



Introduction
The Guadalupian–Lopingian boundary (GLB) biotic crisis has been regarded as one of the least understood crisis events in the Phanerozoic era (Raup and Sepkoski, Reference Raup and Sepkoski1982; Bambach, Reference Bambach2002; Jost et al., Reference Jost, Mundil, He, Brown, Altiner, Sun, DePaolo and Payne2014). There remain debates about whether it was a sudden extinction (Stanley and Yang, Reference Stanley and Yang1994; Isozaki and Servais, Reference Isozaki and Servais2018; Arefifard and Payne, Reference Arefifard and Payne2020; Rampino and Shen, Reference Rampino and Shen2020) or a gradual diversity reduction (Clapham et al., Reference Clapham, Shen and Bottjer2009; Fan et al., Reference Fan2020; Shen et al., Reference Shen2020; Lee et al., Reference Lee, Shim, Nakrem, Woo and Tazawa2022), which may be related to the varied crisis times over different areas and selective extinctions among different marine organisms. During the GLB crisis, brachiopods were severely affected, and 30% of genera and 87% of species became extinct (Shen and Shi, Reference Shen and Shi1996; Sun and Shen, Reference Sun, Shen, Rong and Fang2004), but this crisis was restricted to the genus and species level (Shen et al., Reference Shen, Zhang, Li, Mu and Xie2006). Following this, the brachiopod fauna had relatively low diversity in the initial early Wuchiapingian recovery and then rapidly radiated in the late Wuchiapingian (Chen et al., Reference Chen, Campi, Shi and Kaiho2005; Shen et al., Reference Shen, Zhang, Li, Mu and Xie2006; Shen and Zhang, Reference Shen and Zhang2008; Shen and Shi, Reference Shen and Shi2009; Fan et al., Reference Fan2020). During the GLB crisis, the dominant Guadalupian brachiopod genera, Neoplicatifera, Urushtenoidea, and Vediproductus, were replaced by Permophricodothyris, Haydenella. and Edriosteges in the Wuchiapingian (Shen et al., Reference Shen2019). However, due to the widespread absence of marine strata related to the global regression event (Shen et al., Reference Shen2020), brachiopod fossil data in the GLB interval remain deficient (Shen and Zhang, Reference Shen and Zhang2008; Shen and Clapham, Reference Shen and Clapham2009; Ghaderi et al., Reference Ghaderi, Garbelli, Angiolini, Ashouri, Korn, Rettori and Gharaie2014; Tazawa et al., Reference Tazawa, Kaneko, Suzuki and Hasegawa2015; Xu et al., Reference Xu, Cao, Yuan, Zhang and Shen2018). More robust, section-based studies on GLB brachiopod faunas are required to investigate the crisis and recovery/radiation patterns.
In the 1980s, numerous works gave overviews of the Lopingian brachiopod faunas from both the Yangtze block and southeastern China (Zhan in Hou et al., Reference Hou, Zhan and Chen1979; Liao, Reference Liao1980; Wang et al., Reference Wang, Liu, Jin, Hu, Liang and Liao1982; Yang, Reference Yang, Feng, Xu, Lin and Yang1984; Liao in Yang et al., Reference Yang, Yin, Wu, Yang, Ding and Xu1987; Hu, Reference Hu1989; Li et al., Reference Li, Zhan, Dai, Jin, Zhu, Zhang, Huang, Xu, Yan and Li1989). Later, several more-extensive systematic works for Wuchiapingian brachiopods from South China were published (Chen et al., Reference Chen, Campi, Shi and Kaiho2005; Li and Shen, Reference Li and Shen2008; Shen and Zhang, Reference Shen and Zhang2008; Shen and Shi, Reference Shen and Shi2009), most of which focused on brachiopod fauna from the Yangtze block. The present paper provides a detailed description of a brachiopod fauna from the Liannan area of Guangdong Province, southeastern China, and discusses the faunal changes across different environments in the Wuchiapingian and the implications for brachiopod faunal recovery/radiation after the GLB crisis.
Geological setting and age
The described specimens were collected from the Shuizhutang Formation at the Liannan section, which is located about 10 km southeast of Liannan County in Qingyuan, northern Guangdong Province (Fig. 1). In the Lopingian (late Permian), the studied area recorded two regressive–transgressive cycles, represented by the Wuchiapingian Longtan Formation and Shuizhutang Formation (from coal-bearing deposits to carbonates) and the Changhsingian Wangpanli Formation and Changxing Formation (from coal-bearing deposits to carbonates), respectively (Hou et al., Reference Hou, Zhan and Chen1979). The Shuizhutang Formation represented in the Liannan section was deposited in a conjunction area between the shallow-water restricted carbonate platform (Lianshan–Yangshan area) and the clastic shelf, which is close to the Yunkai old land (Fig. 2). The Shuizhutang Formation comprises mainly dark grey siliceous mudstone in the lower part and yellowish siliceous and calcareous mudstone in the middle to upper part (Fig. 3).

Figure 1. Location of the Liannan section, northern Guangdong Province, southeastern China.

Figure 2. Paleogeographical map of South China during Wuchiapingian (from Feng et al., Reference Feng, Yang, Jin, Li and Bao1997), showing the location of the Liannan section as well as the sites of the Yansha, Xiaoyuanchong, and Daijiagou sections also referred to in this paper.

Figure 3. Distribution of brachiopods from the Shuizhutang Formation at the Liannan section.
Judging from the yielded fusulinids Codonofusiella kwangsiana Sheng, Reference Sheng1963 and Gallowayinella meitienensis Chen, Reference Chen1934 and ammonoids Prototoceras, Araxoceras, and Konglingites (Hou et al., Reference Hou, Zhan and Chen1979), the age of the Shuizhutang Formation should be assigned to the late Wuchiapingian (Shen and Zhang, Reference Shen and Zhang2008; Shen, Reference Shen2018). In addition, the Liannan brachiopod fauna, which is dominated mainly by Fusichonetes, Neochonetes, Transennatia, Cathaysia, and Orthothetina, should belong to the Transennatia–Orthothetina Assemblage and also indicates the late Wuchiapingian age (Yang et al., Reference Yang, Yin, Wu, Yang, Ding and Xu1987).
Materials and methods
In total, 1,900 complete brachiopod specimens were collected from the upper Wuchiapingian of the Liannan section. The Liannan fauna contains 57 brachiopod species in 28 genera and is summarized in Figure 3.
To verify the distinctness of the spondylium development of A. minor n. sp., spondylium length and width and total shell length and width of all Araxathyris specimens from the Liannan section were measured. To highlight the large variation in width/length ratios of Cathaysia chonetoides (Chao, Reference Chao1927), shell width and length of all Cathaysia specimens from the Liannan section were also measured. Further, to present the shell outline and size of the subgenus Neochonetes (Zhongyingia), we measured shell width and length of all specimens of this subgenus from the Liannan section and Lopingian specimens in the literature. Both the shell size and spondylium size were approximated with the geometric mean of length and width, following Jablonski (Reference Jablonski, Jablonski, Erwin and Lipps1996). To show the similarity of brachiopod faunas from selected sections, absence–presence data at genus level were collected, and cluster analysis using the unweighted paired group algorithm and the Raup–Crick index were adopted. All analyses were carried out using the software PAST (Hammer et al., Reference Hammer, Harper and Ryan2001).
Repository and institutional abbreviation
Types and other specimens examined in this study are deposited in the Faculty of Earth Science and Resources, China University of Geosciences (Beijing) (CUGB), China, with the prefix LN for the Liannan section in Guangdong Province, South China.
Results and discussion
Comparison of the Liannan fauna with other contemporaneous faunas from South China
There have been only several Wuchiapingian brachiopod faunas reported from South China, including faunas from the Yansha and Xiaoyuanchong sections (Shen and Zhang, Reference Shen and Zhang2008) and the Daijiagou section (Chen et al., Reference Chen, Campi, Shi and Kaiho2005) (Fig. 2). In addition, there are Wuchiapingian brachiopod faunas described by Shen and Zhang (Reference Shen and Zhang2008) and Chen et al. (Reference Chen, Campi, Shi and Kaiho2005) from the Yanhu, Sanhe, Yejiwo, and Chuanmu sections. Since these latter four sections all yielded fewer than five species, they are not included herein.
Compared with the three aforementioned sections, the Liannan fauna shows much higher α diversity. There are 57 species in the Liannan section, 11 species from the Xiaoyuanchong section, 34 species from the Yansha section, and 16 species from the Daijiagou section. According to the result of cluster analysis, the Liannan fauna shows greatest similarity to the Xiaoyuanchong and Yansha faunas and least similarity to the Daijiagou fauna (Fig. 4). Some common genera in the Lopingian of South China, including Spinomarginifera, Neochonetes, Transennatia, Tyloplecta, and Haydenella, all occurred in the Liannan, Yansha, and Xiaoyuanchong sections (Fig. 5). Although Spinomarginifera and Transennatia also occurred in the Daijiagou section, genera that only occasionally occurred in the Yangtze and other blocks, such as Magniderbyia and Niutoushania, existed only in the Daijiagou fauna.

