservoir effect is a well-known phenomenon in radiocarbon dating. The 14C age of dissolved inorganic carbon (DIC) in seawater is on average 400 yr older than that of the coeval atmosphere, and marine carbon is 400 14C yr older than terrestrial carbon (Stuiver et al. Reference Stuiver, Pearson and Braziunas1986, Reference Stuiver, Pearson and Braziunas1998; Stuiver and Braziunas Reference Stuiver and Braziunas1993; Hughen et al. Reference Hughen, Baillie, Bard, Beck, Bertrand, Blackwell, Buck, Burr, Cutler, Damon, Edwards, Fairbanks, Friedrich, Guilderson, Kromer, McCormac, Manning, Bronk Ramsey, Reimer, Reimer, Remmele, Southon, Stuiver, Talamo, Taylor, van der Plicht and Weyhenmeyer2004). This effect occurs because the surface ocean (surface mixed layer) exchanges DIC with carbon in the deep ocean, where DIC residence times are sufficiently long that 14C decay is significant. Therefore, 14C ages of materials based on the marine food chain are affected by the marine reservoir effect (Fujio et al. Reference Fujio, Imamura and Nishimoto2005; Horiuchi et al. Reference Horiuchi, Miyata, Kamijo, Cramp and Evershed2015). The marine reservoir effect is especially large in the North Pacific because 14C-depleted seawater that originates from North Atlantic Deep Water upwells there (Stommel Reference Stommel1958; Broecker et al. Reference Broecker, Peng, Ostlund and Stuiver1985; Broecker Reference Broecker1991). Such local variations of the marine reservoir effect can be corrected by using a regional correction value (∆R), which is estimated by subtracting model-based marine reservoir ages from the observed total marine reservoir ages of marine shells or other marine materials of known age (Stuiver et al. Reference Stuiver, Pearson and Braziunas1986; Stuiver and Braziunas Reference Stuiver and Braziunas1993; Southon et al. Reference Southon, Rodman and True1995, Reference Southon, Kashgarian, Fontugne, Metivier and Yim2002; Reimer et al. Reference Reimer, McCormac, Moore, McCormick and Murray2002). The model marine ages used in ∆R calculation are from the international calibration curves (currently Marine13, Reimer et al. Reference Reimer, Bard, Bayliss, Beck, Blackwell, Bronk Ramsey, Grootes, Guilderson, Haflidason, Hajdas, Hatté, Heaton, Hoffmann, Hogg, Hughen, Kaiser, Kromer, Manning, Niu, Reimer, Richards, Scott, Southon, Staff, Turney and van der Plicht2013). The localization of ΔR and its changes with elapsed time are estimated by using the apparent 14C ages of pre-bomb molluskan shells or other marine materials of known calendar age.
Charred materials on potsherds from northern Tohoku to Hokkaido, Japan, often give older ages than the coeval atmosphere (Fujio et al. Reference Fujio, Imamura and Nishimoto2005; Horiuchi et al. Reference Horiuchi, Miyata, Kamijo, Cramp and Evershed2015). These materials are likely affected by the marine reservoir effect because marine products may have been cooked in the pottery vessels. Since the marine reservoir effect is a function of time and location and because the charred materials have not necessarily originated from marine products only, it is difficult to estimate the time when the pottery was used from observed 14C ages of the charred material. In addition, if we measure the 14C ages of human bones from people who consumed marine products, their 14C ages will be older than the assumed actual age because of the marine reservoir effect, and it is very hard to correct these observed ages to obtain the actual age.
Thus, the 14C ages of archaeological remains affected by the marine reservoir effect have not always been used effectively. However, the marine reservoir effect of archaeological remains is very likely to reflect the diet (paleodiet) or habitat of the organisms whose remains are studied, and knowledge of their diet and habitat might allow the reconstruction of the marine environment when the archaeological site was inhabited (i.e. the paleoenvironment). Although the ages of charred materials from potsherds may be as much as a few hundred years older than the reservoir age of the ocean current flowing near the site, it is not easy to interpret these ages without additional information. Instead, they are likely to be dismissed as outliers.
With a sufficiently detailed archaeological examination, we can use apparent marine reservoir age differences to infer details of paleodiets and habitats and, hence, paleomarine environments. First, we need to date different kinds of archaeological remains preserved in the same layer, and we must be able to exclude remains affected by mixing with materials from the layers above and below. Then, we may be able to estimate the paleodiets of the animals whose remains are dated, such as mollusks, fish, and sea mammals, and reconstruct the paleomarine environment by referring to the habitats of those animals and the ocean currents near the site at which the remains are found. To attempt such a reconstruction, we chose the Hamanaka 2 site, where Miyata et al. (Reference Miyata, Horiuchi and Nishimoto2009) have already studied the paleodiet indicated by charred materials on pottery, and where different kinds of archaeological remains occur in the same layer and have a clear archaeological context (Nishimoto Reference Nishimoto2000).
HAMANAKA 2 ARCHAEOLOGICAL SITE
The excavation surveys at Hamanaka 2 were conducted by the National Museum of Japanese History (NMJH) from 1994 to 1997. Their purpose was to shed light on the northern cultural influence in Hokkaido, Japan, because Rebun Island is between Hokkaido and Sakhalin (Figure 1). The excavation findings, along with earlier research, showed that the site was occupied intermittently for about 2500 yr, from the latter half of the Jomon period to the Ainu period (1300 BC to AD 1200). Moreover, it was clearly indicated that not only the people from the north but also from the south repeatedly visited Rebun Island, where an active cultural exchange occurred (Maeda and Yamaura Reference Maeda and Yamaura1992; Fukuda and Maeda Reference Fukuda and Maeda1998; Nishimoto Reference Nishimoto2000). With regard to the marine environment around Rebun Island, the Soya Warm Current, a branch of the Tsushima Warm Current, passes from the Japan Sea through the Soya Strait and into the Okhotsk Sea and then flows southeast along the northeast coast of Hokkaido (see Figure 1). Todo Island, where historically a pinniped rookery was located, lies just north of Rebun Island (Maeda and Yamamura Reference Maeda and Yamaura1992; Nishimoto Reference Nishimoto2000; Sakaguchi Reference Sakaguchi2007).
