


Exposure to stressful life events is common (see for example Breslau et al Reference Breslau, Davis, Andreski and Peterson1 ) and is associated with psychiatric distress, most notably internalising symptomology (see for example Galea et al, Reference Galea, Brewin, Gruber, Jones, King and King2 Kendler et al Reference Kendler, Kessler, Walters, MacLean, Neale and Heath3 and Kessler et al Reference Kessler, Sonnega, Bromet, Hughes and Nelson4 ). Variability in response to stressful life events has been demonstrated, with some individuals experiencing psychiatric symptoms, whereas others remain asymptomatic. Reference Bonanno5 The concept of resiliency, a term that is often used to describe adaptive levels of functioning in the face of an adversity, is of great interest. Reference Rutter6 There are wide-reaching public health implications resulting from a better understanding of the aetiological factors related to resilience, and refining this phenotype is a key step towards this goal.
Resilience has been defined in different ways, Reference Luthar, Cicchetti and Becker7 leading to divergent findings regarding the contributing aetiological factors. For example, the heritability estimate of behavioural and cognitive resilience to economic deprivation in youth was roughly 70%, Reference Kim-Cohen, Moffitt, Caspi and Taylor8 whereas the heritability of resilience as defined by positive affect in the face of social and interpersonal stressors in adults was estimated at 38% for women and 52% for men. Reference Broadman, Blalock and Button9 Neither the adversities nor the outcomes measured in the two extant genetically informed studies shed light on resilience as it is often conceptualised in the field of psychiatry, and the present study aims to fill this gap.
Luthar and colleagues Reference Luthar, Cicchetti and Becker7 suggest that studies of resilience should: focus on events that have high associated risk for the outcome of interest; carefully choose the post-stressor domain of assessment, as stressors may have an impact on more than a single outcome; and define resilience along a continuum rather than as a arbitrary dichotomy. In light of these recommendations, our study used a large, longitudinal study of adult twins who were assessed for exposure to a variety of stressful life events that have been shown to have an impact on internalising symptoms, that measured global internalising psychiatric symptoms rather than focusing on narrow diagnoses, and lastly, defined resilience as a quantitative variable. We sought to answer four questions. First, to what degree do genetic v. environmental factors contribute to individual differences in resilience to stressful life events in adults? Second, is resilience to these events, and the genetic and environmental factors that contribute to this resilience, stable over time? Third, are there quantitative gender differences for this phenotype (i.e. are genetic factors for resilience more important for males than females)? Finally, are there qualitative gender effects (i.e. are the genes involved in heritability of resilience different in males than females)?
Method
Participants
Participants were derived from two interrelated Virginia Adult Twin Studies of Psychiatric and Substance Use Disorders (VATSPSUD) studies of White individuals, Reference Kendler and Prescott10 ascertained from the birth-certificate-based Virginia Twin Registry. Female-female (FF) twin pairs, born 1934-1974, were eligible if both members responded to a mailed questionnaire in 1987-1988. Data in this report utilised interviews completed at the first (FF1) and third interview wave (FF3), conducted in 1987-1989 and 1992-1995 respectively. Reference Kendler and Prescott10 Cooperation at the FF1 and FF3 interviews was 92% and 88% respectively (and 93% for the intervening second interview wave). Reference Kendler and Prescott10 Data on the male-male and male-female pairs (MMMF) came from a sample (birth years 1940-1974) initially ascertained, with a 72% cooperation rate, directly from registry records containing all twin births. The first interview (MMMF1) was completed largely by telephone in 1993-1996. The second wave of interviews (MMMF2), conducted in 1994-1998, had a retention rate of 83%. Zygosity was determined by discriminate function analyses using standard twin questions validated against DNA genotyping in 496 pairs. Reference Kendler and Prescott11 The mean age and years of education of the twins were 35.1 (s.d. = 7.5) years and 14.3 (s.d. = 2.2) at the FF3 interview, and 37.0 (s.d. = 9.1) years and 13.6 (s.d. = 2.6) at the MMMF2 interview.
These analyses utilised data from 7500 twins, including both members of 3084 pairs (503 monozygotic (MZ) female-female, 346 dizygotic (DZ) female-female, 703 MZ male-male, 485 DZ male-male, and 1047 opposite-gender DZ pairs) and 1325 twins without their co-twin. (These numbers do not sum because all possible pairings for triplet and quadruplet sets were included).