Figure 4. Q-mode cluster analysis of species abundance data using unweighted paired group algorithm and the Raup–Crick index.
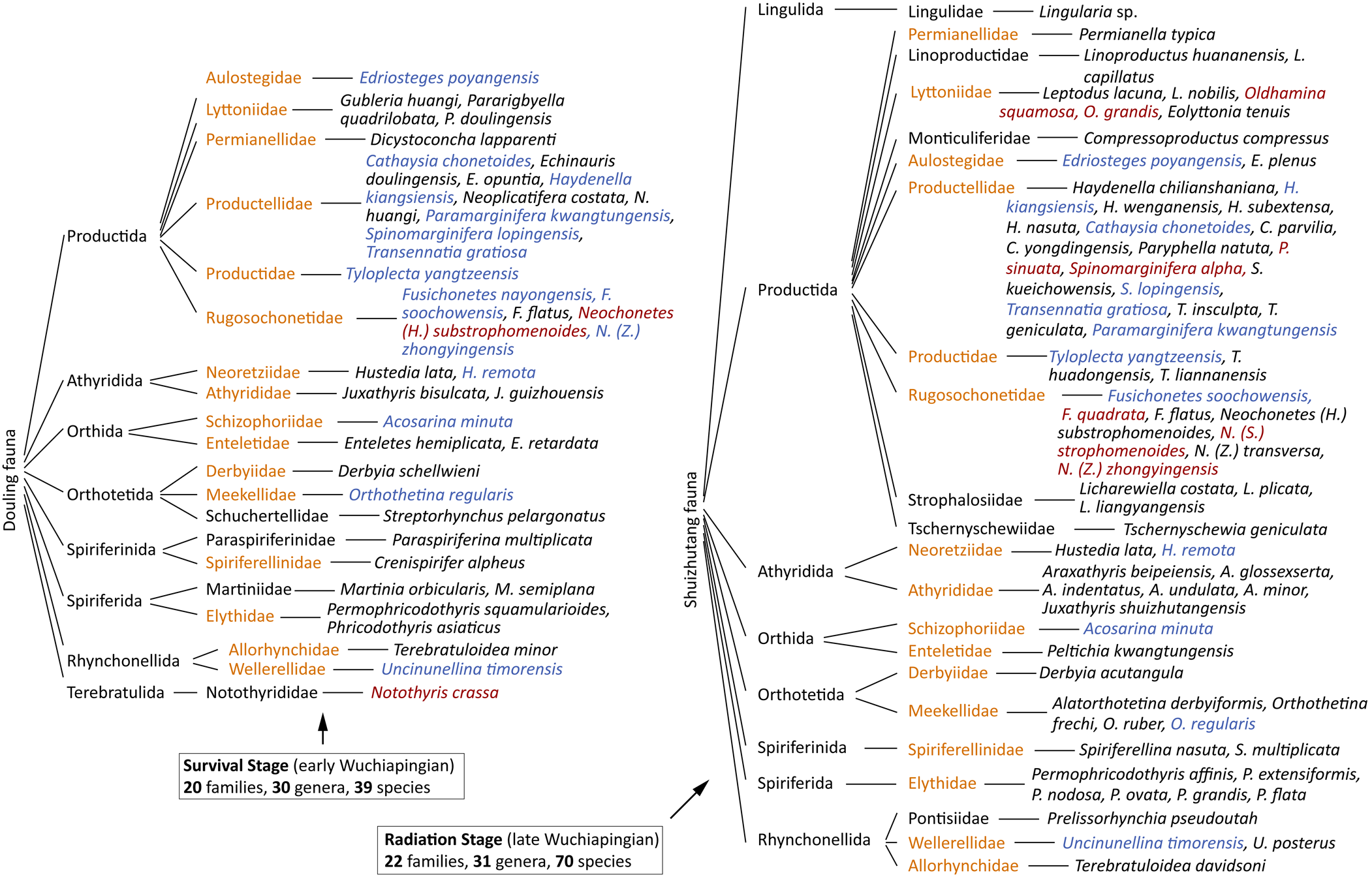
Figure 5. Faunal composition of the Douling fauna and the Shuizhutang fauna. Yellow words represent their shared families. Blue-gray words represent their shared species. Red words represent Changhsingian key taxa.
The similarity of the faunal composition but difference in α diversity among the four studied sections can be explained by their paleogeographical positions during the Wuchiapingian. Although they are all located in or close to the shallow-water clastic shelf, the Liannan, Yansha, and Xiaoyuanchong sections are all situated in southeastern China while the Daijiagou section is in the northwestern Yangtze block; thus, the former three faunas show greater similarity. A similar pattern is also seen in Changhsingian brachiopods from South China. The Sanhe and Dapaichong faunas (which are from southeastern China) are distinctly differentiated from the Daijiagou, Beifengjing, Yutianbao, Nantong, and Tuanxi faunas (which are from the Upper Yangtze) in the network diagram (Wu et al., Reference Wu, Zhang, Stubbs and Sun2020, fig. 4).
Implications for brachiopod faunal recovery/radiation from the GLB crisis
After the GLB crisis, the brachiopods maintained a relatively low diversity in the early Wuchiapingian (survival stage) and then rapidly radiated in the late Wuchiapingian (radiation stage) in South China (Sun and Shen, Reference Sun, Shen, Rong and Fang2004). During this transitional interval, southern Hunan Province–northern Guangdong Province serves as one of the key areas (Fig. 2), yielding unusually diverse brachiopod faunas (Shen and Zhang, Reference Shen and Zhang2008; this study).
In southern Hunan Province, an earliest Wuchiapingian brachiopod fauna was reported from the upper part of Douling Formation (which contains mainly calcareous mudstone interbedded with thin-bedded bioclastic limestone, deposited in the continental shelf sedimentary environment; see Shen and Zhang, Reference Shen and Zhang2008 for details) at the Yanshan, Xiaoyuanchong, Douling, Yanhu, Yejiwo, and Sanhe sections (Wang and Jin, Reference Wang and Jin1991; Shen and Archbold, Reference Shen and Archbold2002; Shen and Zhang, Reference Shen and Zhang2008), which sheds light on the survival stage just after the GLB crisis. The Douling brachiopod fauna includes 39 species in 30 genera and 20 families (without undetermined species), which consist mainly of the Wuchiapingian taxa (e.g., Acosarina minuta (Abich, Reference Abich1878), Transennatia gratiosa (Waagen, 1884), Edriosteges poyangensis (Kayser, Reference Kayser and von Richthofen1883), Haydenella kiangsiensis (Kayser, Reference Kayser and von Richthofen1883), Tyloplecta yangtzeensis (Chao, Reference Chao1927); see Fig. 5 for details). However, this fauna still has relatively low diversity and contains only a few key Changhsingian brachiopod taxa (e.g., Neochonetes (Huangichonetes) substrophomenoides Shen and Archbold, Reference Shen and Archbold2002 and Notothyris crassa Reed, Reference Reed1944; see Fig. 5).
In the northern Guangdong Province, late Wuchiapingian brachiopods were reported from the Shuizhutang Formation at Liannan, Lianshan, and Yangshan areas (Hou et al., Reference Hou, Zhan and Chen1979; this study). Compared with faunas from the Douling Formation, this fauna presents a much higher diversity (70 species in 31 genera and 22 families, without undetermined species), which shows the rapid recovery/radiation stage of brachiopods in the late Wuchiapingian. This fauna also presents many more key Changhsingian taxa (e.g., Oldhamina squamosa Huang, Reference Huang1932, Paryphella sinuata (Zhan in Hou et al., Reference Hou, Zhan and Chen1979), Spinomarginifera alpha Huang, Reference Huang1932; see Fig. 5 for details).
It appears, therefore, that brachiopod faunas from the Douling Formation and Shuizhutang Formation, respectively, present the survival stage (early Wuchiapingian) and recovery/radiation stage (late Wuchiapingian) after the GLB major crisis. From comparison of these two faunas, it is clear that they share most brachiopod families, implying that the recovery/radiation occurred mainly in the genus-/species-level richness, which is consistent with the GLB brachiopod crisis pattern (Sun and Shen, Reference Sun, Shen, Rong and Fang2004). The similar diversity of genera and species in the Douling fauna prove that, during the survival stage, the brachiopod initial recovery occurred mainly at the genus level. Finally, the highly diversified Shuizhutang fauna indicates that rapid radiation occurred at both genus and species levels, and the presence of more key Changhsingian taxa highlights the significant influence on the Changhsingian brachiopod fauna in South China.
Systematic paleontology
The classification above genus level adopted herein follows Kaesler (Reference Kaesler2000a, Reference Kaeslerb, Reference Kaesler2002, Reference Kaesler2006) and Selden (Reference Selden2007). Herein, species that have been fully described in recent papers are only listed in Table 1 with illustrations (mainly Figs. 6—8 and a few specimens in the other figures), and their detailed descriptions are shown in the Supplementary text.

Figure 6. Brachiopods from the Shuizhutang Formation. (1) Lingularia sp., a ventral valve, LN191001. (2–9, 26, 27) Fusichonetes soochowensis (Chao, Reference Chao1928): (2–6, 9) external molds of dorsal valves: (2) LN221803, (3) LN223301, (4) LN223402, (5) LN223601, (6) LN216601, (9) LN221701; (7, 8, 26, 27) internal molds of ventral valves: (7) LN001801, (8) LN221301, (26) LN001401, (27) LN001701. (10–14) F. quadrata (Zhan in Hou et al., Reference Hou, Zhan and Chen1979): (10–12, 14) external molds of dorsal valves: (10) LN188903, (11) LN222301, (12) LN222703, (14) LN223703; (13) an internal mold of a ventral valve, LN222802. (15–17) F. flatus (Shen and Archbold, Reference Shen and Archbold2002), external molds of dorsal valves: (15) LN002603, (16) LN221102, (17) LN222003. (18–21) Neochonetes (Huangichonetes) substrophomenoides: (18–20) internal molds of ventral valves: (18) LN001201, (19) LN002403, (20) LN216503; (21) dorsal interior, LN220503. (22–25) N. (Sommeriella) strophomenoides Waagen, 1884: (22–24) internal molds of ventral valves: (22) LN003403, (23) LN218003, (24) LN219901; (25) an internal mold of a dorsal valve, LN220303. Scale bars = 3 mm.

Figure 7. Brachiopods from the Shuizhutang Formation. (1–6) N. (Zhongyingia) transversa Zhang et al., Reference Zhang, He, Shi, Zhang and Wu2015: (1, 3–5) internal molds of ventral valves: (1) LN217101, (3) LN000501, (4) LN002001, (5) LN217801; (2) an external mold of a dorsal valve, LN216301; (6) an internal mold of a dorsal valve, LN217401. (7–10) N. (Z.) zhongyingensis Liao, Reference Liao1980: (7, 8) external molds of dorsal valves: (7) LN217703, (8) LN218301; (9) an internal mold of a ventral valve, LN218501; (10) an internal mold of a dorsal valve, LN218601. (11–15) Cathaysia chonetoides: (11, 12, 14, 15) internal molds of ventral valves: (11) LN200402, (12) LN183703, (14) LN197803, (15) LN198403; (13) an external mold of a dorsal valve, LN195502. (16–19) C. parvulia Chang in Yang, Reference Yang, Ni, Chang and Zhao1977, internal molds of ventral valves: (16) LN002503, (17) LN195102, (18) LN196703, (19) LN197102. (20–22) C. yongdingensis Liao in Wang et al., Reference Wang, Liu, Jin, Hu, Liang and Liao1982: (20, 22) external molds of dorsal valves: (20) LN196103, (22) LN200503; (21) an internal mold of a ventral valve, LN196903. (23, 24) Paryphella nasuta Liao, Reference Liao1984, internal molds of ventral valves: (23) LN209201, (24) LN209601. Scale bars = 3 mm.

Figure 8. Brachiopods from the Shuizhutang Formation. (1–4) Acosarina minuta: (1, 3) internal molds of dorsal valves: (1) LN242203, (3) LN243302; (2, 4) internal molds of ventral valves: (2) LN243002, (4) LN243802. (5, 6) Hustedia lata Grabau, Reference Grabau1931: (5) an external mold of a dorsal valve, LN191502, (6) an external mold of a ventral valve, LN191702. (7–11) H. remota (Eichwald, Reference Eichwald1860): (7, 8) internal molds of dorsal valves: (7) LN001902, (8) LN000803; (9–11) internal molds of ventral valves: (9) LN001003, (10) LN192503, (11) LN212203. (12–18) Prelissorhynchia pseudoutah (Huang, Reference Huang1933): (12, 14) internal molds of ventral valves: (12) LN000602, (14) LN001903; (13, 18) internal molds of dorsal valves: (13) LN000702, (18) LN191101; (15–17) an inner core of a conjoined shell, LN192702: (15) ventral view, (16) dorsal view, (17) anterior view. (19) Uncinunellina postera (Waagen, 1883), an internal mold of a dorsal valve, LN177501. (20, 21) Uncinunellina sp., ventral valves: (20) LN186603, (21) LN188801. (22–24) Araxathyris beipeiensis Xu and Grant, Reference Xu and Grant1994, internal molds of ventral valves: (22) LN169903, (23) LN176104, (24) LN187802. Scale bars = 5 mm.
Table 1. List of species with only illustrations in the text.


Figure 9. Brachiopods from the Shuizhutang Formation. (1) Haydenella chilianshaniana, a ventral valve, LN179103. (2–11) H. kiangsiensis: (2, 10, 11) internal molds of ventral valves: (2) LN184303, (10) LN211103, (11) LN172003; (3–7, 9) external molds of dorsal valves: (3) LN202903, (4) LN204702, (5) LN207103, (6) LN207302, (7) LN208103, (9) LN208603; (8) lateral view of (7). (12) H. subextensa, an internal mold of a ventral valve, LN201403. (13–17) H. nasuta: (13–15, 17) external molds of dorsal valves: (13) LN201303, (14) LN202003, (15) LN204303, (17) LN208801; (16) an internal mold of a ventral valve, LN207603. (18–20) Spinomarginifera kueichowensis Huang, Reference Huang1932, an internal mold of a conjoined shell, LN268503. Scale bars = 3 mm.

