



Introduction
Spirocerca lupi (Spirurida: Spirocercidae) is a parasitic nematode that causes spirocercosis in canids and is associated with canine oesophageal sarcomas (Mazaki-Tovi et al., Reference Mazaki-Tovi, Baneth and Aroch2002; van der Merwe et al., Reference Van der Merwe, Kirberger, Clift, Williams, Keller and Naidoo2008; Rojas et al., Reference Rojas, Dvir and Baneth2020). Its indirect life cycle requires two obligatory hosts, canid as definitive hosts and dung beetles as intermediate hosts. Moreover, a variety of beetle-eating small animals can serve as paratenic hosts (Bailey et al., Reference Bailey, Cabrera and Diamond1963; van der Merwe et al., Reference Van der Merwe, Kirberger, Clift, Williams, Keller and Naidoo2008; Rojas et al., Reference Rojas, Dvir and Baneth2020). The infective third-stage larvae in intermediate and paratenic hosts are similar, except for differences in their body length and size of the internal organs (Sen & Anantaraman, Reference Sen and Anantaraman1971), and are recognized by two cephalic horns at the anterior end and rosette spinous processes at the posterior end (Chhabra, Reference Chhabra1968; Sen & Anantaraman, Reference Sen and Anantaraman1971; Chhabra & Singh, Reference Chhabra and Singh1972). However, the cephalic horns and spinous tail are common characteristics of larvae of spirurid nematodes in dung beetle intermediate hosts (Seurat, Reference Seurat1915; Alicata, Reference Alicata1935; Nichols & Gómez, Reference Nichols and Gómez2014), thus misidentification may occur.
Previous studies on S. lupi larvae from intermediate hosts usually reported this species alone (Chowdhury & Pande, Reference Chowdhury and Pande1969; Du Toit et al., Reference Du Toit, Scholtz and Hyman2008; Gottlieb et al., Reference Gottlieb, Markovics, Klement, Naor, Samish, Aroch and Lavy2011; Mohtasebi et al., Reference Mohtasebi, Teimouri, Abbaszadeh Afshar, Mobedi, Abbasian, Totonchian and Mowlavi2021). So far, various dung beetle species of the family Scarabaeidae have been reported as intermediate hosts of S. lupi and vary among different geographical locations from the United States, Africa and Asia (Faust, Reference Faust1928; Ono, Reference Ono1929; Anantaraman & Jayalakshmi, Reference Anantaraman and Jayalakshmi1963; Bailey et al., Reference Bailey, Cabrera and Diamond1963; Chowdhury & Pande, Reference Chowdhury and Pande1969; Bailey, Reference Bailey1972; Chhabra & Singh, Reference Chhabra and Singh1972; Du Toit et al., Reference Du Toit, Scholtz and Hyman2008; Gottlieb et al., Reference Gottlieb, Markovics, Klement, Naor, Samish, Aroch and Lavy2011; Mohtasebi et al., Reference Mohtasebi, Teimouri, Abbaszadeh Afshar, Mobedi, Abbasian, Totonchian and Mowlavi2021). In Asia, the majority of reports are from China, India, Israel and Iran (Faust, Reference Faust1928; Ono, Reference Ono1929; Anantaraman & Jayalakshmi, Reference Anantaraman and Jayalakshmi1963; Chowdhury & Pande, Reference Chowdhury and Pande1969; Chhabra & Singh, Reference Chhabra and Singh1972, Reference Chhabra and Singh1973; Gottlieb et al., Reference Gottlieb, Markovics, Klement, Naor, Samish, Aroch and Lavy2011; Mohtasebi et al., Reference Mohtasebi, Teimouri, Abbaszadeh Afshar, Mobedi, Abbasian, Totonchian and Mowlavi2021). To the best of our knowledge, there has been no report from south-east Asian countries where the warm climate favours for development of the S. lupi nematode and its beetle intermediate hosts. The aim of this study is, therefore, to survey S. lupi larvae in dung beetle hosts in northern Vietnam, where we found adults of this nematode in dogs in a previous study (Hoa et al., Reference Hoa, Hien, Phuc, Lan and Doanh2021). In addition to larvae of S. lupi, we also detected those of an unidentified spirurid species in different or the same individuals of two dung beetle species. Morphological variation and molecular analyses of these larvae are discussed herein.