Internalising symptoms
Participants completed a shortened version of the Symptom Checklist-90 (SCL-90), Reference Derogatis, Lipman and Covi12 which utilised a past-month time frame. There were 27 items from four of the SCL subscales: depression (10 items), somatisation (5 items), anxiety (7 items), phobic anxiety (5 items) and 3 items that assessed sleep difficulty. All items had five response options: not at all, a little bit, moderately, quite a bit and extremely. This measure demonstrated high internal reliability for both waves (Cronbach’s α = 0.74).
Stressful life events
We assessed stressful life events that were personal in nature (assault, serious marital problems, divorce, job loss, loss of a confidant, serious illness, major financial problems, being robbed, serious legal problems), and ‘network’ events (i.e. events that occurred primarily to, or in interaction with, an individual in the respondent’s social network): death or severe illness of the respondent’s spouse, child, parent, co-twin or other relative, serious trouble getting along with others close to the respondent. All stressful life events were assessed using a past-year time frame. Interrater reliability for determining the occurrence of the events was high (kappa (κ) = 0.93). Reference Kendler, Kessler, Walters, MacLean, Neale and Heath3 A sum of stressful life events was computed.
Definition of resilience
Resilience was operationalised as the residual of the SCL score after the effect of recent number of stressful life events had been regressed out for each wave (i.e. the difference between actual and predicted SCL). If a twin’s SCL was lower than predicted by the regression, this would result in a negative residual, reflecting resilience; if a twin’s SCL was higher than expected, this would result in a positive residual, reflecting low levels of resilience.
Twin modelling
Phenotypic variation in twin models is decomposed into additive genetic factors (A), which contribute twice as much to the correlations between MZ twins as they do for DZ twins; common environmental factors (C), which are the shared factors (for example parental attitudes, economic disadvantage) that make twins reared together more similar and contribute equally to the correlation between MZ and DZ twins; and individual-specific environmental (E) sources, which reflect environmental experiences not shared by twins and therefore contribute to differences between the twins and errors of measurement.
We tested for qualitative gender differences (i.e. whether or not the same genetic factors influenced liability to resilience for males and females, quantified by rg ), and for quantitative gender differences (i.e. if there is equality in the estimates of the genetic contribution in males and females) by fitting three models that were compared with the saturated model. First, we constrained rg to 1.0 to test for qualitative gender effects. Second, we tested for quantitative gender effects by constraining the estimates of A, C and E to be equal in males and females. Finally, we computed a model to test for both qualitative and quantitative gender effects. To evaluate the fit of the twin models, full-information maximum likelihood approach to raw data was implemented in Classic Mx Reference Neale, Boker, Xie and Maes13 on a Windows-based PC. Akaike’s information criteria (AIC) is used as a guide to evaluating different models and produces an index of goodness of fit (i.e. balance of explanatory power and parsimony). Reference Akaike14 Parsimony is important because in maximum likelihood estimation, log likelihoods will continue to decrease with additional model parameters potentially resulting in ‘over-fitting’. Akaike’s information criteria penalises models with increasing numbers of parameters thereby providing an appropriate balance between model complexity and explanatory power as manifest by the degree of misfit. Reference Williams and Holahan15
Univariate models
We conducted univariate models of the wave 1 and 2 data, fitting the saturated models and models with constraints for qualitative and quantitative gender effects. After choosing the best-fit ACE model from the gender-specific testing from the wave 1 and 2 models, we then fitted subsequent models in which parameters were dropped to determine the most parsimonious model (AE and CE models).
Measurement models
We then utilised both waves of data simultaneously in a measurement model that assumes that there is a true latent liability to resilience. Each of the two estimates of resilience is considered to be a fallible index of the true latent liability. The paths λ1 and λ2 represent the degree to which the assessments of resilience obtained at the two waves reflect the true liability. The product of these paths reflects the reliability (or long-term stability) of the assessment of resilience. The latent liability, as well as wave-specific liability, for resilience was then modelled in a standard ACE twin design, as outlined above, including testing for gender effects. Two notable differences from the standard twin model and the measurement model are present. First, the model allows for separate estimates of occasion-specific influences (including measurement error) and true or enduring individual-specific environmental effects. Second, an estimate of the degree to which the time-specific measurements of resilience are reflective of the true latent liability for resilience is produced.