Figure 10. Brachiopods from the Shuizhutang Formation. (1–3) Spinomarginifera kueichowensis: (1) an internal mold of a ventral valve, LN186803; (2) an internal mold of a dorsal valve, LN213702; (3) an external mold of a dorsal valve, LN214800. (4–8) S. sp.: (4, 7) external molds of ventral valves: (4) LN213401, (7) LN214901; (5) an internal mold of a ventral valve, LN214401; (6) dorsal interior, LN214600; (8) an external mold of a dorsal valve, LN215001. (9–20) Transennatia gratiosa: (9, 11, 18) internal molds of ventral valves: (9) LN182901, (11) LN215901, (18) LN234801; (10, 13) posterior views of (11); (12) lateral view of (11); (14, 15, 17, 19, 20) external molds of dorsal valves: (14) LN227301, (15) LN233001, (17) LN233201, (19) LN234903, (20) LN235403; (16) an internal mold of a dorsal valve, LN233101. (21–24) T. insculpta: (21) ventral valve, LN205201; (22–24) external molds of dorsal valves: (22) LN224403, (23) LN224901, (24) LN225203. Scale bars = 3 mm.
Order Productida Sarytcheva and Sokolskaja, Reference Sarytcheva and Sokolskaya1959
Suborder Productidina Waagen, Reference Waagen1883
Superfamily Productoidea Gray, Reference Gray1840
Family Productellidae Schuchert, Reference Schuchert and Pompeckj1929
Subfamily Productininae Muir-Wood and Cooper, Reference Muir-Wood and Cooper1960
Tribe Chonetellini Licharew in Sarytcheva et al., Reference Sarytcheva, Licharew, Sokolskaja and Orlov1960
Genus Haydenella Reed, Reference Reed1944
Type species
Productus kiangsiensis Kayser, Reference Kayser and von Richthofen1883 from upper Carboniferous of Loping, Jiangxi, China.
Remarks
On the basis of the Productus kiangsiensis Kayser, Reference Kayser and von Richthofen1883 collected from the Productus Limestones of the Salt Range as the type species, Reed (Reference Reed1944) proposed the genus Haydenella. However, there have been debates concerning the affiliation of type species before and after the nomenclature of Haydenella, and the classification position of this genus has remained confused for over 100 years. In this study, we follow the classification system of Brachiopoda in the Treatise (Brunton et al., Reference Brunton, Lazarev, Grant, Jin and Williams2000, p. 428).
By having a nearly globose ventral valve, Argentiproductus Cooper and Muir-Wood, Reference Cooper and Muir-Wood1951 is similar to this genus but differs in the presence of lamellae and absence of rugae on ears. Ogbinia Sarytcheva in Ruzhentsev and Sarytcheva, Reference Ruzhentsev and Sarytcheva1965 resembles Haydenella in the low ribs and short dorsal medium septum, but the former has an elongate outline and no rugae on the ears. Avonia Thomas, Reference Thomas1914 is similar to the present genus in the concavoconvex profile, weak ribs, and short dorsal medium septum, but the former possesses weak lamellae and bifid cardinal processes. In the ribs on shell surface and rugae on ears and weakly marked brachial ridges, Linoproductus Chao, Reference Chao1927 is very similar to Haydenella, but the former has an elongate subcircular outline and maximum shell width at the hingeline.
Haydenella chilianshaniana Jin and Ye, Reference Jin and Ye1979
Figure 9.1
- 1979
Haydenella chilianshaniana Jin and Ye, p. 84, pl. 24, figs. 7–12.
Holotype
42631, Bayinhe Group, Tianjun County, Qinghai, China (Jin and Ye, Reference Jin and Ye1979, pl. 24, figs. 11, 12).
Occurrence
Permian; China.
Description
Medium size for genus, semicircular outline; cardinal extremities blunt, with a cardinal angle of about 100°; hingeline slightly shorter than the maximum width. Ventral valve moderately convex, with maximum convexity posteriorly; ears small, triangular, and well demarcated from visceral region; visceral disk rounded; sulcus absent. Ornamentation of costellae, absent at ears; several indistinct rugae on ears arranged perpendicularly to hinge.
Material
A ventral valve (LN179103).
Remarks
The lack of a sulcus in the ventral valve makes the present species different from many of its counterparts in the genus. Haydenella bofengensis Zhu, Reference Zhu1990 resembles the present species in the lack of sulcus, convexity of ventral valve, and semicircular outline but differs in having a long trail. Haydenella minuta Sarytcheva in Ruzhentsev and Sarytcheva, Reference Ruzhentsev and Sarytcheva1965 also has no sulcus but differs in possessing a more pointed and less inflated umbo.
Haydenella kiangsiensis (Kayser, Reference Kayser and von Richthofen1883)
Figure 9.2–9.11
- Reference Kayser and von Richthofen1883
Productus kiangsiensis Kayser, p. 185, pl. 26, figs. 6–11.
- Reference Chao1927
Avonia kiangsiensis; Chao, p. 125, pl. 14, figs. 14–16.
- Reference Chao1928
Thomasia kiangsiensis; Chao, p. 50, pl. 6, fig. 18.
- Reference Huang1932
Linoproductus kiangsiensis; Huang, p. 46, pl. 3, figs. 13–19.
- Reference Reed1944
Productus (Haydenella) kiangsiensis; Reed, p. 78, pl. 19, fig. 2.
- Reference Muir-Wood and Cooper1960
Haydenella kiangsiensis; Muir-Wood and Cooper, pl. 65, figs. 1–14.
- Reference Zhang and Jin1961
Argentiproductus kiangsiensis; Zhang and Jin, p. 411, pl. 3, figs. 13, 14.
- Reference Jin and Hu1978
Haydenella kiangsiensis; Jin and Hu, pl. 2, fig. 25.
- Reference Jin and Ye1979
Haydenella kiangsiensis; Zhan in Hou et al., p. 81, pl. 5, figs. 3, 4.
- Reference Zeng, He and Zhu1995
Haydenella kiangsiensis; Zeng et al., pl. 5, fig. 8.
- Reference Campi and Shi2005
Haydenella kiangsiensis; Campi and Shi, p. 111, figs. Z, bb, cc, ee.
- Reference He, Shi, Bu and Niu2008
Haydenella kiangsiensis; He et al., p. 815, fig. 4.17–4.19.
- Reference Li and Shen2008
Haydenella kiangsiensis; Li and Shen, p. 311, fig. 4(7).
- Reference Shen and Shi2009
Haydenella kiangsiensis; Shen and Clapham, p. 721, pl. 1, fig. 28, pl. 2, fig. 1.
- Reference Crippa and Angiolini2012
Haydenella kiangsiensis; Crippa and Angiolini, p. 138, fig. 11c–j.
- Reference Ghaderi, Garbelli, Angiolini, Ashouri, Korn, Rettori and Gharaie2014
Haydenella kiangsiensis; Ghaderi et al., p. 35, pl. 1, figs. 1, 2.
Holotype
Unknown. Kayser (Reference Kayser and von Richthofen1883) did not designate a holotype for this species.
Occurrence
Permian to the earliest Triassic; Armenia, Azerbaijan, Cambodia, China, Greece, Iran, Japan, Malaysia, Pakistan, the Russian Federation, Tajikistan.
Description
Medium size for genus, outline semicircular to suboval; cardinal extremities blunt; hingeline shorter than the maximum width; maximum width at middle to posterior part. Ventral valve moderately convex, with maximum convexity at posterior part, becoming flattened anteriorly; sulcus absent; costellae low and coarse, bifurcated from midvalve in some specimens and originating from anterior of beak; spines in row between ears and corpus and scattered on the valve; interior with fine and radially distributed papillae, mostly distinct only at posterior part. Dorsal valve moderately concave, with maximum concavity at middle to anterior part; fold absent; ears small, ornamented by regularly distributed and distinct rugae; costellae bifurcated at posterior part in some specimens; cardinal process quadrivalve.
Material
Six external molds of dorsal valves (LN202903, LN204702, LN207103, LN207302, LN208103, LN208603) and three internal molds of ventral valves (LN184303, LN211103, LN172003).
Remarks
H. kiangsiensis is like H. bofengensis in that both have semicircular outline and no sulcus, but the latter has a more convex ventral valve and long trail. The current species differs from H. chilianshaniana by its more rounded outline and more pointed ventral umbo. It resembles Haydenella granti Yanagida and Nakornsri, Reference Yanagida and Nakornsri1999 in rounded outline and pointed umbo, but the latter has relatively more transverse outline and more-developed concentric growth lines.
Haydenella nasuta Zeng, Reference Zeng1993
Figure 9.13–9.17
- Reference Zeng1993
Haydenella nasuta Zeng, p. 751, pl. 1, figs. 4, 5.
Holotype
MPUM 10770, Ruteh Limestone, North Iran (Crippa and Angiolini, Reference Crippa and Angiolini2012, fig. 11l).
Occurrence
Guadalupian to Lopingian; China, Iran.
Description
Small to medium for genus; transversely to slightly elongated subrounded; hingeline slightly shorter than the maximum width; cardinal extremities right-angled to blunt. Ventral valve moderately convex with maximum convexity at umbonal region; flanks gently inclined; nasute protrusion short; sulcus not observed or very weakly developed; ears small, distinct from corpus with a row of spine between them. External surface ornamented by low costellae originating from slightly anterior to umbo; rugae very strong on ears, about 2–3 on each ear, disposed nearly at about right angle to hingeline; erect body spines sporadically scattered on surface. Internal surface with fine and radially distributed papillae.
Dorsal valve slightly to moderately concave, with maximum concavity at middle to anterior part; umbo normally not or slightly over hingeline; ears small; a long and wide nasute protrusion developed, beginning from anterior part of midvalve; fold not observed. External surface with coarse costellae originating from anterior part of umbo, bifurcated into several much finer costellae from midvalve in some specimens; 3–4 pairs of rugae developed on ears.
Material
Four external molds of dorsal valves (LN201303, LN202003, LN204303, LN208801) and an internal mold of a ventral valve (LN207603).
Remarks
This species is most similar to Haydenella eminens Crippa and Angiolini, Reference Crippa and Angiolini2012 in both having a prominent nasute protrusion, but the latter has much finer costellae.
Haydenella subextensa Zhan in Hou et al., Reference Hou, Zhan and Chen1979
Figure 9.12
- Reference Hou, Zhan and Chen1979
Haydenella subextensa Zhan in Hou et al., p. 82, pl. 11, figs. 9–13.
- Reference Zeng, He and Zhu1995
Haydenella subextensa; Zeng et al., pl. 6, fig. 5.
- Reference Wu, Zhang, Stubbs, Liu and Sun2021
Haydenella? sp.; Wu et al., p. 874, fig. 10S–V.
Holotype
K0125, Wangpanli Formation, Guangdong Province, China (Hou et al., Reference Hou, Zhan and Chen1979, pl. 11, fig. 11).
Occurrence
Lopingian; China.
Description
Shell medium size for genus, transversely suboval in outline; hingeline shorter than shell maximum width. Ventral valve moderately convex, somewhat flattened transversely; umbo pointed and slightly over hingeline; ears small and ornamented with rugae. External surface ornamented with faint costellae; body spine sporadically scattered on visceral region. Internal surface with fine papillae radially distributed on visceral region.
Material
An internal mold of a ventral valve (LN201403).
Remarks
Haydenella heilongjiangensis Li and Gu, Reference Li and Gu1980 and Haydenella libera Li in Li et al., Reference Li, Yang and Feng1986 are similar to the present species with their shared transverse outlines, but the former two have acuter cardinal extremities and a hingeline equal to the maximum width.
Tribe Paucispiniferini Muir-Wood and Cooper, Reference Muir-Wood and Cooper1960
Genus Transennatia Waterhouse, Reference Waterhouse1975
Type species
Productus gratiosus Waagen, Reference Waagen1884 from the Capitanian of Salt Range, Pakistan.
Remarks
With the type species Productus gratiosus Waagen, Reference Waagen1884, Waterhouse (Reference Waterhouse1975, p. 10) and Grant (Reference Grant1976, p. 131), respectively, proposed the genera Transennatia and Gratiosina. On the basis of the Law of Priority, Transennatia, which was characterized by its typical reticulation formed by strong costae and rugae on the visceral part, was formally accepted in the Treatise (Brunton et al., Reference Brunton, Lazarev, Grant, Jin and Williams2000, p. 447). In South China, Zhan (in Hou et al., Reference Hou, Zhan and Chen1979) established the genus Asioproductus (with the type species Asioproductus bellus), which is widely distributed in Lopingian strata and was considered a junior subjective synonym of Transenatia (Brunton et al., Reference Brunton, Lazarev, Grant, Jin and Williams2000, p. 447; for detailed discussions see Shi et al., Reference Shi, Shen and Zhan2003).
Transennatia gratiosa (Waagen, Reference Waagen1884)
Figure 10.9–10.20
- Reference Waagen1884
Productus gratiosus Waagen, p. 691, pl. 72, figs. 3–7.
- Reference Frech1911
Productus gratiosus; Frech, p. 127, pl. 19, fig. 4a, b.
- Reference Chao1927
Productus gratiosus; Chao, p, 44, pl. 4, figs. 6–10.
- Reference Huang1932
Productus (Dictyoclostus) aff. gratiosus; Huang, p. 32, pl. 2, fig. 3.
- Reference Huang1932
Productus (Dictyoclostus) cf. P. gratiosus; Huang, p. 33, pl. 2, fig. 4–5.
- Reference Grabau1934
Productus gratiosus; Grabau, p. 34, pl. 10, figs. 4–6.
- Reference Licharew1937
Productus (Marginifera) gratiosus var. timorensis Hamlet; Licharew, p. 118, pl. 4, figs. 11–19; pl. 9, fig. 37.
- Reference Zhang and Jin1961
Dictyoclostus gratiosus; Zhang and Jin, p. 411, pl. 4, figs. 12–18.
- Reference Wang, Jin and Fang1964
Dictyoclostus gratiosus; Wang et al., p. 291, pl. 45, figs. 14–19.
- Reference Yang, Ni, Chang and Zhao1977
Asioproductus gratiosus; Zhan in Yang et al., p. 350, pl. 140, fig. 5.
- Reference Feng and Jiang1978
Asioproductus gratiosus Zhan in Yang et al.; Feng and Jiang, p. 254, pl. 90, figs. 1, 2.
- Reference Tong1978
Asioproductus gratiosus Zhan in Yang et al.; Tong, p. 228, pl. 80, fig. 7.
- Reference Licharew, Kotlyar and Popeko1978
Gratiosina gratiosa (Waagen); Licharew and Kotlyar, pl. 20, fig. 1a, b.
- Reference Hou, Zhan and Chen1979
Asioproductus gratiosus; Zhan in Hou et al., p. 85, pl. 6, figs. 7–13; pl. 9, figs. 8–10.
- Reference Wang, Liu, Jin, Hu, Liang and Liao1982
Transenatia gratiosa (Waagen); Wang et al., p. 214, pl. 92, figs. 6–8; pl. 103, figs. 4–9.
- Reference Shi and Chen2003
Transenatia gratiosa (Waagen); Shi et al., p. 1059, fig. 3.15–3.19.
Holotype
Cephalopod bed, Salt Range, Pakistan (Waagen, Reference Waagen1884, pl. 72, fig. 3).
Occurrence
Carboniferous to Permian; cosmopolitan.
Description
Shell medium to large for genus, outline subquadrate, maximum width at hingeline or midvalve. Ventral valve strongly convex, geniculate at posterior part; ears small; sulcus narrow and deep. External surface with strong costae, converging into ventral sulcus, sometimes inconsistent in breadth in some specimens; concentric rugae developed, resulting in reticulated at visceral disc; body spine sparsely scattered, absent from sulcus, with a pair of spines located on the anterolateral slope. Interior with dense endospines, variable in size; a weak and short median septum developed.
Dorsal valve slightly to moderately concave, maximum concavity at middle to anterior part; ears medium in size, well demarcated from visceral disc; fold distinct and narrow, originating from beak. Exterior with strong costae, originating from beak, bifurcating at the most anterior part, covering the whole shell but ears; concentric rugae strongly developed on the whole shell except for a narrow circle near the margins, resulting in the very distinctly and densely reticulate ornament. Interior with radically distributed endospines, becoming thicker around brachial ridges; a pair of elongate adductor marks distinctly developed; a thin and weak median septum originating from umbo, extending about one-third of shell length.
Material
Three internal molds of ventral valves (LN182901, LN215901, LN234801), five external molds of dorsal valves (LN227301, LN233001, LN233201, LN234903, LN235403), and an internal mold of a dorsal valve (LN233101).
Remarks
The present species is like Transennatia sulcata Campi and Shi, Reference Campi and Shi2007 with a ventral convexity and outline, but the latter has much coarser costae without converging into sulcus. It also resembles Transennatia anshunensis Liao, Reference Liao1980 in the development of sulcus and shell outline, but the latter differs in having much coarser costae.
Transennatia insculpta (Grant, Reference Grant1976)
Figures 10.21–10.24, 11, 12.1–12.12
- Reference Grant1976
Gratiosina insculpta Grant, p. 135, pl. 32, figs. 1–37, pl. 33, figs. 1–16.
Holotype
USNM 212386, Ko Muk, southern Thailand (Grant, Reference Grant1976, pl. 32, figs. 25–28).
Occurrence
Permian; China, Malaysia, Thailand.
Description
Shell medium to large for genus, reverse trapezoid to subtriangular in outline, maximum width mostly at hingeline. Ventral valve strongly convex, with maximum convexity at middle to posterior part; ears large, moderately extended, ornamented with concentric rugae; sulcus moderately developed, originating from beak or midvalve. External surface covered with distinct costae, beginning from beak and bifurcating at middle to anterior part, becoming thinner and convergent in sulcus; concentric rugae evenly and densely distributed; visceral disc strongly geniculate by crossing of costae and rugae; several body spines scattered on valve. Interior with radially distributed endospines.
Dorsal valve slightly to moderately concave, with maximum concavity at anterior part where a distinct geniculation normally developed; beak narrow and pointed; ears large; fold moderately to distinctly developed. Exterior with costae, distinctly originating from beak, with bifurcation at middle to anterior valve; concentric rugae strong and densely distributed, resulting in prominently reticulate visceral region; 2–3 columns of microspines observed in the anterior part of costae between geniculation and margins (Fig. 11.17). Interior with fine endospines, becoming coarse around brachial ridges; a pair of elongate adductor marks distinctly developed; cardinal process trilobed or quadrivalve.