Materials and methods
Materials
Dung beetles were collected manually while rolling dung balls of animals surrounding the residential area in Dong Hy District (21°41′10″N°105°55′43″E), Thai Nguyen Province, Vietnam, where S. lupi infections have been detected from domestic dogs (Hoa et al., Reference Hoa, Hien, Phuc, Lan and Doanh2021). A total of 280 individuals of dung beetles were collected and transferred to the Department of Parasitology, Institute of Ecology and Biological Resources, Vietnam Academy of Science and Technology, using ventilated plastic boxes. They were identified at the species level following Ek-Amnuay (Reference Ek-Amnuay2008) and Bui et al. (Reference Bui, Dumack and Bonkowski2018). Two species, Catharsius molossus (Linnaeus, 1758) (210 individuals; fig. 1a, b) and Copris szechouanicus Balthasar, 1958 (70 individuals; fig. 1c, d), were identified. They were reared in separate plastic tanks containing crushed soil moistened with tap water daily and were fed with sterilized cow dung every two days (Mukaratirwa et al., Reference Mukaratirwa, Pillay and Munsammy2010).

Fig. 1. Two dung beetle species collected from Dong Hy District, Thai Nguyen Province, Vietnam: (a) and (b) male and female of Catharsius molossus; and (c) and (d) male and female of Copris szechouanicus.
Each beetle was dissected in normal saline solution in a Petri dish (90 mm) and was checked under a stereomicroscope to look for nematode larvae. For positive beetles, larvae were transferred into another Petri dish (35 mm) containing normal saline to wash until clean. Then, the larvae were treated with hot water to be straightened and were observed under a light microscope for initial morphological examination. Morphologically different types of larvae were separated from each other. One to two larvae of each species were preserved in 96% ethanol for molecular analyses, and the others were preserved in 4% formalin for morphological studies in details.
Morphological study
The larvae preserved in 4% formalin were transferred to lactophenol solution in a Petri dish until the body became transparent, then they were placed in a glass slide, covered with a coverslip, and were examined under a light microscope (Nikon Eclipse Ni., code KST2019-KHV01, IEBR) connected to a digital camera and a computer. Measurements of the body and taxonomic internal organs were taken with the aid of NIS-Elements BR software.
Molecular analyses
Four larvae of two morphological types from the two beetle species were used for molecular identification. Larvae preserved in 96% ethanol were washed with sterile distilled water to remove ethanol. Then, DNA was extracted using QIAamp DNA Mini Kit (Qiagen, Valencia, California, USA). A partial fragment (393 base pairs) of the mitochondrial cytochrome c oxidase 1 gene (cox1) was amplified using primers JB3 and JB4.5 (Bowles et al., Reference Bowles, Hope, Tiu, Liu and McManus1993) following the protocol described by Rojas et al. (Reference Rojas, Dvir, Farkas, Sarma, Borthakur, Jabbar, Markovics, Otranto and Baneth2018) and Hoa et al. (Reference Hoa, Hien, Phuc, Lan and Doanh2021). Polymerase chain reaction (PCR) products were electrophoresed on 1.5% agarose gels and visualized using an ultraviolet transilluminator. Positive PCR products were directly sequenced using an ABI 3100 automated sequencer (Applied Biosystems, Waltham, Massachusetts, USA). Nucleotide sequences obtained in this study were deposited in GenBank under accession numbers LC731325 to LC731328.
The obtained sequences in this study were compared with sequences available in GenBank using the Basic Local Alignment Search Tool (https://blast.ncbi.nlm.nih.gov/Blast.cgi). Nucleotide sequences in GenBank with high coverage and identity over 80% were downloaded and analysed in MEGA7 software (Kumar et al., Reference Kumar, Stecher and Tamura2016). The sequence (LC057236) of Crassicauda giliakiana (Spirurida: Tetrameridae) was used as an outgroup for phylogenetic analysis. Multiple sequence alignments were carried out using the ClustalW algorithm. A phylogenetic tree was reconstructed using the maximum likelihood. The best nucleotide substitution model was statistically selected by MEGA7 software (Kumar et al., Reference Kumar, Stecher and Tamura2016). All positions containing gaps and missing data were eliminated. Bootstraps of 1000 replicates were used for the assessment of topology reliability of the trees.