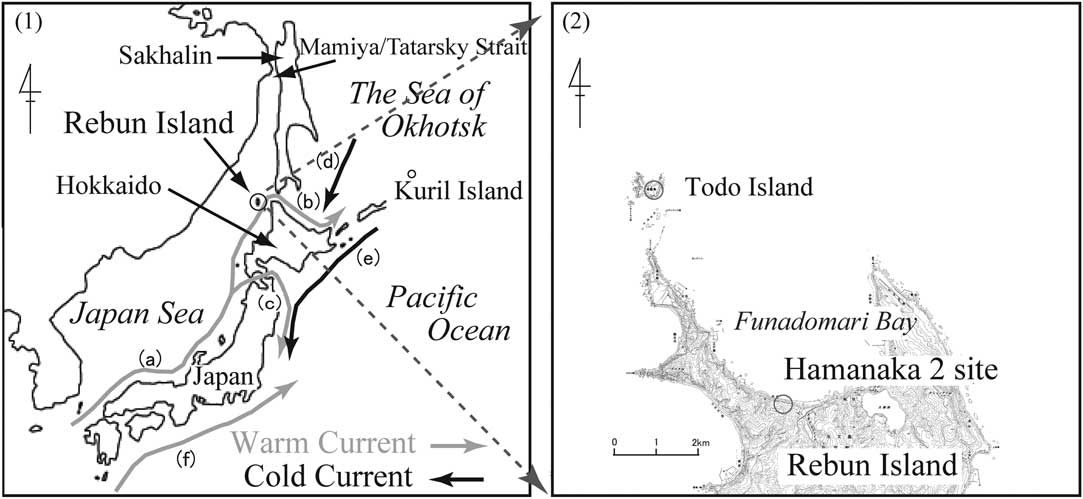
Figure 1 (1) Japanese Archipelago, together with arrows labeled with letter indicate currents: (a) Tsushima Current, (b) Soya Warm Current, (c) Tsugaru Warm Current, (d) East Sakhalin Current, (e) Oyashio Current, (f) Kuroshio Current. (2) Hamanaka 2 archaeological site, Rebun Island, Hokkaido, Japan. This figure is based partly on a 1:25,000 scale topographic map (Rebun; NL-54-16-16-3) made by the Geographic Survey Institute, Japan.
Hamanaka 2 is an archaeological midden site situated on the Hamanaka sand hill, which faces Funadomari Bay to the north. The research area (location R) is about 150 m from the present-day shoreline (Figure 2). The archaeological remains at this site have very likely been preserved because of the close packing of the sand. The target of this study was layer V. Layer V is composed of black sand about 20 cm thick, and it has been dated to the latter half of the Late Jomon period (1300–1200 cal BC; Miyata et al. Reference Miyata, Horiuchi and Nishimoto2009). The base of layer V is ~5 m above sea level (Figure 2). This layer contains traces of more than 34 fire pits on the low parts of the sand dune that were used repeatedly, but no pit dwellings, unlike Funadomari 1, a settlement site about 2 km from Hamanaka 2 (Nishimoto Reference Nishimoto2000). A large quantity of sea mammal bones (especially Japanese sea lion) were found in this layer, along with about 3000 potsherds from large pots and a large quantity of stone flakes. Only a few bones from other mammals or birds occur in the layer, and fish bones and molluskan shells are also rare. Almost all of the excavated potsherds are of the Doubayashi pottery type, which means that pottery was used at the site for a relatively short time, at most several decades, during the latter half of the Late Jomon period (Nishimoto Reference Nishimoto2000; Miyata et al. Reference Miyata, Horiuchi and Nishimoto2009). People probably occupied Hamanaka 2 only during summer, when it may have been used as a temporary campsite for hunting Japanese sea lions (Zalophus californianus japonicus), and for cooking (boiling) the catch and processing the animal oils and fats in pottery vessels (Nishimoto Reference Nishimoto2000; Kami Reference Kami2001). Such sites are known by archaeologists as “kill sites” or “processing sites” (Rackham Reference Rackham1994; Nishimoto Reference Nishimoto2000).
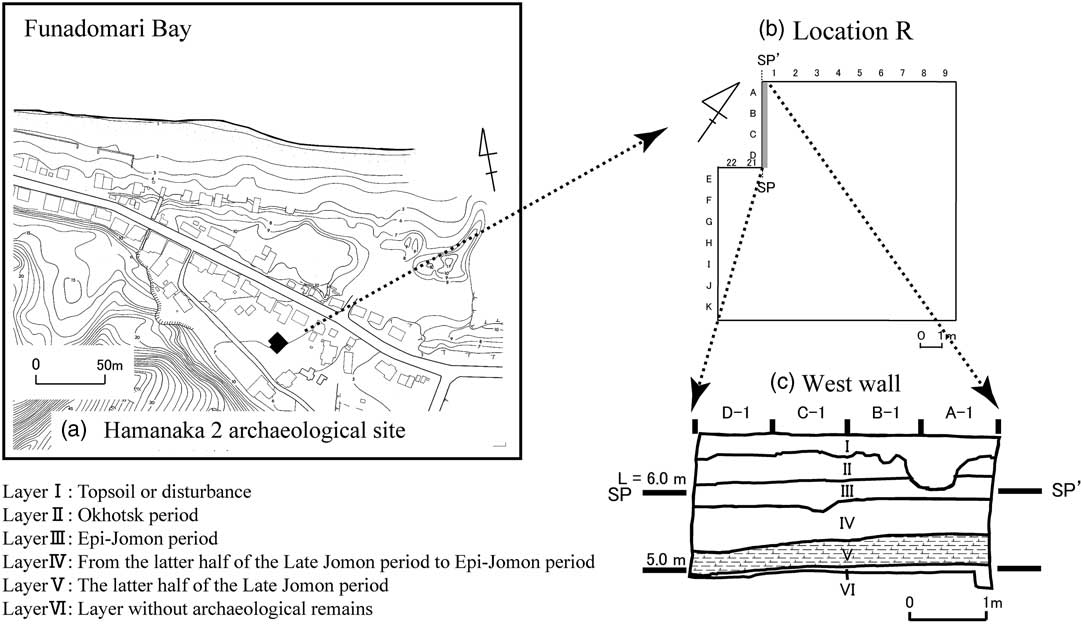
Figure 2 (a, b) Location R, Hamanaka 2, Rebun Island, Japan. (c) Archaeological stratigraphy showing the position of layer V. This figure is from Miyata et al. (Reference Miyata, Horiuchi and Nishimoto2009), which was modified from Nishimoto (Reference Nishimoto2000).