Figure 11. Brachiopods from the Shuizhutang Formation: T. insculpta. (1) a conjoined shell, LN226203; (2, 3, 5, 7–10, 13, 16, 18–20, 22, 24, 25) external molds of dorsal valves: (2) LN227001, (3) LN228601, (5) LN230401, (7) LN235003, (8) LN238001, (9) LN224603, (10) LN170103, (13) LN225801, (16) LN227701, (18) LN227903, (19) LN229003, (20) LN229603, (22) LN229703, (24) LN230201, (25) LN232403; (17) enlarged rectangular area in (16); (4, 6, 11) internal molds of ventral valves: (4) LN229203, (6) LN232001, (11) LN225403; (12, 14, 15, 21, 23) lateral views of (11, 13, 16, 20, 22), respectively. Scale bars = 3 mm.
Material
A ventral valve (LN205201), 23 external molds of dorsal valves (LN224403, LN224901, LN225203, LN227001, LN228601, LN230401, LN235003, LN238001, LN224603, LN170103, LN225801, LN227701, LN227903, LN229003, LN229603, LN229703, LN230201, LN232403, LN233901, LN234303, LN235302, LN238303, LN238803), a conjoined shell (LN226203), an internal mold of a dorsal valve (LN236301), and four internal molds of ventral valves (LN229203, LN232001, LN236503, LN225403).
Remarks
Grant (Reference Grant1976, p. 136) compared the differences between T. insculpta and T. gratiosa and argued that the “Thailand species (T. insculpta) differs from the Pakistan species (T. gratiosa) in being only about half the size, having somewhat more extended ears, a proportionately shorter, narrower and less distinctly lobate cardinal process, weaker lateral adductor muscle marks, indistinct taleolae in the ventral valve, and a single row of about 5 lateral spines along each flank rather than a dense band that may be considered as many as 3 rows as in T. gratiosa.” However, the Pakistan specimens are weathered and too obscure to identify the type of cardinal process and development of adductor muscle and taleolae in the interior. Both the Thailand and Pakistan specimens have spines on the flank area of ventral valve. Herein, we suggest that T. gratiosa can be distinguished from the current species by having less-extended ears, geniculate ventral valve, and sometimes more-distinct sulcus and fold. Transennatia timorensis Hamlet, Reference Hamlet1928 is comparable to the present species in that both have a moderately convex ventral valve, but it differs in having much coarser costae and rugae.
Family Productidae Gray, Reference Gray1840
Subfamily Leioproductinae Muir-Wood and Cooper, Reference Muir-Wood and Cooper1960
Tribe Tyloplectini Termier and Termier, Reference Termier and Termier1970
Genus Tyloplecta Muir-Wood and Cooper, Reference Muir-Wood and Cooper1960
Type species
Productus scabriculus mut. nankingensis Frech, Reference Frech1911 from the early Permian of Anhui Province, China.
Remarks
A global review of morphology and classification of the genus has been given in Shi and Chen (Reference Shi and Chen2003).
Tyloplecta huadongensis Liang, Reference Liang1990
Figure 12.13–12.19
- Reference Liang1990
Tyloplecta huadongensis Liang, p. 191, pl. 32, figs. 1–9, pl. 33, fig. 10.