Results
Morphological data
Nematode larvae with two cephalic horns at the anterior end and spines at the posterior end were collected from two beetle species, C. molossus and C. szechouanicus. Two morphological types of the third-stage larvae were recognized. They were distinguished from each other in their body size (fig. 2) and other morphometric data (table 1). The main morphological characteristics of the two types are as follows.
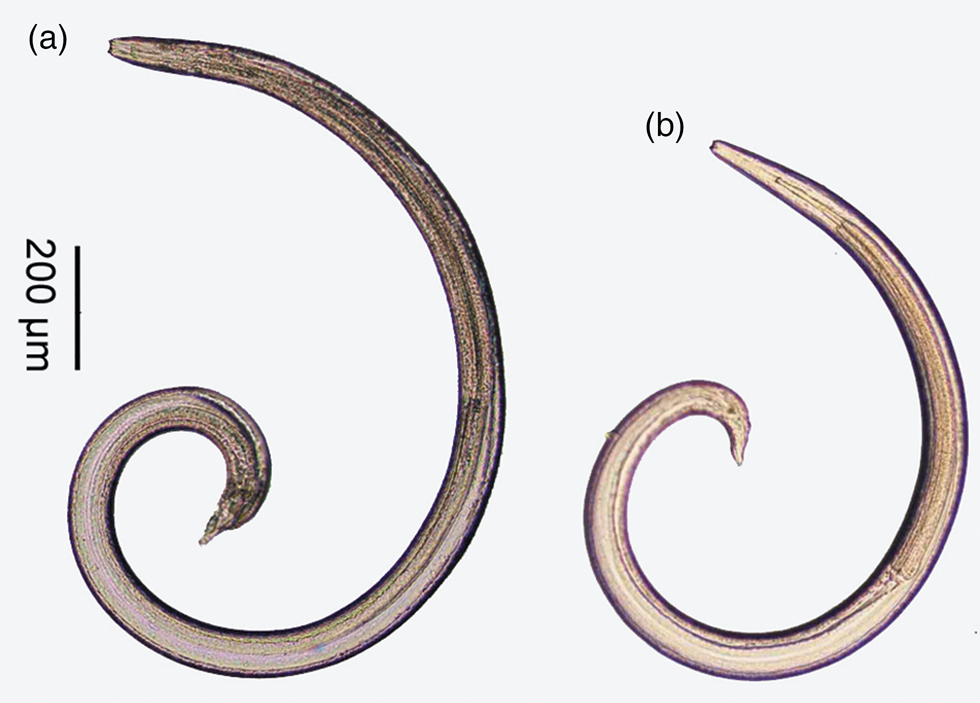
Fig. 2. Whole body of third stage larvae of two spirurid species: (a) Spirocerca lupi; and (b) Physocephalus sp.
Table 1. Comparison of mophometric data of Spirocerca lupi and Physocephalus spp. larvae found in this study and previous report.

Note: data in parentheses indicate average measurements and standard deviation values.
The first larval type (fig. 3a, b): third-stage larvae were found to be free or encysted in various parts of the body of the beetles. Body slender, tapering slightly anteriorly and rather abruptly posterior to anus, 1880–2662 μm in length; the anterior end has two cephalic horns and the posterior end looks like a cup bottom carrying a rosette of closely cuticular spines at the bottom of the tail. The buccal cavity is clear, the oesophagus shows the anterior muscular and posterior glandular parts. The buccal cavity is short, equal to about one-third (26–34%) of the muscular oesophagus. The nerve ring lies at the posterior end of the muscular oesophagus; the excretory pore opens slightly posterior to the nerve ring, around the conjunction between the muscular and glandular oesophagus.