Miyata et al. (Reference Miyata, Horiuchi and Nishimoto2009) reconstructed the prehistoric human diets from the stable isotope compositions (δ13C, δ15N) of carbonized matter adhering to the inner surfaces of pottery excavated from layer V, location R, at the Hamanaka 2 site (Figures 1 and 2). The lipids adsorbed to the pottery matrix were also analyzed. More than 80% of the detected sterols consisted of cholesterol, and this suggests that they were of animal origin. The stable isotope compositions indicate that the food residues probably derived from marine mammals or fish. The 14C ages of charred materials from the inner surface of potsherds from Hamanaka 2 show marine reservoir effects, which support the interpretation that they originated from marine products cooked in the pots. The apparent 14C age differences between the charred materials on the inner surfaces of the potsherds and charred wood samples from the same archaeological context indicate a correction value for the marine reservoir effect (∆R) in the northwestern Pacific of 444±55 14C yr (±1σ) for materials with a 14C age of 3010 BP (Miyata et al. Reference Miyata, Horiuchi and Nishimoto2009). The results of sterol, stable isotope analyses, and 14C dating are consistent with the archaeological hypothesis that sea mammals were cooked in pottery vessels to obtain animal oils and fats at the Hamanaka 2 archaeological site during the latter half of the Late Jomon period (1300–1200 cal BC).
MATERIALS AND METHODS
Molluskan shells, animal bones, and charred materials from the inner surfaces of pottery that were used in this study were all from layer V, Location R at the Hamanaka 2 archaeological site. Five marine molluskan shell samples were analyzed. A section of the outside edge of the shells was cut with a diamond cutter and ultrasonically cleaned in Milli-Q water and acetone. Then, the sample was leached by HCl to remove any contamination remaining after the physical cleaning. The purified shell samples were reacted with 100% phosphoric acid (Hoefs Reference Hoefs1987; Suzuki et al. Reference Suzuki, Kawahata, Tanimoto, Tsukamoto, Gupta and Yukino2000, Reference Suzuki, Gagan, Fabricius, Isdale, Yukino and Kawahata2003) and converted to graphite at the Radiocarbon Dating Materials Laboratory, NMJH (Miyata et al. Reference Miyata, Sakamoto, Ozaki, Shinmen, Onbe, Nishimoto and Imamura2005). Two animal bone samples were also measured in this study. One was a shin bone of a Japanese sea lion (Zalophus californianus japonicus), a species that is now extinct (Wada and Ito Reference Wada and Ito1999). The other was an Alcidae arm bone, from either a common murre (Uria aalge) or thick-billed murre (Uria lomvia); the sample was too small to identify the species with certainty. The Alcidae are a family of fish-eating birds that can typically dive up to about 50 m to capture prey, but they also eat small fish and squid from the surface layer. Finely powdered bone sample was demineralized in 1M HCl. The acid-insoluble fraction was treated with 0.1M NaOH to remove humates and lipids. The gelatin fraction was extracted by heating the residue with 0.001M HCl at 90°C for 10 hr, and then freeze-dried (Kondo et al. Reference Kondo, Matsu’ura, Nakai, Nakamura and Matsui1992; Matsu’ura and Kondo Reference Matsu’ura and Kondo2001; Kondo and Matsu’ura Reference Kondo and Matsu’ura2005). It was converted to graphite at Paleo Labo Co., Ltd (Gifu, Japan). The graphite samples from the molluskan shells and bone collagen were measured with an NEC Compact AMS at Paleo Labo Co. Ltd., Gunma, Japan (Kobayashi et al. Reference Kobayashi, Niu, Itoh, Yamagata, Lomtatidze, Jorjoliani, Nakamura and Fujine2007). We also measured the stable carbon and nitrogen isotope compositions and contents of the bone collagen at the University Museum, University of Tokyo, Japan. These analyses were performed with an elemental analyzer and mass spectrometer linked together by a continuous-flow system (EA3028-HT and Micromass IsoPrime IRMS). The stable carbon and nitrogen isotope results are reported in per mil notation relative to V-PDB for δ13C and air for δ15N (Kunikita et al. Reference Kunikita, Yoshida, Miyazaki, Saito, Endo, Matsuzaki, Ito, Kobayashi, Fujimoto, Kuznetsov, Krupyanko and Tabarev2007).
The marine reservoir age (R) is calculated as the difference between the observed 14C age and the atmospheric (terrestrial) age of materials of the same calendar age (Stuiver et al. Reference Stuiver, Pearson and Braziunas1986; Stuiver and Braziunas Reference Stuiver and Braziunas1993; Southon et al. Reference Southon, Kashgarian, Fontugne, Metivier and Yim2002). The local marine reservoir correction value (∆R) is defined as the difference between the observed 14C age and the model marine age for the same calendar age. However, in this work, we were not able to obtain the real calendar age of the samples because they were archaeological remains of unknown age. Thus, we determined the calendar age of the sample assemblages by calibration of measured 14C ages of terrestrial materials based on IntCal13 (Reimer et al. Reference Reimer, Bard, Bayliss, Beck, Blackwell, Bronk Ramsey, Grootes, Guilderson, Haflidason, Hajdas, Hatté, Heaton, Hoffmann, Hogg, Hughen, Kaiser, Kromer, Manning, Niu, Reimer, Richards, Scott, Southon, Staff, Turney and van der Plicht2013) and then used the calibrated ages to estimate the model marine ages of the measured samples based on Marine13 (Reimer et al. Reference Reimer, Bard, Bayliss, Beck, Blackwell, Bronk Ramsey, Grootes, Guilderson, Haflidason, Hajdas, Hatté, Heaton, Hoffmann, Hogg, Hughen, Kaiser, Kromer, Manning, Niu, Reimer, Richards, Scott, Southon, Staff, Turney and van der Plicht2013). Finally, the marine model age was subtracted from the observed 14C age to evaluate ∆R (Southon et al. Reference Southon, Rodman and True1995; Nakamura et al. Reference Nakamura, Nishida, Takada, Okuno, Minami and Oda2007). The uncertainty of the ∆R value was estimated by the square root of the sum of squared uncertainties of the model marine age and the measured marine age. The 14C ages and R and ∆R values of the archaeological remains excavated from the site are shown in Table 1 together with the carbon and nitrogen isotope compositions, C/N ratio, gelatin yield, and recovery rate of the bone collagens. In this study, the charred wood fragments (terrestrial materials) used for R and ∆R calculation were collected from the same horizon (layer V) at Hamanaka 2, where marine samples were excavated (Miyata et al. Reference Miyata, Horiuchi and Nishimoto2009).