Results
Our definition of resilience relies on two premises: first, stressful life events are associated with the level of internalising symptoms, and second, there is variability in responses to stressful life events. Indeed, as the number of such events increases the average total SCL score also increases, as does the variability around the point estimate (Fig. 1). The relationship between stressful life events and SCL scores was monotonic, with no evidence of a ceiling effect. Stressful life events were significantly and very similarly associated with SCL scores for wave 1 (β = 0.21, R 2 = 0.03, P<0.001) and wave 2 (β = 0.20, R 2 = 0.03, P<0.001). From these regressions we saved the standardised residuals, which represent resilience to the effects of stressful life events on internalising symptoms.
Univariate twin models
The between-twin Pearson correlation of resilience was roughly twice as large for MZ pairs as it was for DZ pairs for males (rs = +0.30 and +0.16 respectively) and females (rs = +0.34 and +0.16 respectively). Table 1 outlines the models that were fitted for wave 1 resilience. Models were compared with the saturated model (I). We began in model II by testing for qualitative gender effects. Model III tests for quantitative gender effects, and model IV tests for both gender effects. There was evidence for qualitative but not quantitative gender effects. Therefore, model III was chosen to fit the submodels (AE; CE). These results suggested that C could be dropped from the model without sacrificing fit (model VI), but A could not be dropped (model V). Consequently, the AE model with qualitative gender effects was the best-fit model (V; Fig. 2). The genetic correlation (rg ) is substantial (+0.76), but falls short of unity.
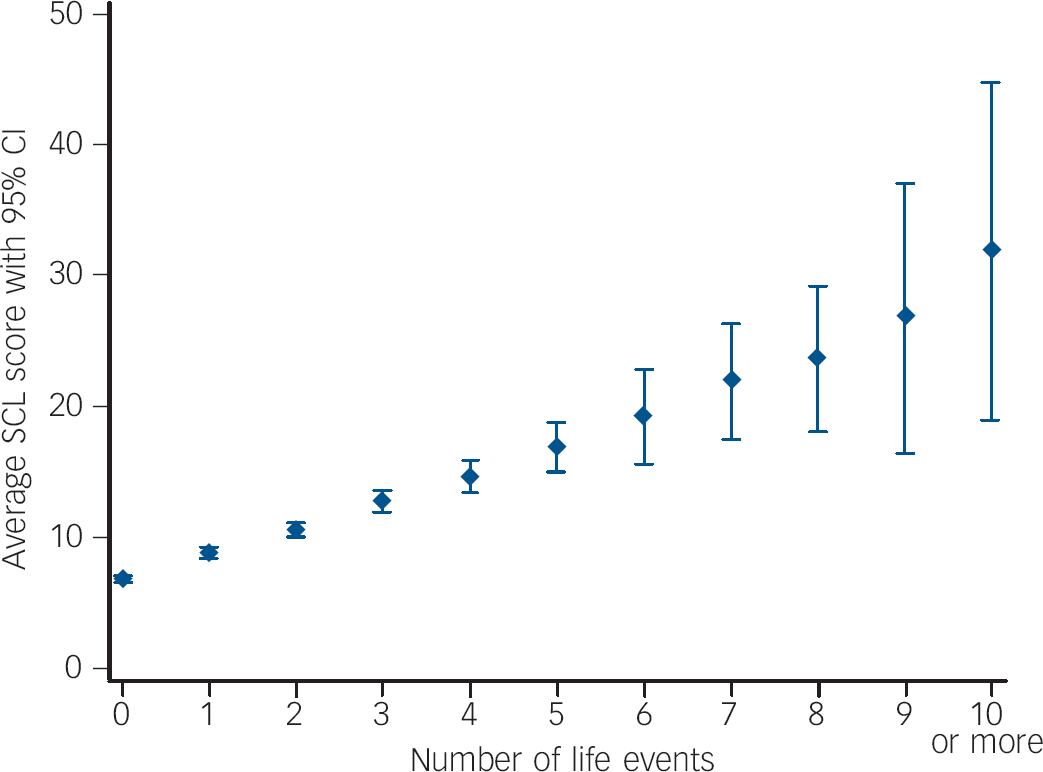
Fig. 1 Number of stressful life events and average Symptom Checklist (SCL) total score for wave 1.