Figure 12. Brachiopods from the Shuizhutang Formation. (1–12) T. insculpta: (1, 2, 5, 10, 11) external molds of dorsal valves: (1) LN233901, (2) LN234303, (5) LN235302, (10) LN238303, (11) LN238803; (3, 4, 9, 12) lateral views of (2, 5, 10, 11), respectively; (6) an internal mold of a dorsal valve, LN236301; (7) an internal mold of a ventral valve, LN236503; (8) lateral view of (7). (13–19) Tyloplecta huadongensis: (14, 15) external molds of dorsal valves: (14) LN161303, (15) LN163903; (13) lateral view of (14); (16) dorsal interior, LN184902; (17, 19) internal molds of ventral valves: (17) LN261403, (19) LN261903; (18) posterior view of (16). (20–29) T. liannanensis n. sp.: (20) ventral exterior, LN185601; (21) lateral view of (20); (22, 27) external molds of dorsal valves: (22) LN160803, (27) LN165903; (23) an internal mold of a dorsal valve, LN163703; (24, 26, 28, 29) internal molds of ventral valves: (24) LN164803, (26) LN165403, (28) LN173103, (29) LN175203; (25) posterior view of (24). (30) Tyloplecta sp., an external mold of a dorsal valve, LN214001. (31) Linoproductus huananensis n. sp., an external mold of a ventral valve, LN163303. Scale bars = 1 cm.
Holotype
ZB50088, Lengwu Formation, Lengwu, Zhejiang Province, China (Liang, Reference Liang1990, pl. 32, figs. 1–4).
Occurrence
Permian; Asia, Europe.
Description
Shell large, suboval to elongate oval in outline, hingeline slightly shorter than the greatest width of the shell. Ventral valve strongly inflated, strongly geniculated at the posterior part, becoming moderately convex and regularly inclined after this geniculation; beak pointed and enrolled; ears small and inflated; sulcus moderately developed, beginning from anterior part of beak. External surface marked by thin and distinct costae, visible on margins of internal mold. Internal surface with a pair of radial adductor muscle scars at the anterior part of beak, a pair of large and pectinate diductor muscle scars at each side of adductor muscle scars; endospines observed on ears; fine and densely distributed pits on the surface anterior to muscle scars.
Dorsal valve moderately concave, marginal area of anterior and lateral parts of the shell geniculated, with maximum concavity at anterior part; ears small and inflated; beak slightly concave, not or slightly over hingeline; fold absent to slightly developed. External surface with low costae, developed mainly on visceral region; concentric rugae covering the whole shell, especially strong on ears.
Material
Two external molds of dorsal valves (LN161303, LN163903), a dorsal interior (LN184902), and three internal molds of ventral valves (LN261403, LN261703, LN261903).
Remarks
The present species differs from most species in the genus by its thin costae on the ventral valve and weak nodules caused by crossing costae and rugae. It is similar to Tyloplecta bulangensis Liao, Reference Liao1980 in that both have thin costae, but the latter has a lower beak and large and flat ears. By having obscure nodes and thinner costae, the present species could be distinguished from Tyloplecta yangtzeensis.
Tyloplecta liannanensis new species
Figure 12.20–12.29
Type specimens
Holotype, an external mold of a dorsal valve (LN160803); paratype, an internal mold of a ventral valve (LN175203).
Diagnosis
Medium to large Tyloplecta with not enrolled ventral beak, moderately convex ventral valve, and thin costae.
Occurrence
Wuchiapingian; China.
Description
Shell medium to large, reverse trapezoid in outline, with hingeline marking the greatest width of the shell. Ventral valve moderately convex and regularly inflated; the longitudinal curve regular and gradual; in the transverse direction, the median portion almost flat and gently inclined toward both sides; beak narrow and pointed, not enrolled, slightly over hingeline, with an angle of umbonal region of about 100°; ears flat to slightly convex, large and marked off from the reminder of the valve by a distinct concavity; cardinal extremities slightly extended, with cardinal angles of about 60°–90°; sulcus absent or very weakly developed, represented mostly by a very shallow impression along the median portion of the shell. Costae thin, rarely bifurcating anteriorly, about 6–8 at anterior margin; rugae thin, present only on visceral disc, formed obscure nodes with costae. Interior with a pair of distinct radial adductor muscle scars in the middle at the anterior part of beak, a pair of wide and strong diductor muscle scars at each side; endospines randomly arranged on ears, umbonal region and anterior part, varied in size; strong pits developed on the anterior region of muscle scars.
Dorsal valve moderately concave, maximum concavity at anterior part; in the longitudinal direction, the anterior portion distinctly inclined dorsally; ears large, triangular, slightly concave; cardinal extremities slightly extended; fold weak, beginning from midvalve. Exterior covered with low costae, originating from anterior part of beak; secondary capillae developed on the whole shell except ears; concentric rugae existed from beak to margins, strong on shell from hingeline to lateral slope, very weak or nearly absent. Interior with very fine pits.
Etymology
Named for the Liannan County where the Liannan section is located.
Other material
A ventral exterior (LN185601), an external mold of a dorsal valve (LN165903), an internal mold of a dorsal valve (LN163703), and three internal molds of ventral valves (LN164803, LN165403, LN173103).
Remarks
The species can be distinguished from most species in the genus as it lacks an enrolled ventral beak and has a less convex ventral valve and thinner costae. Tyloplecta bulangensis Liao, Reference Liao1980 and Tyloplecta fulingensis Zhu, Reference Zhu1990 also have a low ventral beak that is not enrolled, but the former differs from the present species due to a subcircular outline and wider umbonal region, and the latter is differentiated from the present species by its more obtuse cardinal extremities.
Tyloplecta sp.
Figure 12.30
Occurrence
Wuchiapingian; China.
Description
Shell medium size, reverse trapezoid in outline, maximum width at hingeline. Dorsal valve moderately convex, maximum convexity at midvalve, regularly curved in longitudinal and transverse directions; ears moderate in size and inflated; cardinal extremities blunt, with cardinal angles of about 90°; sulcus absent. Exterior with randomly scattered body spines; costae sparsely distributed; concentric lirae developed mainly on ears and marginal areas; coarse nodes prominently developed mainly on the visceral region. Interior with quadrivalve cardinal process.
Material
An external mold of a dorsal valve (LN214001).
Remarks
The present species differs from all known species in the genus by its much smaller size, reverse trapezoid outline, and prominent nodes.
Superfamily Linoproductoidea Stehli, Reference Stehli1954
Family Linoproductidae Stehli, Reference Stehli1954
Genus Linoproductus Chao, Reference Chao1927
Type species
Productus cora d'Orbigny, Reference d'Orbigny1842 from the Moscovian of Patapatani, Bolivia, and the Asselian of Yaurichambi, Bolivia.
Linoproductus huananensis new species
Figures 12.31, 13.1–13.8

Figure 13. Brachiopods from the Shuizhutang Formation. (1–8) Linoproductus huananensis n. sp.: (1) an external mold of a dorsal valve, LN163103; (2, 3) enlargements of rectangles in (1) and (8), respectively; (4, 8) external molds of dorsal valves: (4) LN182103, (8) LN228302; (5) an external mold of a dorsal valve with part of internal mold of a ventral valve and part of ventral valve, LN212601; (6) an internal mold of a ventral valve, LN212901; (7) a ventral exterior, LN213601. (9–12) Licharewiella costata, internal molds of ventral valves: (9) LN194901, (10) LN202503, (11) LN206501, (12) LN210001. (13–24) L. liangyangensis, internal molds of ventral valves: (13) LN168200, (14) LN204000, (15) LN166603, (16) LN201603, (17) LN201803, (18) LN202203, (19) LN202703, (20) LN203102, (21) LN203302, (22) LN203703, (23) LN203902, (24) LN205702. (25) Leptodus lacuna Liang, Reference Liang1990, an internal mold of a ventral valve, LN240403. Scale bars = 1 cm.
Type specimens
Holotype, an external mold of a dorsal valve (LN228302); paratype, a ventral exterior (LN213601).
Diagnosis
Medium-sized Linoproductus with moderately convex ventral valve and slightly inflated ventral beak.
Occurrence
Wuchiapingian; China.
Description
Medium size for genus, outline rounded quadrate to transversely suboval, maximum width at midvalve. Ventral valve slightly to moderately convex, maximum convexity at umbo; ears small and slightly convex, not marked off from body of valve; umbo slightly overhanging hingeline; sulcus absent. External surface covered with very fine and closely set capillae; concentric rugae strongly and densely developed on ears, weak on visceral region; coarse body spines scattered on the whole valve. Interior with densely arranged endospines, coarse and water-drop-shaped in the middle part, fine and rounded in other areas.
Dorsal valve slightly to moderately concave, maximum concavity normally at midvalve; ears small and slightly concave; umbo pointed to moderately wide; fold absent. External surface with very fine capillae, with lots of bifurcations and intercalations; concentric rugae strong on ears and weaken toward midvalve.
Etymology
Named for the South China area (Huanan) where the Liannan section is located.
Other material
Three external molds of dorsal valves (LN163103, LN182103, LN228302), an external mold of a ventral valve (LN163303), an external mold of a dorsal valve with part of internal mold of a ventral valve and part of ventral valve (LN212601), and an internal mold of a ventral valve (LN212901).
Remarks
This species can be easily differentiated from most counterparts in the genus by its moderately convex ventral valve and absence of an enrolled ventral beak. Linoproductus capillatus Zhan in Hou et al., Reference Hou, Zhan and Chen1979 and Linoproductus fujianensis Wang et al., Reference Wang, Liu, Jin, Hu, Liang and Liao1982 also have rounded outlines and low convexity of ventral valve but differ from the present species by much more developed rugae, and L. fujianensis has coarser capillae. Linoproductus pigrami Archbold, Reference Archbold1981 is similar to the present species in the low convexity of ventral valve and development of capillae and rugae but differs by having larger and pointed ears and more flattened profile.
Suborder Strophalosiidina Schuchert, Reference Schuchert1913
Superfamily Strophalosioidea Schuchert, Reference Schuchert1913
Family Strophalosiidae Schuchert, Reference Schuchert1913
Subfamily Strophalosiinae Schuchert, Reference Schuchert1913
Genus Licharewiella Ustritsky in Ustritsky, Hu, and Chan, Reference Ustritsky, Hu and Chan1960
Type species
Strophalosia costata Waagen, Reference Waagen1884 from the Wordian of Salt Range, Pakistan.
Remarks
The internal structure of the genus has rarely been reported. The Liannan section yields some well-preserved ventral valves, providing interior information of the genus. The ventral interior of Licharewiella has a very strong and short median septum, a series of linear diductor muscle scars in the posterior part, and endospines variably developed.
Licharewiella costata (Waagen, Reference Waagen1884)
Figure 13.9–13.12
- Reference Waagen1882
Strophalosia costata Waagen, p. 655, pl. 63, figs. 7, 8, pl. 64, fig. 1.
- Reference Yang, Ni, Chang and Zhao1977
Licharewiella costata; Yang et al., p. 335, pl. 136, fig. 14.
Holotype
Unknown. Waagen (Reference Waagen1884) did not designate a holotype for this species.
Occurrence
Permian; China, Pakistan.
Description
Shell medium size, transversely oval in outline, maximum width at midvalve. Ventral valve moderately convex; ears small and slightly inflated; beak truncated or moderately pointed; sulcus slightly developed, originating from anterior part of beak. Interior with strong and short median septum, about one-eighth to one-fourth of shell length; linear diductor muscle scars developed on both sides of median septum; endospines coarse, restricted to anterior part of shell. Exterior with strong costae, beginning from anterior part of beak, becoming coarser and stronger forward.
Material
Four internal molds of ventral valves (LN194901, LN202503, LN206501, LN210001).
Remarks
The species is similar to Licharewiella plicosa Waagen, 1884 in the rounded outline, but the latter has thinner costae and more-pointed ventral beak.
Licharewiella liangyangensis Zhan in Hou et al., Reference Hou, Zhan and Chen1979
Figure 13.13–13.24
- Reference Hou, Zhan and Chen1979
Licharewiella liangyangensis Zhan in Hou et al., p. 75, pl. 6, fig. 18.
- Reference Liao1982
Truncatenia heshanensis Liao, p. 540, pl. 1, figs. 10–15, 18–23.
Holotype
K0083, Shuizhutang Formation, Lianxian, Guangdong, China (Hou et al., Reference Hou, Zhan and Chen1979, pl. 6, fig. 18).
Occurrence
Wuchiapingian; China.
Description
Shell medium size for genus, transversely reverse trapezoid to subquadrate in outline, maximum width at hingeline. Ventral valve moderately to strongly convex, maximum convexity at posterior part; ears variable in size, slightly inflated, smooth or partly covered with costae; umbonal region wide and obtuse, sometimes strongly truncated; sulcus absent. Interior with very strong and short median septum, about one-tenth to one-sixth of shell length; a series of linear diductor muscle scars developed in the front of median septum; fine endospines observed mainly in the visceral region. Exterior with strong and simple costae, beginning from the front of beak, occasionally intercalation and bifurcation in the anterior part; one to three pairs of spines distributed on each side of hinge.
Material
Twelve internal molds of ventral valves (LN168200, LN204000, LN166603, LN201603, LN201803, LN202203, LN202703, LN203102, LN203302, LN203703, LN203902, LN205702).
Remarks
The species can be easily differentiated from its counterparts by its outline and wide hingeline. Truncatenia Liao, Reference Liao1982 was considered a synonym of Licharewiella by Kaesler (Reference Kaesler2000b). Truncatenia heshanensis Liao, Reference Liao1982 is characterized by a reverse trapezoid outline and coarse costae on the valve and has no difference from the current species; thus, it should be a synonym of L. liangyangensis.
Suborder Lyttoniidina Williams, Harper, and Grant in Kaesler, Reference Kaesler2000
Superfamily Permianelloidea He and Zhu, Reference He and Zhu1979
Family Permianellidae He and Zhu, Reference He and Zhu1979
Genus Permianella He and Zhu, Reference He and Zhu1979
Type species
Permianella typica He and Zhu, Reference He and Zhu1979 from the Lopingian of Jiangxi and Sichuan Province, China.
Remarks
When He and Zhu (Reference He and Zhu1979) first reported this genus, they stated that the shell featured pseudopunctae on the inner surface and had a smooth outer surface with sparse and weak concentric striae. Later, Wang and Jin (Reference Wang and Jin1991) provided a detailed discussion of permianellids, covering their shape, ornament, and shell structures. Although based on Dicystoconcha and lyttoniids specimens instead of Permianella, Wang and Jin (Reference Wang and Jin1991) concluded that the shell of the whole group developed tubercles on the outer surface and pseudopunctae on the inner surface. They stated that the tubercles are about 150 μm in diameter and unevenly distributed, and pseudopunctae are about 10 μm in diameter and also unevenly distributed. However, on the basis of the very well-preserved specimens from the Liannan section, it was found that the tubercles and pseudopunctae are very similar in size. Tubercles are evenly distributed and somewhat concentrically arranged in the middle to anterior part and number about 30–50 per mm2. Pseudopunctae are also evenly distributed, absent at marginal brims, and number about 40–50 per mm2.
Permianella typica He and Zhu, Reference He and Zhu1979
Figure 14.10, 14.11, 14.15–14.17
- Reference He and Zhu1979
Permianella typica He and Zhu, p. 132, pl. 1, fig. 1, pl. 2, 3.
- Reference Wang and Jin1991
Permianella typica; Wang and Jin, p. 496, pl. 2, figs. 1–3.