Table 1 Radiocarbon ages of archaeological remains excavated from layer V, location R, Hamanaka 2 site.
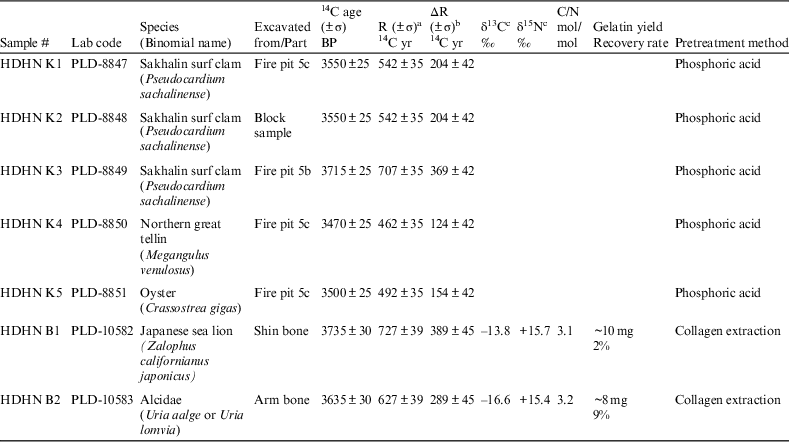
a R was obtained by subtracting the average age of charred wood (3008±25 BP; Miyata et al. Reference Miyata, Horiuchi and Nishimoto2009) from the observed 14C age.
b ΔR was obtained by subtracting the marine model 14C age (3346±34 BP), which corresponded to the age of the charred wood [3008±25 BP; 1375-1130 cal BC (95.4%)] from the observed 14C age.
c Measurement error was±0.1‰.
The average 14C ages, R, and ∆R of various species used to evaluate the marine reservoir effect of various archaeological remains are summarized in Table 2 (Miyata et al. Reference Miyata, Horiuchi and Nishimoto2009). As described in the next section, Sample HDHN K3 was excluded from the calculation of the average.
Table 2 Radiocarbon ages of various types of archaeological remains excavated from layer V, location R at the Hamanaka 2 site.

We also measured the 14C ages and C/N ratios of charred materials from the inner surfaces of potsherds after each cleaning step during the acid-alkali-acid (AAA) treatment to determine their origin (Table 3 and Figure 6). The charred materials were prepared as previously described (Mook and Streurman Reference Mook and Streurman1983; Miyata et al. Reference Miyata, Horiuchi and Nishimoto2009, Reference Miyata, Minami, Onbe, Sakamoto, Matsuzaki, Nakamura and Imamura2011). In brief, the charred materials taken from the potsherds were mixed well and washed with an ultrasonic using acetone. Each sample, then, was reacted 2 or 3 times with 1N HCl, 1 or 2 times with 0.1N NaOH, 4 or 5 times with 1N NaOH, and twice with 1N HCl for 1 hr each time at 80°C. The purified sample was then neutralized, washed with pure H2O, filtered, and dried. The dried sample was sent to Paleo Labo, Inc., Gifu, Japan, where it was converted to CO2 gas and then reduced to graphite. The graphite from the charred material was measured to determine the 14C age with the Compact AMS at Paleo Labo, Co., Ltd., Gunma, Japan (Kobayashi et al. Reference Kobayashi, Niu, Itoh, Yamagata, Lomtatidze, Jorjoliani, Nakamura and Fujine2007). The carbon and nitrogen isotope compositions and contents of the carbonized materials after each step of the AAA cleaning treatment were measured by SI Company Co., Ltd., Saitama, Japan.
Table 3 Radiocarbon ages, carbon and nitrogen content, and C/N ratios of charred materials from inner surfaces of potsherds from layer V, measured after each step of the AAA treatment.
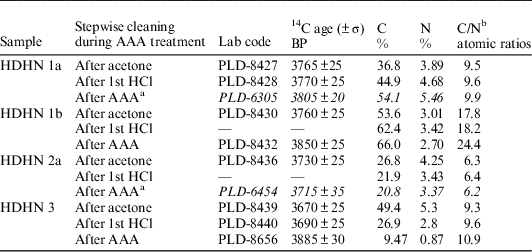
a From Miyata et al. (Reference Miyata, Horiuchi and Nishimoto2009).
b The standard errors of the C/N ratio were typically within 0.1.
RESULTS AND DISCUSSION
ΔR Values of Soya Warm Current Deduced from Marine Molluskan Shells
When we arrange dated samples of the various types of archaeological remains according to the expected degree of marine influence (Table 2), we find that as the marine influence increased, the marine reservoir effect gradually increased (see Figure 3).
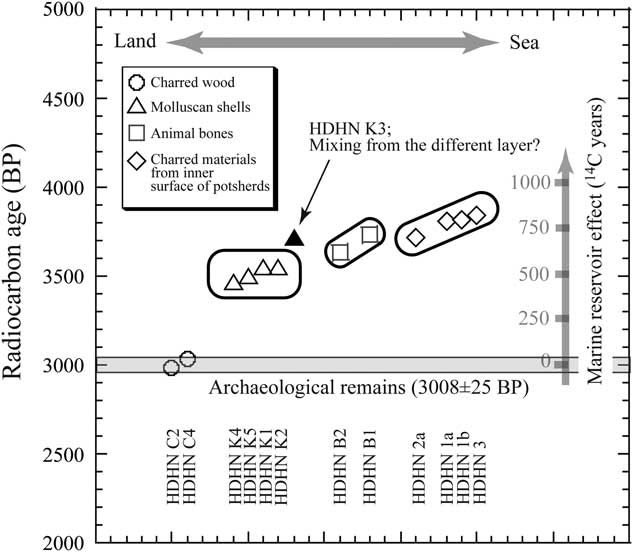
Figure 3 Radiocarbon ages of archaeological remains from the Hamanaka 2 site.