Models fit for wave 2 are presented in Table 1. In contrast to wave 1, for wave 2 the best-fit model was model X, without qualitative or quantitative gender effects. The AE and CE models were computed next, suggesting that C could be dropped from the model (XII) but A could not (XI).
Longitudinal twin model
We conservatively began by fitting the longitudinal ACE measurement model with qualitative but not quantitative gender effects estimated (model I). From this model we then fit the AE and CE submodels. As shown in Table 2, dropping C resulted in a superior model fit (model II), whereas dropping A resulted in a deterioration in fit (model III). From the AE model other constraints were tested to examine whether parsimony and fit could be improved. We constrained the λ to equality to test whether the wave 1 and 2 assessments reflected with the same accuracy the underlying latent level of resilience. This constraint resulted in an improved fit (model IV; Fig. 3). We tested whether the time-specific A estimates could be set to 0 for waves 1 and 2, models V and VI, respectively. Model fit deteriorated for both of these constraints, making model IV our final model. In this model, r g was high, but statistically different from unity (+0.88).
Table 1 Results of model fitting to determine influence of genetic and environmental factors on resilience for waves 1 and 2

| Model | Variables | Qualitative/ quantitative |
-2LL | AIC | d.f. | Δd.f. | Δ(-2LL) | ΔAIC |
|---|---|---|---|---|---|---|---|---|
| Wave 1 | ||||||||
| I | ACE | + / + | 24530.05 | 6808.05 | 8861 | - | - | - |
| II | ACE | - / + | - | - | - | 1 | 0.43 | –1.57 |
| III | ACE | + / - | - | - | - | 2 | 1.32 | –2.69 |
| IV | ACE | - / - | - | - | - | 3 | 3.51 | –2.49 |
| V | AE | + / - | - | - | - | 3 | 1.32 | –4.69 |
| VI | CE | + / - | - | - | - | 4 | 42.74 | 34.74 |
| Wave 2 | ||||||||
| VII | ACE | + / + | 19681.87 | 5435.87 | 7123 | - | - | - |
| VIII | ACE | - / + | - | - | - | 1 | 0.000 | –2.00 |
| IX | ACE | + / - | - | - | - | 2 | 0.051 | –3.95 |
| X | ACE | - / - | - | - | - | 3 | 0.051 | –5.95 |
| XI | AE | - / - | - | - | - | 4 | 0.069 | –7.93 |
| XII | CE | - / - | - | - | - | 4 | 22.432 | 14.43 |
–2LL, –2 log likelihood; AIC, Akaike’s information criteria; A, additive genetic factor; C, common environmental factor; E, individual-specific environmental factor.
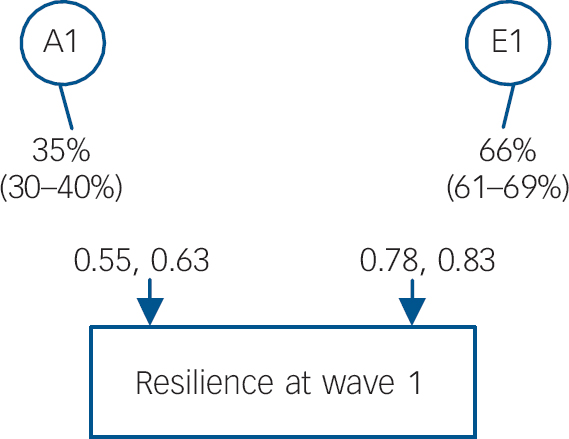
Fig. 2 Percent of variance (with 95% CI) accounted for from the best-fitting model for genetic and environmental factors for resilience at wave 1: female-female and male-male twins.
Opposite-gender twins: genetic correlation rg = 0.38 estimated; -2 log likelihood (-2LL) = 24531.369 (d.f. = 8864), Akaike’s information criteria = 6803.369. A1, first (and only) additive genetic factor; E1, first (and only) individual-specific environmental factor.