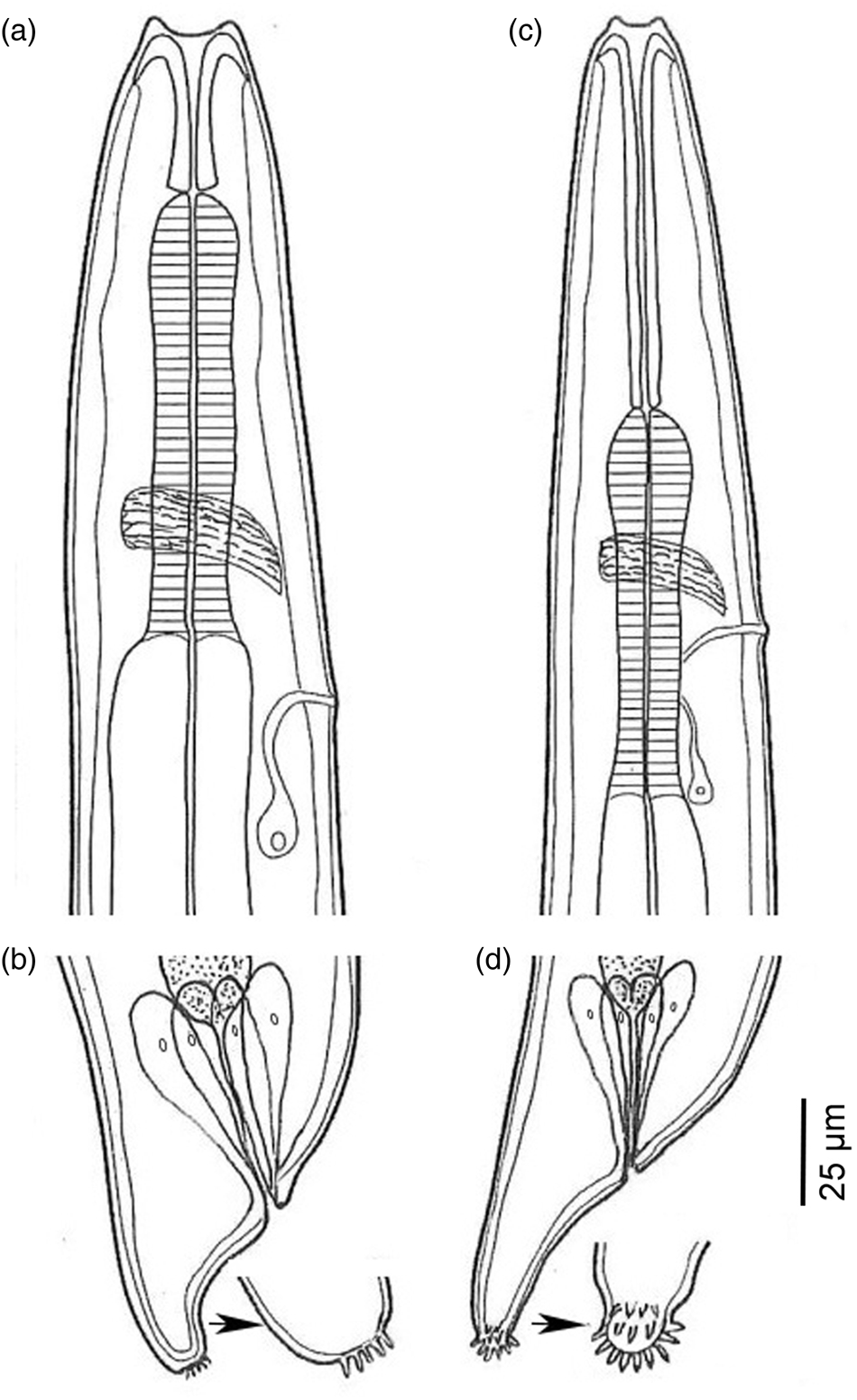
Fig. 3. Anterior and posterior parts of larvae of Spirocerca lupi and Physocephalus sp. showing differences between two species: (a) and (b) anterior and posterior parts of Spirocerca lupi larva; and (c) and (d) anterior and posterior parts of Physocephalus sp. larva.