Figure 14. Brachiopods from the Shuizhutang Formation. (1–5) Leptodus lacuna, internal molds of ventral valves: (1) LN262403, (2) LN262803, (3) LN262903, (4) LN263803, (5) LN268003. (6–8) Oldhamina squamosa, internal mold of a ventral valve, LN263203: (6) ventral view, (7) posterior view, (8) lateral view. (9) Perigeyerella sp., an internal mold of a ventral valve, LN190803. (10, 11, 15–17) Permianella typica: (10) an external mold of a dorsal valve with part of inner core, LN262101; (15, 16) respectively, enlargements of upper and lower rectangles in (10); (11) an external mold of a ventral valve, LN262201; (17) enlargement of rectangle in (11). (12) Derbyia sp., an internal mold of a ventral valve, LN177902. (13, 14) Alatorthotetina derbyiformis He and Zhu, Reference He and Zhu1985: (13) an internal mold of a dorsal valve, LN240102; (14) an internal mold of a ventral valve, LN166204. (18–23) Orthothetina frechi (Huang, Reference Huang1933): (18, 20, 21) internal molds of ventral valves: (18) LN239403, (20) LN239203, (21) LN240902; (19, 22, 23) internal molds of dorsal valves: (19) LN239503, (22) LN241202, (23) LN182603. (24–29) Peltichia kwangtungensis (Zhan in Hou et al., Reference Hou, Zhan and Chen1979): (24, 28) internal molds of dorsal valves: (24) LN172502, (28) LN180101; (25–27) an inner core of a conjoined shell, LN172703: (25) dorsal view, (26) posterior view, (27) ventral view; (29) an internal mold of a ventral valve, LN240700. (1–14, 18–29) Scale bars = 1 cm; (15–17) scale bars = 2 mm.
Syntype
Specimens 19791, 19796, 19799, Longtan Formation, Jiangxi and Sichuan Province, China (He and Zhu, Reference He and Zhu1979, pl. 1, fig. 1; pl. 2, fig. 3; pl. 3, fig. 3).
Occurrence
Permian; China, Japan, Malaysia.
Description
Shell of large size for genus, elongated bilobate in outline, greatest width at middle to anterior part; length 63.3 mm, width 25.3 mm, hingeline width 6.7 mm, length of incision 45.3 mm; posterolateral margins converging posteriorly at an umbonal angle about 70°, lateral margins nearly parallel. Ventral valve slightly convex in anterior profile and nearly flat in lateral profile; sulcus distinct, about one-fourth of shell length; external surface ornamented with fine and dense tubercles, sometimes concentrically arranged at the visceral region, randomly and more densely distributed at marginal brim; internal surface with randomly arranged pseudopunctae, absent near margins. Dorsal valve also flattened in lateral profile and slightly concave in anterior profile; fold beginning from the front of beak, widening and heightening anteriorly; exterior with very fine and densely distributed tubercles.
Material
An external mold of a ventral valve (LN262201) and an external mold of a dorsal valve with a part of inner core (LN262101).
Remarks
There are only two species and a few specimens of the genus reported. It differs from Permianella grunti Shen and Shi, Reference Shen and Shi1997 by its larger umbonal angle, parallel lateral margins, and flattened shells. The specimen is slightly different from the material in Campi et al. (Reference Campi, Shen, Shi and Leman2000) by the latter having a smooth shell except fine radial fila on posterior half of valve.
Suborder Athyrididina Boucot, Johnson, and Staton, Reference Boucot, Johnson and Staton1964
Superfamily Athyridoidea Davidson, Reference Davidson1881
Family Athyrididae Davidson, Reference Davidson1881
Genus Araxathyris Grunt in Ruzhentsev and Sarytcheva, Reference Ruzhentsev and Sarytcheva1965
Type species
Spirigera protea Abich, 1878 from the Lopingian of Azerbaijan.
Araxathyris minor new species
Figure 15.12–15.16
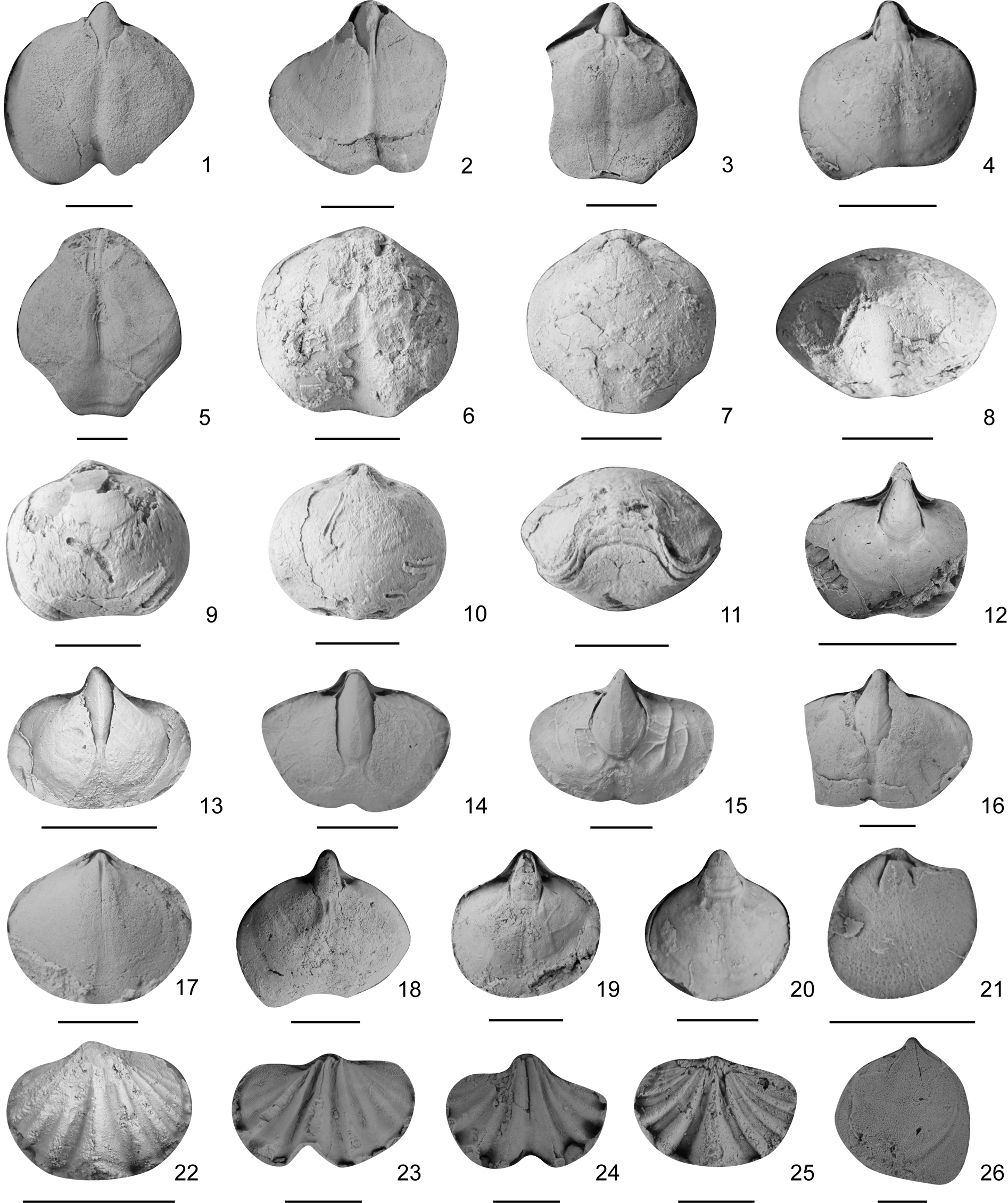
Figure 15. Brachiopods from the Shuizhutang Formation. (1–4) Araxathyris beipeiensis, internal molds of ventral valves: (1) LN244300, (2) LN246503, (3) LN248903, (4) LN249101. (5) A. glossexserta Zeng et al., Reference Zeng, He and Zhu1995, an internal mold of a ventral valve, LN245003. (6–11) A. indentatus Zeng et al., Reference Zeng, He and Zhu1995, two inner cores of two conjoined shells: (6–8) LN192903: (6) ventral view, (7) dorsal view, (8) anterior view; (9–11) LN193903: (9) ventral view, (10) dorsal view, (11) anterior view. (12–16) A. minor n. sp., internal molds of ventral valves: (12) LN000101, (13) LN000104, (14) LN245200, (15) LN245303, (16) LN245903. (17–20) A. undulata Shen et al., Reference Shen, He and Zhu1992: (17) an internal mold of a dorsal valve, LN246802; (18–20) internal molds of ventral valves: (18) LN247501, (19) LN247701, (20) LN248701. (21) Crurithyris sp., an internal mold of a dorsal valve, LN186902. (22–25) Spiriferellina nasuta Cooper and Grant, Reference Cooper and Grant1976: (22, 23, 25) internal molds of dorsal valves: (22) LN001303, (23) LN178301, (25) LN180401; (24) an internal mold of a ventral valve, LN178401. (26) Qinglongia sp., an internal mold of a dorsal valve, LN176404. Scale bars = 5 mm.
Type specimens
Holotype, an internal mold of a ventral valve (LN245303); paratype, an internal mold of a ventral valve (LN000101).
Diagnosis
Small Araxathyris with large and strong spondylium.
Occurrence
Wuchiapingian; China.
Description
Shell small for genus, transversely suboval in outline, greatest width around shell midlength. Ventral valve moderately convex, posterior sides nearly straight and horizontal, lateral sides rounded, and anterior side slightly concave; sulcus slightly to moderately developed, beginning from midlength, deepening and widening anteriorly. Interior with very strong and large spondylium, highly overhanging hingeline, extending about half to two-thirds of shell length.
Etymology
Named for its very small size.
Other materials
Three internal molds of ventral valves (LN000104, LN245200, LN245903).
Remarks
This species can be easily differentiated from other species by its very small size and very strong and large spondylium. Data for spondylium size and body-size of all specimens of the genus in the Liannan section were collected and are shown in Figure 16. It is clear that spondylium size in the present species is distinctly separated from, and larger than, other species and is a stable character because it steadily grows as body size increases.