There are five notable aspects of the final model. First, each wave provided a good index of the true level of resilience with the predicted stability of our measure across waves equal to +0.53. Second, for wave 1, the overall heritability was 31%, and of this, roughly 90% of this estimate is stable across time and driven by the latent measure, and 10% was occasion specific. Similarly, for wave 2, the overall heritability was 32%, with approximately 86% coming from the stable latent measure, and the remaining 14% was time specific. Third, when unconfounded by measurement error the heritability increased (∼50%). Fourth, qualitative gender effects were found. Fifth, for both waves larger proportions of the variance in E was as a result of the occasion-specific effects (i.e. transient environmental effects and measurement error) than was as a result of environmental factors with an enduring effect (i.e. those accounted for by the latent E). Specifically, 61% of the variance in E was time specific for both waves, with the remaining variance attributed to stable E.
Discussion
This study marks the first estimation of the genetic and environmental contributions to psychiatric resilience to stressful life events, as well as the first estimation of the stability of the resilience phenotype and its aetiology assessed over time. The longitudinal nature of this study afforded the ability to incorporate unreliability of measurement of resilience (which is influenced by error of measurement from two sources: the stressful life events assessment and internalising symptom assessment) directly into our twin model, allowing for the separation of error of measurement and occasion-specific effects from the enduring environmental differences. In doing so, a striking conclusion can be made: genetic constitution and enduring environmental influences contribute roughly equally to the latent construct of resilience. Here we discuss each component in turn, and then end with a comment on implications for measurement of this important construct.
Table 2 Results of measurement model fitting to determine influence of genetic and environmental factors on resilience for waves 1 and 2
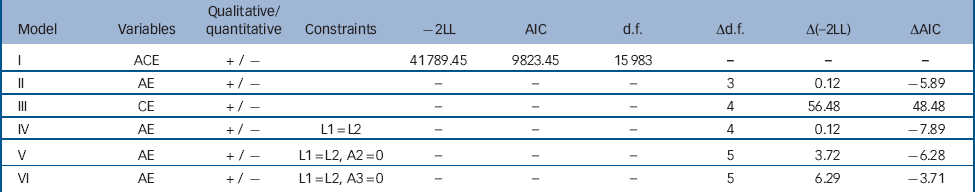
| Model | Variables | Qualitative/ quantitative |
Constraints | -2LL | AIC | d.f. | Δd.f. | Δ(-2LL) | ΔAIC |
|---|---|---|---|---|---|---|---|---|---|
| I | ACE | + / – | 41 789.45 | 9823.45 | 15 983 | - | - | - | |
| II | AE | + / – | - | - | - | 3 | 0.12 | –5.89 | |
| III | CE | + / – | - | - | - | 4 | 56.48 | 48.48 | |
| IV | AE | + / – | L1 = L2 | - | - | - | 4 | 0.12 | –7.89 |
| V | AE | + / – | L1 = L2, A2 = 0 | - | - | - | 5 | 3.72 | –6.28 |
| VI | AE | + / – | L1 = L2, A3 = 0 | - | - | - | 5 | 6.29 | –3.71 |
–2LL, –2 log likelihood; AIC, Akaike’s information criteria; A, additive genetic factor; C, common environmental factor; E, individual-specific environmental factor; L, lambda; A1, additive genetic factor for resilience at wave 1; A2, additive genetic factor for resilience at wave 2.
Genetic contribution
We found a modest genetic influence for both waves of the study (heritability ∼0.31), suggesting that genes have an impact on the way that individuals respond in the face of stressful life events. In a qualitative sense, we can interpret the wave-specific heritability estimates in the ‘mild to moderate’ range. Although we could not develop an aetiological model for resilience without incorporation of genetic liability, genes are only contributing a modest amount for each measurement occasion. These heritability estimates are similar to those of anxiety disorders, Reference Kendler, Gardner and Prescott16 which are among the lower end of psychiatric disorders in terms of their genetic contribution. Reference Kendler and Prescott10 Another useful benchmark for comparison is with relevant personality factors (such as neuroticism, heritability ∼0.41). Reference Lake, Eaves, Maes, Health and Martin17 Our results are consistent with the two prior genetic investigations of resilience, which have each adopted a different definition of the stressor as well as the outcomes. Reference Kim-Cohen, Moffitt, Caspi and Taylor8,Reference Broadman, Blalock and Button9 Nonetheless, both studies produced mild to moderate heritability estimates. In short, as human traits go, the wave-specific heritability estimates of resilience are on the lower end of the spectrum.

Fig. 3 Percent of variance (with 95% CI) accounted for from the best-fitting model for genetic and environmental factors for resilience at waves 1 and 2.