To identify the influence of the Soya Warm Current, which flows around Rebun Island (see Figure 1), on the marine reservoir effect, we measured the 14C ages of five molluskan shells (Table 1). During shell calcification, mollusks utilize the DIC in seawater. As a result, the isotopic composition of the carbonate of the accretionary growth point at the tip of the shell reflects the 14C activity in local surface water. The marine reservoir effect was evaluated by measuring the apparent 14C age difference between the molluskan shells and the charred wood, whose 14C activity corresponded to that of the coeval atmosphere (Miyata et al. Reference Miyata, Horiuchi and Nishimoto2009).
Although the molluskan shells are from different species, the ages of three of the five (HDHN K1, K4, and K5) from fire pit 5c were similar within error (3507±40 BP; n=3). The 14C age of sample HDHN K2 (3550±25 BP), from a block sample of layer V, was within the same range, whereas that of HDHN K3 (3715±25 BP) from fire pit 5b was 200 yr older than the average age of the other shells (3518±39 BP; n=4; Table 2). However, stratigraphically, fire pit 5b is directly in contact with and above fire pit 5c. This stratigraphic inconsistency with the dates can be explained if a molluskan shell (sample HDHN K3) from a horizon below layer V became mixed with younger materials in fire pit 5b. Therefore, HDHN K3 was inferred to be from an older stratigraphic layer and excluded from the calculation of the average age.
ΔR for the Soya Warm Current is obtained by subtracting the marine model 14C age (3346±34 BP), which corresponded to the age of the charred wood fragments [3008±25 BP; 1375–1130 cal BC (95.4%); Miyata et al. Reference Miyata, Horiuchi and Nishimoto2009], from the observed 14C age, so that ∆R for the Soya Warm Current is 172±39 14C yr (n=4; Table 2) at the 14C age 3008±25 BP. This value is a slightly larger than the previously reported pre-bomb ∆R (94±43 14C yr; n=1; marine shell value collected in 1899 at Otaru Bay, Hokkaido; Yoneda et al. Reference Yoneda, Uno, Shibata, Suzuki, Kumamoto, Yoshida, Sasaki, Suzuki and Kawahata2007). This discrepancy may be due to the difference in sampling time and the complicated oceanic circulation around Hokkaido, where water masses originated from the East Sakhalin Current (∆R=399±22 14C yr; n=2; Yoneda et al. Reference Yoneda, Uno, Shibata, Suzuki, Kumamoto, Yoshida, Sasaki, Suzuki and Kawahata2007), which flows from the Okhotsk Sea north of Hokkaido, are substantially depleted in 14C (“old” in 14C terms), whereas water masses from the Soya Warm Current are “younger” and have smaller reservoir ages (see Figure 1; Yoneda et al. Reference Yoneda, Uno, Shibata, Suzuki, Kumamoto, Yoshida, Sasaki, Suzuki and Kawahata2007). Moreover, the ∆R value of the Soya Warm Current may change because of fluctuations of the Tsushima Warm Current (from which the Soya Warm Current originates) during the Holocene on a timescale of 103 yr (see Figure 1; Koizumi Reference Koizumi1987, Reference Koizumi1989; Kito et al. Reference Kito, Noda and Minami1998; Matsushima Reference Matsushima2010). A similar tendency is also observed in the Δ14C values of modern surface water (after bomb) between “old” Okhotsk water around Kuril island (Δ14C, –40‰ to –20‰, collected in 1998; Aramaki et al. Reference Aramaki, Watanabe, Kuji and Wakatsuchi2001; Figure 1) and “younger” northern Japan Sea water from west of Sakhalin Island (Δ14C, +50‰ to +70‰, collected in 2002; Aramaki et al. Reference Aramaki, Senjyu, Togawa, Otosaka, Suzuki, Kitamura, Amano and Volkov2007; Figure 1) that originated from the Tsushima Warm Current.
Apparent Reservoir Age Difference between Japanese Sea Lion and Alcidae
In this study, by measuring the 14C ages of molluskan shells and animal bones of archaeological remains from the same layer with a clear archaeological context, we investigated the relationship between the marine reservoir effect and the type of material being dated. The marine reservoir effects shown by the dates of these remains reflect the overall diet of the animals, including the effects of the ocean currents and the marine food web when the animals were alive. Thus, we can estimate how the remains were influenced by the marine environment through the marine reservoir effects.
Table 1 shows the 14C ages obtained for individual shellfish and animal bone collagen together with the calculated R and ∆R values. δ13C and δ15N and the carbon to nitrogen atomic ratio (C/N) of Japanese sea lion and Alcidae bone collagens are also listed. The similar C/N ratios of Japanese sea lion and Alcidae indicate that the purified collagens had not been affected by diagenesis (DeNiro Reference DeNiro1985; Ambrose Reference Ambrose1990; van Klinken Reference van Klinken1999).
The 14C age and ∆R of Japanese sea lion (HDHN B1) were 3735±30 BP and 389±45 14C yr, respectively, and those of Alcidae (HDHN B2) were 3635±30 BP and 289±45 14C yr, respectively (Tables 1 and 2). The apparent difference in ∆R between Japanese sea lions and Alcidae probably reflects dietary differences, as discussed below.