Figure 16. Graph of spondylium size to body size of all Araxathyris specimens from the Liannan section.
Suborder Delthyridina Ivanova, Reference Ivanova1972
Superfamily Reticularioidea Waagen, Reference Waagen1883
Family Elythidae Fredericks, Reference Fredericks1924
Subfamily Phricodothyridinae Caster, Reference Caster1939
Genus Permophricodothyris Pavlova, Reference Pavlova1965
Type species
Permophricodothyris ovata Pavlova, Reference Pavlova1965 from the Wuchiapingian of Azerbaijan.
Remarks
There are 24 species in the genus. Most species are distinguished by shell size, convexity, development of the sulcus and fold, and distribution patterns of concentric lamellae. The development, shape, and position of muscle scars, which have been clearly shown with sketch by Shi et al. (Reference Shi, Chen and Han2002), were rarely referred to when comparing different species in the genus. Herein, the very well-preserved Liannan specimens distinctly show the different development of muscle scars.
Permophricodothyris affinis (Gemmellaro, Reference Gemmellaro1899)
Figure 17.1–17.9
- Reference Gemmellaro1899
Reticularia affinis Gemmellaro, p. 194, pl. 34, figs. 5–8; pl. 46, figs. 10, 11.
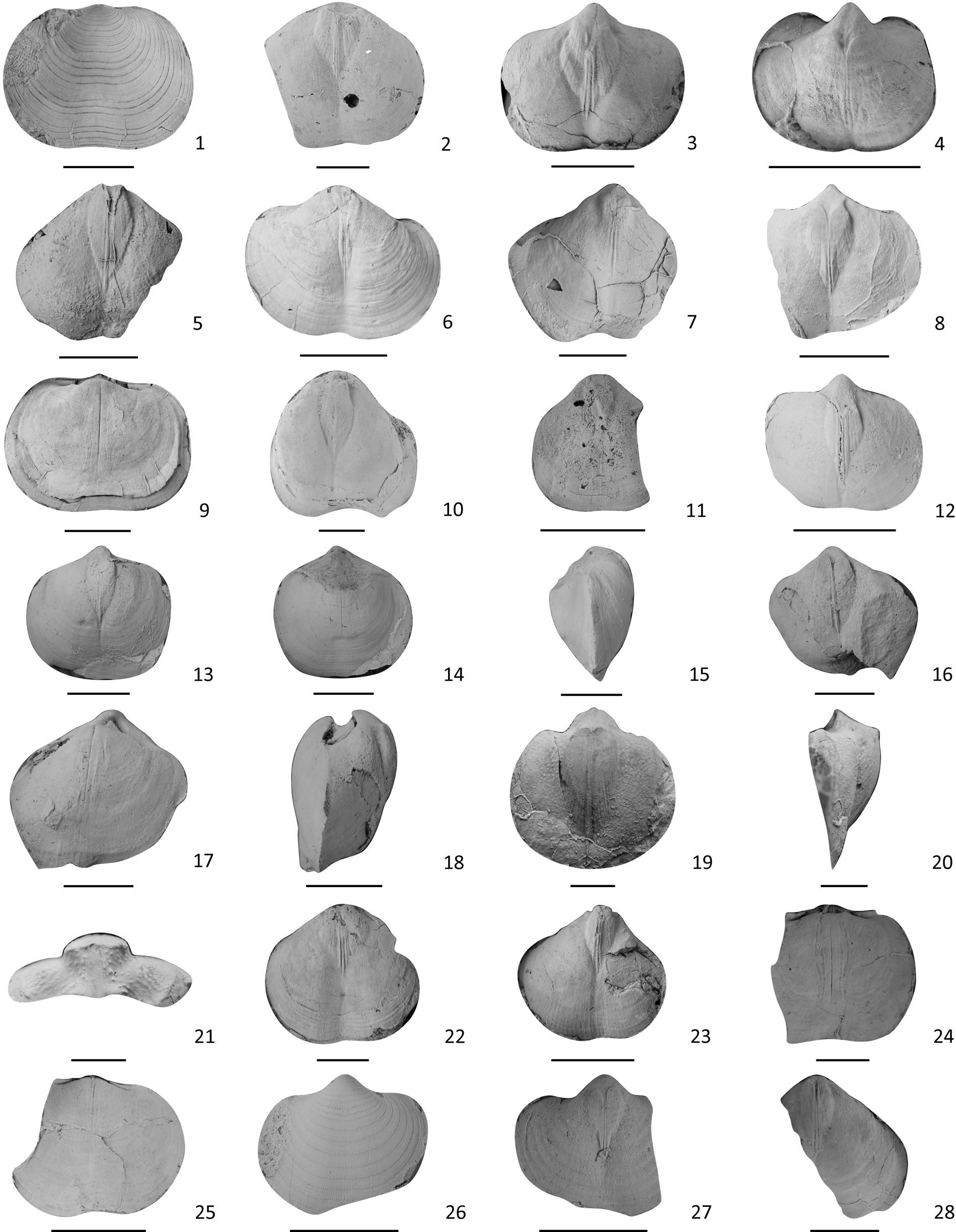
Figure 17. Brachiopods from the Shuizhutang Formation. (1–9) Permophricodothyris affinis: (1) an external mold of a dorsal valve, LN169403; (2–8) internal molds of ventral valves: (2) LN170303, (3) LN186403, (4) LN187103, (5) LN250403, (6) LN252403, (7) LN252702, (8) LN252903; (9) an internal mold of a dorsal valve, LN256303. (10–21) P. extensiformis: (10–12, 19) internal molds of ventral valves: (10) LN161903, (11) LN168403, (12) LN254102, (19) LN162503; (20, 21) lateral and posterior views of (19), respectively; (13–18) two inner cores of two conjoined shells: (13–15) LN254102: (13) ventral view, (14) dorsal view, (15) anterior view; (16–18) LN260702: (16) ventral view, (17) dorsal view, (18) anterior view. (22–28) P. nodosa: (22, 23, 27, 28) internal molds of ventral valves: (22) LN166403, (23) LN171703, (27) LN183103, (28) LN183503; (24, 25) internal molds of dorsal valves: (24) LN179503, (25) LN181603; (26) an external mold of a ventral valve, LN182503. Scale bars = 1 cm.
Holotype
Unknown. Gemmellaro (Reference Gemmellaro1899) did not designate a holotype for this species.
Occurrence
Guadalupian to Lopingian; China, Italy, Tunisia, Turkey.
Description
Shell small to medium for genus, moderately biconvex; outline suboval, sometimes moderately transverse, greatest width normally at or posterior to midlength of ventral valve. Ventral valve moderately convex; beak thick, moderately incurved; sulcus distinct, beginning from the front of beak, deepening anteriorly; interior with heart-shaped muscle scars, including three linear adductor scars in the middle and a pair of flabellate diductor scars composed of 7–10 radial lobes on each side. Dorsal valve slightly and more evenly convex, beak blunt and short; interior with a thin and long myophragm, beginning from front of beak, extending to about three-fourths of shell length; socket elongate and wide, inner socket ridge and outer socket ridge thin. Exterior with irregularly arranged concentric lamellae, each with one or two rows of thorny spines.
Material
An external mold of a dorsal valve (LN169403), seven internal molds of ventral valves (LN170303, LN186403, LN187103, LN250403, LN252403, LN252702, LN252903), and an internal mold of a dorsal valve (LN256303).
Remarks
The present species can be easily separated from other species by the narrow and distinct ventral sulcus. Permophricodothyris caroli (Gemmellaro, Reference Gemmellaro1899) also has a prominent ventral sulcus but differs by having a much higher and narrower ventral beak.
Permophricodothyris extensiformis (Chang in Yang et al., Reference Yang, Ni, Chang and Zhao1977)
Figure 17.10–17.21
- Reference Yang, Ni, Chang and Zhao1977
Squamularia extensiformis Chang in Yang et al., p. 452, pl. 180, fig. 6.
Holotype
IV47210, Qixia Formation, Hunan Province, China (Chang in Yang et al., Reference Yang, Ni, Chang and Zhao1977, pl. 180, fig. 6).
Occurrence
Permian; China.
Description
Medium size for genus, suboval to roundly subquadrate in outline, biconvex with ventral valve equal to or more inflated than dorsal valve; commissure rectimarginate; surface with irregularly spaced growth lamellae. Ventral beak moderately hanging over hingeline, thick and incurved; interarea small; sulcus absent; interior with large and distinct heart-shaped muscle scars, with a strong and linear adductor scar in the middle and a pair of flabellate diductor scars on each side. Dorsal valve slightly to moderately convex, maximum convexity at umbo; fold absent; interior with a thin and weak myophragm, extending about half of shell length.
Material
Four internal molds of ventral valves (LN161903, LN168403, LN254102, LN162503) and two inner cores of two conjoined shells (LN254102, LN260702).
Remarks
The present species is characterized by rounded outline and lateral profile, lack of sulcus and fold, and blunt umbo. It is similar to Permophricodothyris jiangshuiensis (Chang in Yang et al., Reference Yang, Ni, Chang and Zhao1977) in the lack of sulcus, fold, and outline, but the latter has a more-pointed ventral beak and more-prominent concentric lamellae. Permophricodothyris notialasiatica Grant, Reference Grant1976 also has no sulcus and fold developed but differs by having a much lower and pointed beak.
Permophricodothyris nodosa (Chao, Reference Chao1929)
Figures 17.22–17.28, 18.1–18.12
- Reference Chao1929
Squamularia nodosa Chao, p. 95, pl. 11, figs. 4–6.
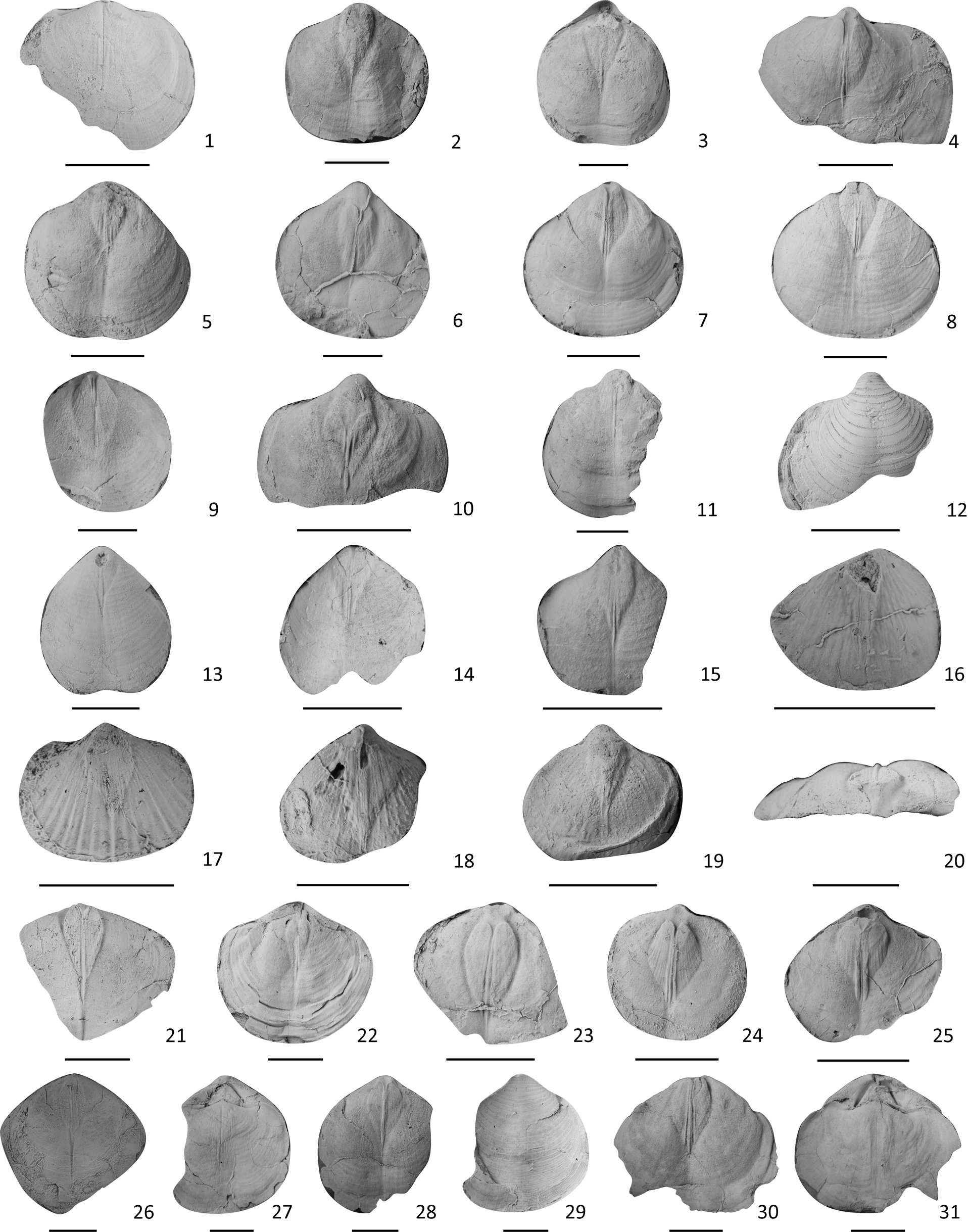
Figure 18. Brachiopods from the Shuizhutang Formation. (1–12) Permophricodothyris nodosa: (1) an internal mold of a dorsal valve, LN185203; (2–11) internal molds of ventral valves: (2) LN253303, (3) LN249803, (4) LN250003, (5) LN250203, (6) LN250702, (7) LN250903, (8) LN251403, (9) LN251603, (10) LN251803, (11) LN259203; (12) an external mold of a ventral valve, LN255603. (13–15) P. ovata: internal molds of ventral valves: (13) LN170700, (14) LN251003, (15) LN255101. (16–19) Permophricodothyris sp., internal molds of ventral valves: (16) LN176901, (17) LN183301, (18) LN188704, (19) LN253901. (20–31) P. flata n. sp.: (21–26) internal molds of ventral valves: (21) LN183904, (22) LN251104, (23) LN252503, (24) LN253103, (25) LN253703, (26) LN175804; (20) posterior view of (21); (27, 28, 30, 31) two inner cores of two conjoined shells: (27, 28) LN259302: (27) ventral view, (28) dorsal view; (30, 31) LN259802: (30) ventral view, (31) dorsal view; (29) the external mold of the ventral valve of (28). Scale bars = 1 cm.
Syntype
Specimens 2028, 2029, 2030, Permian coal series, Jiangxi Province, China (Chao, Reference Chao1929, pl. 11, figs. 4–6).
Occurrence
Permian; China, Thailand, United States.
Description