A1, additive genetic factor on the latent measure of resilience; A2, A3, additive to genetic factors for wave 1 and wave 2 respectively; E1, individual-specific environmental factor on the latent measure of resilience; E2, E3, individual-specific environmental factors for wave 1 and wave 2 resilience respectively.
When unconfounded by measurement errors and occasion-specific influences, the heritability estimate for the latent factor increased substantially from the wave estimates; roughly 50% of the individual differences in resilience were attributed to the latent genetic factor. The construct of resilience is complex, and its measurement can introduce sources of error from how the stressor is measured to how the outcome is assessed. Thus, when these two sources of error are corrected for in the measurement model (which removes the transient environmental effects as well as true error), our heritability estimate was no longer in the ‘mild’ range (where the heritability estimates of most anxiety disorders are) but was well within the ‘moderate’ range, comparable with depression (see for example Kendler & Prescott Reference Kendler and Prescott10 ). Our finding that the ‘stable component’ of resilience is approximately 50% heritable adds to our understanding of the genetic architecture of the construct and is of value in increasing understanding regarding individual differences in responses to stressful life events. Given that resilience is influenced by genetic factors, future studies aimed at identification of specific genetic variation that accounts for the heritability are warranted. The majority of the genetic influence was driven by the latent measure of resilience, yet there was a modest amount of genetic variance that was occasion specific. These ‘new genes’ that are related to resilience may be demonstrating a time-dependent gene-expression effect related to development (for example maturity, learning, treatment). However, we do not know what these processes may be.
In contrast to Broadman et al, Reference Broadman, Blalock and Button9 we found that although the heritability of resilience is equivalent for males and females, the genes affecting resilience are not equivalent between males and females. This could suggest genetic differences between the genders in the contribution of sex hormones, fluctuations across the menstrual cycle in women, etc. This gene-environment correlation could also be accounted for by the ways in which men and women are socialised. Related phenotypes have also demonstrated qualitative gender effects (for example in major depression, alcohol dependence). Reference Kendler and Prescott10
How might we understand our demonstrated genetic contribution to resilience? Resilience research has its historical roots in developmental psychology, where temperamental factors (such as self-mastery, self-efficacy) have been associated with resilience in children exposed to adversity. Reference Rutter18,Reference Garmezy, Masten and Tellegen19 Research on adults has also documented the role of temperament and personality in determining response to stressful life events. It is plausible that shared genetic variance between resilience and these personality factors may be in play. For example, neuroticism is moderately heritable Reference Lake, Eaves, Maes, Health and Martin17 and has an effect on response to stressful life events in producing risk for internalising disorders. Reference Hettema, Neale, Myers, Prescott and Kendler20 Genetic influences may also exert their effects through a contribution to known protective factors for post-stressor response that are also moderately heritable (for example social support). Reference Kendler and Baker21 Alternatively, genetic influences may have an impact on resilience by directly shaping the way in which individuals react psychiatrically to stressful life events (for example affording psychological resources such as healthy coping techniques).
Environmental contribution
As important as our genetic findings are those regarding the environmental influences on resilience. This is the first study to quantify the degree to which environmental events affect resilience in an enduring fashion, as cross-sectional studies are confounded by errors of measurement. After partitioning out the measurement error and transient environmental effects, we found that roughly half of the contribution of variation in the latent level of resilience could be accounted for by individual-specific environmental events that had an enduring effect. This suggests that individual experiences are as important as genetic constitution for determining the latent level of resilience. Notably, the environmental estimates were influenced to a greater degree by occasion-specific effects than were the additive genetic effects, which were more stable and influenced to a higher degree by the latent genetic factor.
A large literature demonstrates the enduring negative health ramifications of exposure to types of extreme adverse events that may have been unshared by twins, such as childhood maltreatment (for example sexual abuse, physical abuse). Exposure can have lasting effects on acute stress reactivity Reference Loman and Gunnar22,Reference Heim, Newport, Heit, Graham, Wilcox and Bonsall23 and psychiatric functioning. Reference Nelson, Heath, Madden, Cooper, Dinwiddie and Bucholz24,Reference Kilpatrick, Ruggiero, Acierno, Saunders, Resnick and Best25 Evidence from neuroscience underscores the plasticity of the brain structure and function as influenced by exposure to environmental events, Reference Davidson and McEwen26 particularly with regard to circuit connectivity. Reference McEwen and Gianaros27 Epigenetic modifications are also associated with extreme stressful life events in genes associated with the hypothalamic-pituitary-adrenal axis, Reference McGowan, Sasaki, D'Alessio, Dymov, Labonte and Szyf28 which may be another mechanism through which the environmental influences exert their effect on resilience.