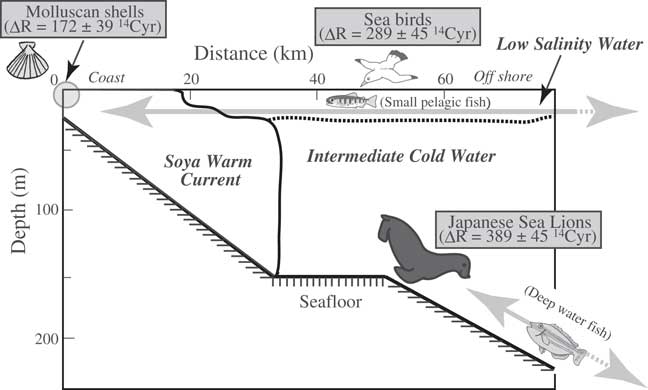
Yoneda et al. (Reference Yoneda, Tanaka, Shibata, Morita, Uzawa, Hirota and Uchida2002) estimated changes in the marine reservoir age in the northwestern Pacific during the period from 4900 to 800 BP from the apparent age differences between terrestrial (deer) and marine (fur seal) mammals excavated from different, but neighboring, archaeological deposits. In this study, we compared the 14C ages of molluskan shells and animal bones from the same layer, which also contained cultural remains. Because the Japanese sea lion is now extinct, its ecology is unknown. However, the diet of Japanese sea lions was reported to be similar to that of Steller’s sea lions (Eumetopias jubatus), and different from that of the northern fur seal (Callorhinus ursinus) (Wada and Ito Reference Wada and Ito1999). Northern fur seals migrate across the open sea and eat small pelagic fish in the surface layer, whereas Steller’s sea lions, which prefer coastal waters and do not migrate very far, eat fish that live in the deep sea (Wada and Ito Reference Wada and Ito1999). Moreover, Steller’s sea lions lived year round and bred on Todo Island, off the northern end of Rebun Island, until the 1950s (Wada Reference Wada2004; see Figure 2). Therefore, Japanese sea lions, which also ate deep-sea fish, may also have lived in coastal waters around Rebun Island year round. The difference in ∆R between Japanese sea lions and Alcidae probably reflects their different diets (Figure 4). The influence of the Soya Warm Current, which flows just a short distance north of Hokkaido, is limited to shallow waters (Aota Reference Aota1975; Takizawa Reference Takizawa1982; Matsuyama et al. Reference Matsuyama, Wadaka, Abe, Aota and Koike2006). Thus, mollusks that live in shallow coastal habitats are strongly affected by the “younger” Soya Warm Current. Further offshore, the surface waters are from the Okhotsk Sea, which are characterized by extremely large reservoir ages (Figure 1; Aramaki et al. Reference Aramaki, Watanabe, Kuji and Wakatsuchi2001; Yoneda et al. Reference Yoneda, Uno, Shibata, Suzuki, Kumamoto, Yoshida, Sasaki, Suzuki and Kawahata2007). Deep-water fish, which are limited to environments well north of the coast, are “old” (in 14C terms) because they feed (in deep water) on sinking particles derived mainly from zooplankton. When they were alive, the zooplankton fed on phytoplankton living and photosynthesizing in the “old” Okhotsk surface water. Small pelagic fishes migrate between coastal and offshore waters (Figure 4) and may therefore have 14C values intermediate between those of coastal mollusks and deep-water fish. Consequently, the ΔR value of Alcidae, which eat the small pelagic fish, is between the ΔR values of Japanese sea lions and molluskan shells (see Figure 4).
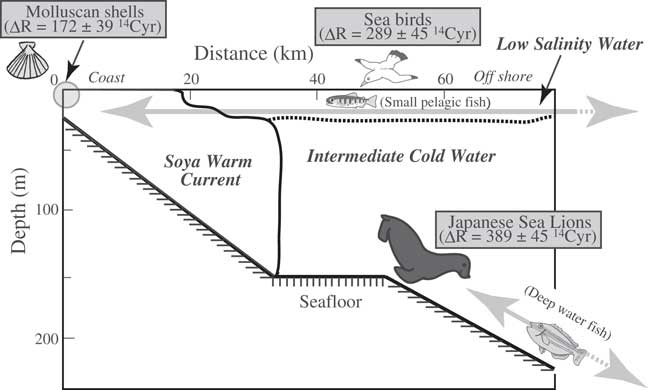
Figure 4 Schematic diagram of the water-mass structure in the Sea of Okhotsk in summer, showing the relationship between the habitats of various fish and shellfish and the animals that prey on them. Modified from Aota (Reference Aota1975).
With regard to the bone collagen of Japanese sea lions and Alcidae, we assumed that the carbon and nitrogen isotopic fractionation between the proteins in their food and in their bone collagen was approximately the same. Investigations of marine food webs have shown that δ15N values in tissues of a consumer species are well correlated with its trophic level, with higher δ15N values being associated with organisms at higher trophic levels (Schoeninger and DeNiro Reference Schoeninger and DeNiro1984; Wada et al. Reference Wada, Terazaki, Kabaya and Nemoto1987). Japanese sea lions and Alcidae presumably belong to the same trophic level because their δ15N values are almost identical (δ15N values of Japanese sea lion and Alcidae are +15.4‰ and +15.7‰, respectively). This result suggests that the Alcidae consume marine fish and mollusks, similar to Japanese sea lions.
14C Age Differences between Japanese Sea Lion Bone Collagen and the Charred Material on the Inner Surface of a Potsherd
The results of sterol analysis, stable isotope analysis, and 14C dating of charred materials from the inner surfaces of potsherds from layer V (Miyata et al. Reference Miyata, Horiuchi and Nishimoto2009) are consistent with the archaeological hypothesis that sea mammals were cooked in pots to obtain animal oils and fats at the Hamanaka 2 site during the latter half of the Late Jomon period (1300–1200 cal BC). The 14C ages of four charred food crust samples ranged from 3715±35 BP (sample HDHN 2a) to 3840±35 BP (sample HDHN 3), and averaged 3794±55 BP (n=4). The calculated local reservoir value (ΔR) ranged from 368±44 14C yr (HDHN 2a) to 493±44 14C yr (HDHN 3), with an average of 447±55 14C yr at 3008±25 BP (n=4) (Miyata et al. Reference Miyata, Horiuchi and Nishimoto2009).
We compared the relationship between the C/N ratio and δ13C of charred materials from the inner surfaces of potsherds with that of possible sources of the charred materials (Figure 5b). The C/N ratio of charred material on the interior surface of one potsherd dated to 3715±35 BP (HDHN 2a) was about 6 (Figure 5b), which is the same as that in soft tissues from muscle and blubber of California sea lions (Zalophus californianus) (Newsome et al. Reference Newsome, Phillips, Culleton, Guilderson and Koch2004). The δ13C value and C/N ratio of sample HDHN 2a both strongly indicate a marine origin. In contrast, the 14C ages of the three other inner surface charred residues (samples HDHN 1a, 1b, and 3) are 100 14C yr older than that of HDHN 2a, and the plots of these samples (Figure 5b) do not lie in the marine animal range but are closer to the regions of non-starchy C3 plants, terrestrial animals, and black soil. This result indicates the three pots on which these residues were found were used to cook both marine animals and terrestrial organisms. The same pattern is observed in the δ15N versus δ13C relationships (Figure 5a). Thus, HDHN 2a likely originated from a marine animal, and the other three samples might be a mixture of marine products with small amounts of residues derived from C3 plants or terrestrial animals. However, if the samples HDHN 1a, 1b, and 3 are charred residues of marine animals and C3 plants, their 14C ages might be younger than the 14C age of HDHN 2a, which is only of marine origin, but the three samples showed older ages. Therefore, the mixing with C3 plants is inconsistent with 14C results.