Shell medium size for genus, with suboval outline; surface with regularly spaced concentric lamellae, each with two rows of thorny spines. Ventral valve moderately convex, maximum convexity at umbonal region, the remaining part of the valve becoming less convex; beak blunt and slightly incurved; sulcus slightly to moderately developed; interior with distinct heart-shaped or rhombic muscle scars, with three strong and linear adductor scars in the middle and a pair of flabellate diductor scars on both sides, sometimes with radiate pallial muscular markings covering the whole surface. Dorsal valve slightly convex; fold slightly developed; interior with a thin myophragm, originating from the front of beak and extending about half of shell length; socket elongate rounded, inner socket ridge and outer socket ridge thin.
Material
Fourteen internal molds of ventral valves (LN166403, LN171703, LN183103, LN183503, LN253303, LN249803, LN250003, LN250203, LN250702, LN250903, LN251403, LN251603, LN251803, LN259203), three internal molds of dorsal valves (LN179503, LN181603, LN185203), and two external molds of ventral valves (LN182503, LN255603).
Remarks
The present species is most like Permophricodothyris elegantula (Waagen, Reference Waagen1883), with low convexity of both valves and a weak sulcus and fold, but the latter has very delicate and small beaks. Permophricodothyris incerta Zhan in Li et al., Reference Li, Zhan, Dai, Jin, Zhu, Zhang, Huang, Xu, Yan and Li1989 also has moderately convex shells but differs by hav9ng a much more elongate outline.
Permophricodothyris ovata Pavlova, Reference Pavlova1965
Figure 18.13–18.15
- Reference Pavlova1965
Permophricodothyris ovata Pavlova, p. 86, fig. 4.
Holotype
Specimen 2071/181, Dorasham II, Azerbaijan (Pavlova, Reference Pavlova1965, fig. 4A–D).
Occurrence
Wordian to Changhsingian; Armenia, Azerbaijan, Iran, China.
Description
Shell about medium size, elongated ovate in outline. Ventral valve strongly convex, very regularly curved in both directions; beak moderately wide and pointed; sulcus slightly developed; interior with weak muscle scars, with a linear adductor scar in the middle and flabellate diductor scars on both sides; external surface with regularly distributed concentric lamellae.
Material
Three internal molds of ventral valves (LN170700, LN251003, LN255101).
Remarks
The present species is characterized by the very regular elliptical outline of the shell, strongly convex valve, and weak sulcus. Permophricodothyris globosa (Feng in Feng and Jiang, Reference Feng and Jiang1978) is similar to P. ovata in outline and development of the sulcus, but P. globosa differs by having a much more inflated ventral valve. It is similar to Permophricodothyris grandis (Chao, Reference Chao1929) in Shi et al. (Reference Shi, Chen and Han2002) in outline and convexity, but the latter has much larger size and stronger muscle scars.
Permophricodothyris flata new species
Figure 18.20–18.31
Type specimens
Holotype, an internal mold of a ventral valve (LN253103); paratype, an inner core of two conjoined shells (LN259302).
Diagnosis
Medium to large Permophricodothyris with slightly convex to nearly flattened shells, a low and slightly inflated ventral beak, and a suboval to subcircular outline.
Occurrence
Wuchiapingian; China.
Description
Medium to large size for genus, outline suboval to subcircular, greatest width normally around midlength of ventral valve; commissure rectimarginate; concentric lamellae numbering about 4–5 per 5 mm at middle to posterior part, about 12–13 per 5 mm at anterior part, each with a row of thorny spines. Ventral valve flat to slightly and evenly convex; beak low and blunt, slightly or not curved; interarea small; delthyrium bounded by deltidial flange on each side, partly occupied by dorsal beak. Interior with distinct and strong muscle scars, heart shaped or diamond shaped, bisected by a short median ridge, sometimes located at almost the middle of valve (Fig. 18.24); three linear adductor muscle marks strong in the middle, flabellate diductor scars on each side.
Dorsal valve slightly convex; beak short and more pointed; ears slightly concave. Interior with deep and elongate sockets, thin and long socket ridges; myophragm weak and long, beginning from umbonal region and extending to anterior part; muscle marks weakly impressed.
Etymology
Named for its nearly flat profile.
Other material
Five internal molds of ventral valves (LN183904, LN251104, LN252503, LN253703, LN175804) and an inner core of two conjoined shells (LN259802).
Remarks
It can be easily distinguished from most of its counterparts by shell convexity. Compared with most species in the genus, P. incerta and P. nodosa have much less convex shells but still have more inflated shells than the present species. Moreover, P. incerta differs by having a more elongated outline, and P. nodosa differs by having a higher and more pointed ventral beak.
Permophricodothyris sp.
Figure 18.16–18.19
Occurrence
Wuchiapingian; China.
Description
Shell small for genus, with suboval outline. Ventral valve moderately convex; beak moderately high and pointed, with an apical angle of 90°–110°; sulcus absent; interior with strong and relatively large muscle scars, linear adductor scars strong in the middle, and diductor scars flabellate on each side; coarse and strong pustules radially arranged around muscle platform margins. In some specimens, strong linear impressions of external laminae evenly and radially distributed and covering the whole shell surface (Fig. 18.17).
Material
Four internal molds of ventral valves (LN176901, LN183301, LN188704, LN253901).
Remarks
The present species is very different from all known species in the genus. Although with small size, it has strong and relatively large muscle scars (0.51 < muscle scar length/shell length < 0.69) and very strong pustules around the muscle scars covering most parts of the inner surface.
Conclusions
A late Wuchiapingian brachiopod fauna, which contains 57 species of 28 genera and is much more diversified than all reported contemporaneous brachiopod faunas from South China, is described. Four new species, Tyloplecta liannanensis, Linoproductus huananensis, Araxathyris minor, and Permophricodothyris flata are proposed. Comparison of the Douling fauna and the Shuizhutang fauna reveals that the brachiopod fauna shows a recovery mainly on the generic level during the survival stage and a rapid radiation on both generic and specific levels during the late Wuchiapingian.
Acknowledgments
We are very grateful for the constructive helpful reviews by G.R. Shi, an anonymous reviewer, R. Zhan, S. Zamora, and the managing editor. The authors thank D. Xi for his help in fossil photography. This paper has been supported by the Natural Science Foundation of China (grant no. 41902008) and the Fundamental Research Funds for the Central Universities (grant no. 2652018131, no. 00/800015A302).
Declaration of competing interests
The authors declare none.
Data availability statement
Data available from the Dryad Digital Repository: http://doi.org/10.5061/dryad.gxd2547q2



















