



Introduction
One of the stumbling blocks to understanding the evolutionary history of the genus Homo concerns the impact of the Early to Middle Pleistocene Transition (EMPT) (1.2–0.75Ma) on hominin populations. This period witnessed a shift in glacial cycle duration (from c. 41ka to c. 100ka) and amplitude (extreme thermal intensity) (Head & Gibbard Reference Head and Gibbard2005), leading to marked climate change. Concomitantly, cultural and biological innovations appear within the hominin lineage. Mode-2 technology and its typical Acheulean handaxes, which emerged earlier (c. 1.75Ma, Lepre et al. Reference Lepre, Roche, Kent, Harmand, Quinn, Brugal, Texier, Lenoble and Feibel2011), become more complex in design and diverse in areas of use, spreading to new regions of Africa and Eurasia at c. 1Ma (Pappu et al. Reference Pappu, Gunnell, Akhilesh, Braucher, Taieb, Demory and Thouveny2011; Gallotti & Mussi Reference Gallotti and Mussi2017). These technological changes may reflect diversification of hominin populations in the Early/Middle Pleistocene (Macchiarelli et al. Reference Macchiarelli, Bondioli, Chech, Coppa, Fiore, Russom, Vecchi, Libsekal and Rook2004). Only three East African sites have yielded well-preserved EMPT hominin fossils—Olorgesailie (Potts et al. Reference Potts, Behrensmeyer, Deino, Ditchfield and Clark2004); Bouri (Gilbert & Asfaw Reference Gilbert and Asfaw2008); and Buia (Macchiarelli et al. Reference Macchiarelli, Bondioli, Chech, Coppa, Fiore, Russom, Vecchi, Libsekal and Rook2004)—and there is no consensus regarding their interpretation within an integrated framework for human evolution.
Two major questions relate to EMPT hominin evolution: (1) what role did the environment play in prompting behavioural and biological changes; and (2) what factors shaped the diversity seen in the archaeological and fossil records during this period. Characterising these aspects is essential to deciphering whether different techno-cultural groups and hominin species existed at the time, and how those differences can be interpreted in a phylogenetic framework. The scarcity of EMPT sites in Africa prevents researchers from addressing these questions, making the discovery and description of new sites a priority.
The present project aims to identify and excavate EMPT sites in the south-west Turkana Basin (Kenya). This article introduces a new site currently under investigation: Kanyimangin.
Geology
Kanyimangin is located east of the Kerio fault, within the Kalabata River circular anomaly (Figure 1), where thick sedimentary deposits are present but limited in the north by later Quaternary deposits, reflecting lake level fluctuations. The sediments are dissected by diverging reverse faults, suspected to be older than 3.7Ma (Vétel et al. Reference Vétel, Gall and Johnson2004).