Alternatively, certain types of events unshared by twins may be ‘immunising’ (i.e. stress inoculation, steeling effects) and exposure may have buffered the individual from the effects of stressful life events. Findings from the preclinical literature have demonstrated that certain doses of stress can protect organisms from having high amounts of a stress reaction to adversity later in life. Reference Maier, Amat, Baratta, Paul and Watkins29 Notably, our quantification of resilience cannot recognise whether immunising or sensitising effects are at play, and this is an important direction for future research. Although the majority of the literature within traumatic stress suggests that trauma has a cumulative risk effect, Reference Hoge, Austin and Pollack30 there are studies that report that increased number of events can be related to an increased sense of locus of control. Reference Regehr, Hill and Glancy31 Supports could also have affected one twin (for example social support, higher education) that served to bolster resilience. However, whereas the majority of the additive genetic effects was stable and accounted for by the latent genetic factor, for the environmental estimates, larger proportions of the variance was accounted for by the time-specific effects (which also include measurement error) than was as a result of environmental factors with enduring effects. Nonetheless, identification of the sources of variation in the environmental events that have enduring effects on resilience is a critical step in this literature.
Defining resilience
Resilience has been defined in many different ways, with regard to the stressors that an individual would demonstrate resilience to when faced with and the phenotype that is characterised as a resilient response. Reference Hoge, Austin and Pollack30,Reference Davydov, Stewart, Ritchie and Chaudieu32 Our statistical approach is similar to Kim-Cohen et al’s, Reference Kim-Cohen, Moffitt, Caspi and Taylor8 who examined the difference between children’s scores on intelligence and behavioural scales and their predicted scores by level of socioeconomic deprivation; however, we focused on the types of stressors most likely to have an impact on internalising symptoms. Socioeconomic deprivation is a different type of stressor given its chronicity and diffuse impact. Our definition of resilience is appealing, as it does not rely on individuals to accurately report on a ‘higher level’ of observation. An alternative approach is to query individuals about their perceived ability to cope with adversities, Reference Hoge, Austin and Pollack30 which is more prone to reporting bias, errors in self-perception and variation with regard to the items included on the measure. Our objective approach provides a novel metric of resilience that statistically quantifies resilience.
Limitations
The present study should be interpreted in light of several weaknesses. First, resilience is a complex and dynamic process that unfolds over time Reference Rutter6 and our assessment method cannot fully capture this process, only the outcome. Specifically, our quantification of resilience is inferred, in one domain (internalising symptoms), and is based off of the individual variation to the stressful life events experiences. Second, as noted above, exposure to stressful life events may be associated with immunising effects, Reference Rutter6,Reference Regehr, Hill and Glancy31 as well as sensitising effects. Reference Loman and Gunnar22,Reference Heim, Newport, Heit, Graham, Wilcox and Bonsall23 Our quantification of resilience cannot differentiate between these effects. Second, although we assessed for a large number of stressful life events, our list was not exhaustive. To that end, we also did not assess for personal reaction to the event (such as fear, helplessness or horror) or the severity of the events. It is possible that individuals who may be resilient to some events may not show as much resilience to other events, thus a more thorough stressful-life-event assessment is warranted for future studies. Third, given demonstrated gene-environment correlation findings Reference Kendler and Baker21 for exposure to stressful life events, it is possible that a modest proportion of the association between adversity and the SCL measure is non-causal (i.e. genetically mediated through personality factors that influence exposure to stressful life events and psychopathology symptoms). Fourth, our outcome of interest was the SCL, which is a broad measure of internalising symptoms; we did not examine broader psychiatric disorders (for example externalising disorders). Fifth, although our sample is likely representative of twins in the Mid-Atlantic region of the USA in the 1990s, the generalisability to other areas and time frames is limited. Finally, we conservatively fitted the measurement model with qualitative gender effects, which were found for wave 1 but not for wave 2.






eLetters
No eLetters have been published for this article.