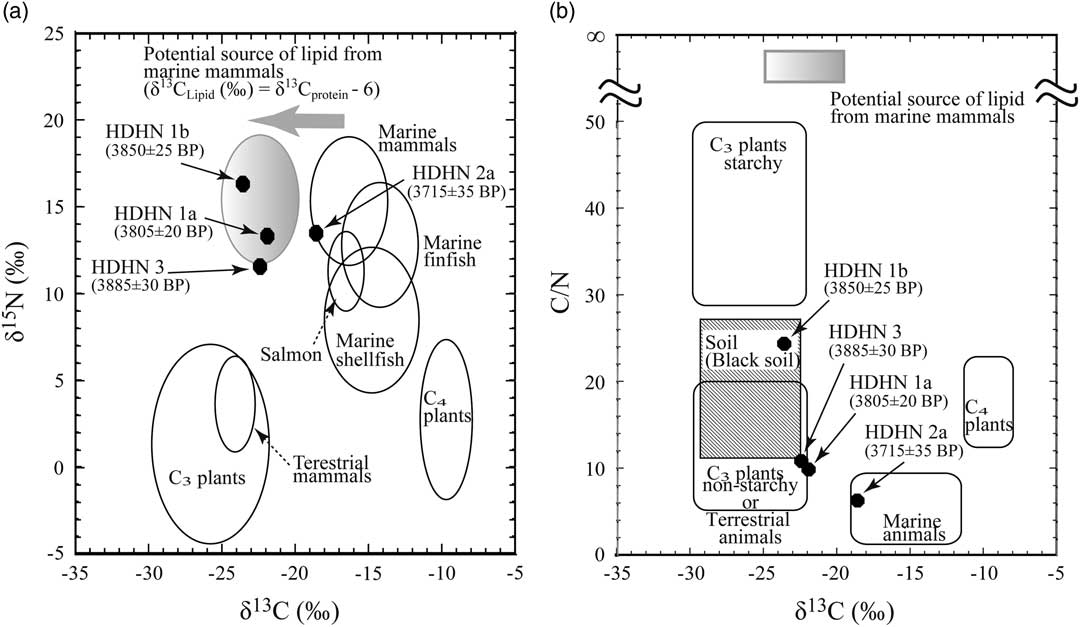
Figure 5 (a) Possible sources of the charred materials on the interior surfaces of potsherds at Hamanaka 2 according to their δ13C and δ15N values, together with the potential source of lipid from marine mammals. The end-member regions are modified from Yoshida et al. (Reference Yoshida, Kunikita, Miyazaki, Nishida, Miyao and Matsuzaki2013). (b) Possible sources of the charred materials on potsherds at Hamanaka 2 according to their δ13C values and C/N ratios, together with the potential source of lipid from marine mammals. The end-member regions are modified from Yoshida et al. (Reference Yoshida, Kunikita, Miyazaki, Nishida, Miyao and Matsuzaki2013).
Accordingly, we consider mixing with lipids from marine mammals. There are differences in the carbon isotopic composition of the major biochemical compositions of an organism, in general. The lipid fraction has a lower δ13C value (~6‰) than the other major components such as protein and carbohydrate fractions (DeNiro and Epstein Reference DeNiro and Epstein1977). In the marine food web structure, the blubber of marine mammals contains large amount of fat in tissue has about 6‰ lower carbon isotopic composition than that of protein, but so slightly nitrogen that, show the C/N ratios more than 2000 (McConnaughey and McRoy Reference McConnaughey and McRoy1979). Then, as shown in Figure 5, we consider the effect from the potential source of lipid from marine mammals as one of the end-members in this study, based on the archaeological hypothesis that sea mammals were cooked in pottery vessels to obtain animal oils and fats at the Hamanaka 2 archaeological site during the latter half of the Late Jomon period (1300–1200 cal BC) (Nishimoto Reference Nishimoto2000).
If there is a potential source of lipids from marine mammals, it is likely to be shown in the stable isotopic compositions of the three samples, other than HDHN 2a, as shown in Figure 5a. Furthermore, Figure 5b shows that the mixing marine animals with the potential source of lipids is likely to explain the relationship between C/N ratio and δ13C values of the three charred residues, except for HDHN 2a, without mixing with C3 plants or terrestrial animals.
We examined the C/N ratios and 14C ages of charred materials after each step of the AAA treatment because we expected the stability of the charred materials during the AAA treatment would provide insight into the origin of charred materials from the inner surfaces of potsherds (Table 3). The C/N ratio of 6 and 14C age of HDHN 2a were changed only slightly by the AAA treatment (Mook and Streurman Reference Mook and Streurman1983; Miyata et al. Reference Miyata, Horiuchi and Nishimoto2009, Reference Miyata, Minami, Onbe, Sakamoto, Matsuzaki, Nakamura and Imamura2011). This result indicates that the char was mostly free of any organic materials, due to either incomplete decomposition of the original foodstuff or secondary contaminants. Thus, the HDHN 2a sample consisted of completely charred material derived from tissues of marine organisms. Therefore, we used HDHN 2a as a reference sample in our analysis (see Figures 5 and 6).