Figure 1. Location of Kanyimangin (figure by Aurélien Mounier and Ann Van Baelen).
Correlations between the layers of 10 geological trenches (GT) (Figure 2) at the site revealed around 15m of sedimentation history preserved by five sandstones (SS1–SS5) that form, west to east, successive tilted ridges in the landscape (approximately 14°W, Figure 2). To the west is a massive ridge formed by an archaeologically sterile pebble conglomerate. Below, without identified contact but on top of SS1, is a stratigraphic layer of laminated silts and sands (GT1) with a palaeosol forming its upper part, where faunal remains (carnivore, suid and microfauna) were found. Further east, the sediments preserved between SS2 and SS4 consist of fine, finely laminated but not very cohesive, silts and sands, forming an archaeologically sterile erosional surface. SS5 can be tracked from GT10 (north) to GT6 (south) (Figure 2), where archaeological and faunal remains are found in stratigraphic context.
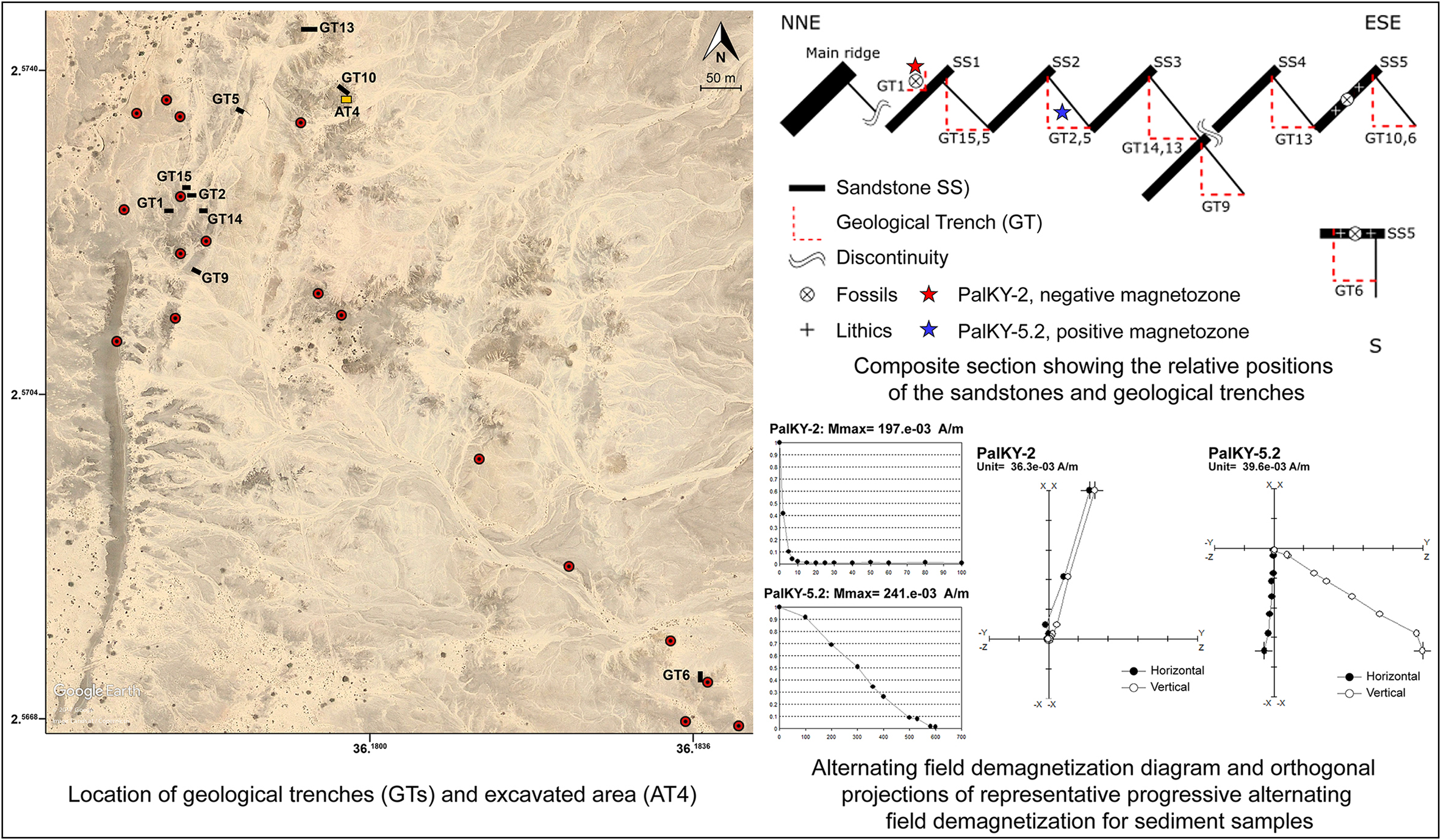
Figure 2. Map of Kanyimangin and geochronological analyses results (figure by Céline Vidal, Cécile Chapon-Sao and Aurélien Mounier).
Palaeomagnetic analyses indicate a reverse magnetozone for sediments from GT1 (PalKY-2, above SS1), and a normal magnetozone for sediments from GT2 (PalKY-5.2, below SS2) (Figure 2).
Archaeology
Archaeological surveys (2017–22) at Kanyimangin yielded 344 lithic artefacts, mostly discovered on erosional surface accumulations. In June 2022, an archaeological trench (AT4) (Figures 2 & 3) was excavated following discovery of fresh lithic artefacts on the sloping surface of a plateau in the northern part of the site. Seven square metres were excavated to a depth of 0.5m; archaeological material was found in sandy sediments and in pockets of brown-reddish silts surrounded by pieces of sandstone belonging to SS5. Sixty-two lithic artefacts were piece-plotted and five fossil fragments were recovered from the sieve (mesh size 2mm).