The second larval type (fig. 3c, d): third-stage larvae were also free or encysted in various parts of the body of the beetles. Body slender, tapering slightly anteriorly and rather abruptly posterior to anus, 1417–1635 μm in length; the anterior end has two cephalic horns and the posterior end is a characteristic small knob, bearing small cuticular spines around the knob tail. The buccal cavity is clear, the oesophagus shows the anterior muscular and posterior glandular parts. The buccal cavity is long, near equal (89–104%) to the muscular oesophagus length. The nerve ring lies at around the middle of the muscular oesophagus; the excretory pore opens at slightly posterior to the nerve ring.
The characteristics of the first type are identical to the S. lupi larva (Seurat, Reference Seurat1915; Chhabra, Reference Chhabra1968) and those of the other are most similar to the Physocephalus sexalatus larva (Seurat, Reference Seurat1915) (table 1).
Molecular analyses
Partial sequences from the cox1 gene were obtained from four larvae of the two types collected from the two beetle species. Two sequences from the larvae, which were morphologically identified as S. lupi, were identical to each other and also identical to S. lupi adults from Vietnam and India, and were highly similar (99.8%) to a Chinese sequence (KC305876), but slightly different (3.3–4.7%) from South African, Israeli and Peruvian sequences, while far distant (8.6%) from a Hungarian sequence (MH634011). In the phylogenetic tree (fig. 4), the cox1 sequences of S. lupi from Vietnam, India and China made a common clade close to the sequences from South Africa, Israel and Peru, while the Hungarian sequence made a separate group. The grouping of genotypes of S. lupi from different geographical origins in the present study is similar to those of previous reports (Rojas et al., Reference Rojas, Dvir, Farkas, Sarma, Borthakur, Jabbar, Markovics, Otranto and Baneth2018; Hoa et al., Reference Hoa, Hien, Phuc, Lan and Doanh2021).

Fig. 4. Phylogenetic tree reconstructed from cox1 sequences by the maximum likelihood method based on the Tamura–Nei model with a discrete Gamma distribution. Bootstrap values higher than 60% based on 1,000 replicates are shown above the nodes. The accession number, species name and geographical origin of each nucleotide sequence are shown. The sequences obtained in this study are printed in boldface type while the others from GenBank database are printed in regular type. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. The scale bar indicated 0.05 changes per nucleotide.
Two sequences from the other species were also completely identical to each other but were not completely identical to any sequences available in GenBank. The result of computing pairwise distances showed that these two cox1 sequences were far distant from sequences of other species, showing closest to Dirofilaria species (Dirofilaria sp. ‘hongkong’, Dirofilaria sp. ‘Thailand’ and Dirofilaria repens), but at high differences from 16.1 to 17.7%. In the phylogenetic tree (fig. 4), they made a distinct group.
Thus, the morphological and molecular data confirmed the first larval type as S. lupi; because the second larval type is highly likely to be P. sexalatus by morphological identification but has not been confirmed by molecular data, we tentatively name it Physocephalus sp.
Prevalence
Both the dung beetle species, C. molossus and C. szechouanicus, were infected with larvae of two nematode species, S. lupi and Physocephalus sp. Prevalences of Physocephalus sp. infection in the two host species (7.1% in both) were slightly higher than those of S. lupi infection (6.2 and 5.7%, respectively). Similarly, intensities of Physocephalus sp. (1–31 larvae per beetle) were slightly higher than those of S. lupi (1–22 larvae per beetle) (table 2).
Table 2. Prevalence of Spirurid larvae in dung bettles from Thai Nguyen Province, Vietnam

Discussion
Dung beetles play an indispensable role as intermediate hosts in the life cycle of spirurid nematodes, including S. lupi (Alicata, Reference Alicata1935; van der Merwe et al., Reference Van der Merwe, Kirberger, Clift, Williams, Keller and Naidoo2008; Nichols & Gómez, Reference Nichols and Gómez2014; Rojas et al., Reference Rojas, Dvir and Baneth2020). In this study, by using morphological and molecular approaches, we confirmed S. lupi larvae in two dung beetle species, C. molossus and C. szechouanicus, as new intermediate hosts. In addition, larvae of another spirurid nematode, Physocephalus sp., were also detected in both beetle species.