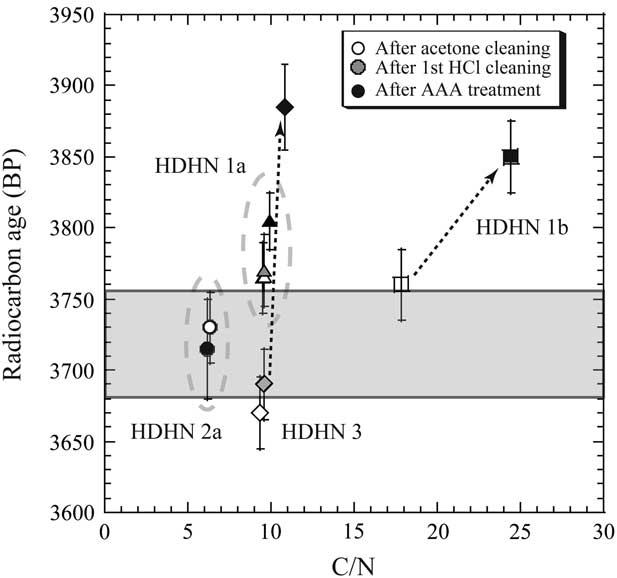
Figure 6 Relationships between 14C ages and C/N ratios of charred materials on potsherds at Hamanaka 2 after each step of the AAA treatment. The shaded area shows the 14C age range of sample HDHN 2a (3715±35 BP) after the AAA treatment.
In contrast, the C/N ratio and 14C age of the other three charred residues increased during the AAA treatment. Secondary contaminants from soil such as humic acid and any organics remaining from the original foodstuff were removed by the NaOH (alkali) cleaning step; thus, carbon with a C/N ratio of less than 10 (sample 1a and 3) or 18 (sample HDHN 1b) and any contaminants with a “young” 14C age were likely removed by this step. The pottery with the analyzed chars were from relatively shallow fire pits dug into a sand hill, not from a deep pit such as a burial pit containing old organics in the fill. Therefore, the carbon in the fill of the abandoned fire pits was likely “young,” rather than “old,” carbon.
The three samples originating from different pottery vessels, other than HDHN 2a, also may have comprised single end-member from mainly “old” marine products (suggesting a larger reservoir effect than that of HDHN 2a), free from a small amount of “young” carbon from C3 plants or terrestrial animals. Therefore, the chemical compositions of all samples may have been consistent with the archaeological hypothesis that sea mammals were cooked in pottery vessels to obtain animal oils and fats at the Hamanaka 2 archaeological site during the latter half of the Late Jomon period (1300–1200 cal BC). Therefore, we infer that ΔR for the bone collagen of Japanese sea lion was less than or equal to that for the charred materials from the inner surface of potsherds.
Comparisons of Marine Reservoir Correction Values, ∆R
We compared the ΔR values of molluskan shells and animal bones with clear archaeological contexts. One sample of charred material (HDHN 2a) was preliminarily inferred to have the original chemical composition of the sea mammal from which it derived (a constant C/N ratio of 6 during AAA treatment), and its 14C age agreed well with that of Japanese sea lion bone collagen, suggesting that the marine reservoir effect was similar in both. However, the 14C ages of the other samples of charred materials were older, presumably because they derived at least in part from organisms with a larger marine reservoir effect. Therefore, on the basis of these data and the 14C ages on shells and bones, we infer the following relationships regarding ΔR values in this study: molluskan shells (Soya Warm Current)<Alcidae bone<Japanese sea lion bone≤charred materials from the inner surface of potsherds (Table 2, Figure 3). When we arranged the dated samples in order of the inferred degree of marine influence (Figure 3), the degree of marine influence increased as the marine reservoir effect increased. Samples appearing out of order (e.g. sample HDHN K3) probably do so either for some reason related to the dated material itself or because they originated from a different layer than that containing the other dated remains. Therefore, the results shown in Figure 3 support the inference that sample HDHN K3 was originally from a layer beneath the layer in which it was found (layer V) and was not contemporary with the other dated remains.
CONCLUSIONS
We measured the 14C ages of two kinds of archaeological remains, molluskan shells and animal bones, from layer V, location R, at the Hamanaka 2 archaeological site, Rebun Island, Japan, which was occupied during the latter half of the Late Jomon period (1300–1200 cal BC). This layer had a clear archaeological context, and it contained potsherds from pots that may have been used to boil and cook sea mammals.
We inferred differences in the diets and habitats of molluskan shellfish, birds, and sea mammals from the apparent 14C age differences caused by marine reservoir effects. We estimated the ∆R value of the Soya Warm Current to be 172±39 14C yr (n=4) from the ages of molluskan shells.
The 14C age of collagen extracted from a Japanese sea lion bone agreed well with that of charred material from the inner surface of one potsherd. We infer that the charred material derived without substantial chemical alteration from Japanese sea lion. This result is also consistent with the archaeological hypothesis that sea mammal meat was cooked in pots at the Hamanaka 2 site. Three samples of charred residues from the inner surfaces of pots with marine reservoir ages 100 yr older, possibly consisted mainly of single end-member from “old” marine products; however, further investigations, such as lipid analysis and compound-specific carbon isotope analysis (e.g. δ13C16:0, δ13C18:0), are needed to confirm the origin of these charred materials.
We obtained the following 14C age relationships: charred wood<marine shells<Alcidae<Japanese sea lion≤charred materials from the inner surfaces of potsherds. Japanese sea lions may have eaten fish living at great depth that early humans could not consume directly, and the small pelagic fishes from the surface layer eaten by seabirds such as Alcidae are unlikely to be preserved in the archaeological record. Therefore, 14C dating and stable isotope analyses of bones of sea mammals and seabirds found at archaeological sites, and of charred materials from the inner surface of potsherds from the same sites, may allow us to characterize the contemporary marine environment. Here, we inferred the water-mass structure in summer of the Sea of Okhotsk, which is near the Hamanaka 2 archaeological site on Rebun Island, and reconstructed the prehistoric diet of the people who used the site and the habitats of the animals excavated there.
ACKNOWLEDGMENTS
We thank Ms N Kami and Dr S Matsu’ura for their helpful discussions; Dr M Imamura, Dr M Mikawa, Dr I Hajdas, Dr D Kunikita, and Dr T Aramaki for their critical comments; Ms S Kobayashi for her support with sample treatment; Mr T Shinmen and Ms I Nanbu for their help with 14C dating; and Ms Y Miyazaki for her help with the stable isotope analysis. We also thank Dr J Southon for his constructive review comment on first submission and two anonymous reviewers for improvement of the manuscript. This work was supported by Grants-in-Aid for Scientific Research (B) Nos. 25282072 (YM) and 15H03262 (KY), Scientific Research (A) No. 16H01824 (YM) and Scientific Research on Innovative Areas No. 15H05969 (MY) from the Japan Society for the Promotion of Science.









 Open access
Open access