Figure 3. Lithic artefacts distribution (figure by Aurélien Mounier and Ann Van Baelen and Sol Sánchez-Dehesa Galán).
The excavated and surface artefacts display similar technological characteristics (Figure 4). Cores with simple debitage methods (unipolar and convergent), discoidal cores and different types of handaxes are consistent with an Early/Middle Pleistocene occupation of the site (e.g. Gallotti & Mussi Reference Gallotti and Mussi2017), although the presence of Levallois cores and flakes with platform preparation could indicate more recent occupational phases.

Figure 4. A selection of lithic artefacts from Kanyimangin (figure by Aurélien Mounier and Sol Sánchez-Dehesa Galán).
Palaeontology
The faunal assemblage recovered at Kanyimangin is composed of 2155 bone fragments representing 212 individual specimens from over 20 taxa (Figure 5). Aquatic and terrestrial taxa were identified using the National Museums of Kenya collections and included extinct elephants (Palaeoloxodon recki cf. recki), pigs (Kolpochoerus heseloni) and crocodiles (narrow snouted Euthecodon brumpti), big cats (Panthera sp.), Hyaena, zebra (Equus grevyi), Rhinocerotidae, Hippopotamus sp., warthog (Phacochoerus sp.), buffalo (Syncerus caffer), oribi (Ourebia ourebi), impala (Aepyceros melampus), hartebeest (Alcelaphus buselaphus), turtles/tortoises, snakes, fish and amphibians. While most of the assemblage was recovered during palaeontological survey, individuals preserving multiple elements were recovered through excavation in the areas surrounding two GTs: GT1 in the north, in the sediments lying on top of SS1, and GT6 in the south, in a layer including SS5 (Figures 5 & 6).

Figure 5. Fauna: geographical and specific distributions. In situ specimens (figure by Marjolein D. Bosch and Aurélien Mounier).

Figure 6. Fossil fauna recovered from Kanyimangin (figure by Aurélien Mounier).
Conclusion
The dating of Kanyimangin is based on the patterned palaeomagnetic sequence of the site's sediments in the context of a maximum biochronological age of elements of the fauna found within these. Geochronologically, the site preserves a sequence of a normal (SS2) followed by a reverse (GT1) magnetozone, which could represent the transition at the end of the Olduvai (c. 1.778Ma), Cobb Mountain (c. 1.173Ma) or Jaramillo (c. 0.988Ma) subchrons of the Matuyama chron (Horng et al. Reference Horng, Lee, Pälike, Wei, Liang, Iizuka and Torii2002). First appearance datums of Palaeoloxodon recki cf. recki (Beden Reference Beden1979), Phacochoerus sp. (Souron Reference Souron2012) and Alcelaphus buselaphus (Kingdon & Hoffmann Reference Kingdon and Hoffmann2013) are younger than the Olduvai subchron and their presence at the site makes Kanyimangin one of the few sites in East Africa known to date from the critical period, 1.2–0.75Ma. As further archaeological works unfold, Kanyimangin archaeological and palaeontological assemblages offer new opportunities to address questions about the EMPT impact on the behavioural and biological evolution of the genus Homo in Africa.
Acknowledgements
The authors wish to thank the In-Africa (ERC 295907) and Ng'ipalajem (ERC 101020478) Projects, National Museums of Kenya, Turkana Basin Institute, IFRA and French Embassy in Nairobi, and NACOSTI for permission (NACOSTI/P/15/2669/4758-NACOSTI/P/18/2669/22086-NACOSTI/P/19/513-NACOSTI/P/21/8475-NACOSTI/P/22/17817).
Funding statement
The project was supported by funding from IRP CNRS-INEE (LIA1262), the French Foreign Office (MEAE), the Fyssen Foundation and a Seal of Excellence (SoE) Fellowship from the Austrian Academy of Sciences (OEAW).







 Open access
Open access