Morphologically, although the body size of Physocephalus sp. larvae is smaller than that of S. lupi, it is difficult to distinguish them at low magnification due to their minute size, especially when they are not concurrently present for comparison. Moreover, the body size and other morphometric data of S. lupi larvae vary among reports. Generally, its body length ranges from 1880–3400 μm (Seurat, Reference Seurat1915; Chhabra, Reference Chhabra1968; Mohtasebi et al., Reference Mohtasebi, Teimouri, Abbaszadeh Afshar, Mobedi, Abbasian, Totonchian and Mowlavi2021; this study), longer than that of Physocephalus larvae (940–1728 μm). However, S. lupi larvae from dung beetles reported by Sen & Anantaraman (Reference Sen and Anantaraman1971) is 810–1400 μm in length, similar to, or even smaller than that of Physocephalus sp. (1417–1635 μm; this study) or P. sexalatus (940–1728 μm; Seurat, Reference Seurat1915). In addition, S. lupi larvae reported by Seurat (Reference Seurat1915), Chhabra, Reference Chhabra1968 and those found in this study have a distance from the nerve ring to the anterior end of the body shorter than that from the excretory pore, whereas, it is converse in S. lupi larvae reported by Sen & Anantaraman (Reference Sen and Anantaraman1971) and by Mohtasebi et al. (Reference Mohtasebi, Teimouri, Abbaszadeh Afshar, Mobedi, Abbasian, Totonchian and Mowlavi2021). These overlaps and differences in morphometric data suggest that misidentification may occur. This potential problem can be avoided by careful observation of their detailed morphological features under a light microscope at magnification above 200×. In the anterior end, S. lupi larva has a short buccal cavity in comparison to the muscular oesophagus (about one-third) while Physocephalus sp. larva has a much longer buccal cavity, almost equal to the length of the muscular oesophagus. The nerve ring and excretory pore of S. lupi are located at the base of the muscular oesophagus, while those of Physocephalus sp. are located at about the middle of the muscular oesophagus. In the posterior end, the tail of S. lupi larva looks like a cup bottom carrying a rosette of closely cuticular spines at the end, while the posterior end of Physocephalus sp. is a characteristic small knob, about 7 to 8 μm long, bearing small cuticular spines around the knob tail. These characteristics of the second larval type are similar to P. sexalatus larva described by Seurat (Reference Seurat1915). However, morphology alone might be insufficient to identify species of nematode larvae because larvae of different genera share common characteristics (Bain et al., Reference Bain, Mutafchiev, Junker and Schmidt-Rhaesa2014) and those of more than one spirurid species can be found in the same dung beetle species (Seurat, Reference Seurat1915; Alicata, Reference Alicata1935). Unfortunately, molecular data do not help in identifying this larva to species level due to the lack of completely identical sequences in GenBank, including the sequence of P. sexalatus to which the larva is morphologically similar. These shortages of necessary data point out that morphological descriptions and molecular data of more spirurid larvae should be investigated and deposited in a database for use in future studies. For morphology, detailed morphological features of the anterior and posterior ends of the body are important and need to be carefully examined.
The available data from studies on S. lupi larvae in beetle hosts showed that a wide variety of dung beetles serve as intermediate hosts of S. lupi and vary from place to place. They are: Scarabaeus sacer, Scarabaeus variolosus and Gymnopleurus sturmi in the United States (Bailey et al., Reference Bailey, Cabrera and Diamond1963); Copris hispana in Algeria (Seurat, Reference Seurat1915); Onthophagus pugionatus, Onthophagus sugillatus, Onthophagus ebenus, Onthophagus obtusicornis, Gymnopleurus virens, Gymnopleurus humanus, Pachylomerus femoralis, Scarabaeus rugosus, Kheper nigroaeneus, Anachalcos convexus and also a millipede species, Daratoagonus crristulatus (experimental infection) in South Africa (Du Toit et al., Reference Du Toit, Scholtz and Hyman2008; Mukaratirwa et al., Reference Mukaratirwa, Pillay and Munsammy2010); Onthophagus sellatus in Israel (Gottlieb et al., Reference Gottlieb, Markovics, Klement, Naor, Samish, Aroch and Lavy2011); Scarabaeus armeniacus in Iran (Mohtasebi et al., Reference Mohtasebi, Teimouri, Abbaszadeh Afshar, Mobedi, Abbasian, Totonchian and Mowlavi2021); Gymnopleurus koenigi, Oniticellus pallens, Oniticellus pallipes, Onthophagus deflexicollis, Onthophagus quadridentatus, Onthophagus bonasus, Onthophagus dama, Onthophagus gazella, Onthophagus mopsus, Onitis philemon, Catharsius pithecius, Euoniticellus pallipes, Scarites indus, Hybosorus orientalis, Hister maindronii and Hister lutarius in India (Anantaraman & Jayalakshmi, Reference Anantaraman and Jayalakshmi1963; Chhabra, Reference Chhabra1968; Chowdhury & Pande, Reference Chowdhury and Pande1969); and Gymnopleurus sinnatus, Gymnopleurus mopsus, Scarabaeus sacer and Cathon spp. in China (Faust, Reference Faust1928; Ono, Reference Ono1929), but Cathon beetles reported by Faust (Reference Faust1928) were suggested to belong to the genus Paragymnopleurus (Theodorides, Reference Theodorides1952). The detection of S. lupi larvae from two beetle species of C. molossus and C. szechouanicus in Vietnam in the present study seems to be the first report from south-east Asia, adding the two species as new intermediate hosts of S. lupi. Since dung beetles are widespread and abundant in tropical Asian regions, particularly in south-east Asia, more surveys in different countries of this region should be conducted to fully understand the diversity of intermediate hosts of S. lupi and other spirurid nematodes.
Concerning prevalence, previous studies showed that infection rates of S. lupi vary from place to place, and from beetle species to species. In India, Chowdhury & Pande (Reference Chowdhury and Pande1969) reported the prevalence of S. lupi larvae in four beetle species, Onthophagus deflexicollis, Onthophagus quadridentatus, Oniticellus pallens and Oniticellus pallipes, these at the same study site were 3.8%, 9.9%, 15%, and 31.8%, respectively, and burdens ranged from 1–24 larvae per beetle. In a study in Iran, Mohtasebi et al. (Reference Mohtasebi, Teimouri, Abbaszadeh Afshar, Mobedi, Abbasian, Totonchian and Mowlavi2021) found an infection rate of 1.5% (3/200) in Scarabaeus armeniacus with an intensity of 1–6 larvae per beetle. A survey in South Africa revealed considerable variation of infection rates in different study sites: 13.5% (7/52) and burdens of 1–129 larvae per beetle in the urban area, 2.3% (3/129) and intensity of 1–10 larvae per beetle in the rural area, while beetles collected in the suburban areas were not infected (Du Toit et al., Reference Du Toit, Scholtz and Hyman2008). In the present study, the prevalence and intensities of S. lupi larvae in two beetle hosts are in these ranges. In addition, the larvae of Physocephalus sp. were found to be concurrently present in both host species with a prevalence slightly higher than that of S. lupi. The genus Physocephalus includes nine species parasitizing suids, equids, camelids, cattle, lagomorphs and rodents (Bain et al., Reference Bain, Mutafchiev, Junker and Schmidt-Rhaesa2014). Physocephalus sp. larva found in this study was similar to P. sexalatus larva – a parasite of pigs – described by Seurat (Reference Seurat1915). The finding of this larva indicated the possibility of Physocephalus infection in pigs in the study site, suggesting that morphological and molecular studies on adult nematodes from pigs are necessary to confirm the species identification and that studies on S. lupi larvae in beetle hosts should also pay attention to other spirurid larvae.
Conclusion
The present study found S. lupi larvae from two beetle species of C. molossus and C. szechouanicus in Vietnam, adding them as new intermediate hosts of S. lupi. In addition, larvae of an unidentified spirurid nematode, Physocephalus sp., was concurrently found in both dung beetle species. Although these larvae are morphologically different from each other in detailed morphological features of the anterior and posterior ends, it is difficult to distinguish them at a low magnification due to their minute size and overlap measurements. To avoid misidentification of spirurid larvae from dung beetles, detailed taxonomic characteristics of larvae should be carefully examined.
Acknowledgement
We thank Dr Pham Van Phu for his help in identification of dung beetles.
Financial support
This study was supported by Vietnam Academy of Science and Technology under code number NVCC09.06/22–23.
Conflicts of interest
None.






