Introduction
Ilex paraguariensis A.St.-Hil. – Aquifoliaceae, known as yerba mate, is a tree species, native, widely consumed and cultivated in Brazil, Paraguay and Argentina (Cardozo Junior and Morand, Reference Cardozo Junior and Morand2016; Mateos et al., Reference Mateos, Baeza, Sarria and Bravo2018); this species has been recognized by its ecological, economic, social and cultural importance in southern Brazil (Croge et al., Reference Croge, Cuquel and Pintro2021; Duarte et al., Reference Duarte, Gabira, Tomasi, Amano, Nogueira and Wendling2022). Traditionally, yerba mate leaves are used in the preparation of tonic and stimulating drinks, such as chimarrão, tereré and mate tea (Cardozo Junior and Morand, Reference Cardozo Junior and Morand2016; Croge et al., Reference Croge, Cuquel and Pintro2021; Duarte et al., Reference Duarte, Gabira, Tomasi, Amano, Nogueira and Wendling2022).
Yerba mate leaves have more than 200 compounds (Melo et al., Reference Melo, Marques, Wendling, Kopka, Erban and Hansel2020); among these, we highlight the methylxanthines (caffeine and theobromine), saponins, minerals and caffeoylquinic acids, with numerous benefits to human health (Cardozo Junior and Morand, Reference Cardozo Junior and Morand2016; Bracesco, Reference Bracesco2019; Valduga et al., Reference Valduga, Gonçalves, Magri and Finzer2019). Caffeine and caffeoylquinic acids are particularly relevant to product development for certain consumer groups and markets, such as energy drinks, decaffeinated infusions or industrial extraction of compounds (Duarte et al., Reference Duarte, Tomasi, Helm, Amano, Lazzarotto, Godoy, Nogueira and Wendling2020).
The current demand and potential use of the species initiated a new growth cycle for the yerba mate economy on which plantations must meet the expectations of productivity and quality standards demanded by the world market (Duarte et al., Reference Duarte, Mireski, Oliszeski, Wendling and Stuepp2019). Therefore, the selection of genotypes with specific chemical characteristics and high productivity has been increasingly relevant for new plantations (Stuepp et al., Reference Stuepp, Bitencourt, Wendling, Koehler and Zuffellato-Ribas2017; Duarte et al., Reference Duarte, Mireski, Oliszeski, Wendling and Stuepp2019). For this purpose, the cuttings must have a high quality, especially with a root system well-formed for plant establishment in the field.
The literature demonstrates that yerba mate vegetative propagation is promising; however, the root formation and the cutting's quality are strongly related to the genotype to be propagated (Kratz et al., Reference Kratz, Pires, Stuepp and Wendling2015; Santin et al., Reference Santin, Wendling, Benedetti, Morandi and Domingos2015; Duarte et al., Reference Duarte, Mireski, Oliszeski, Wendling and Stuepp2019; Gazzana et al., Reference Gazzana, Pimentel, Lohmann, Maculan and Bisognin2020). Thus, for the multiplication of these materials, rhizogenic potential of each genotype – i.e. the ability to form adventitious roots – must be considered (Gazzana et al., Reference Gazzana, Pimentel, Lohmann, Maculan and Bisognin2020). The success of rooting is also related to other factors such as age, sex of parent plants (Wendling and Santin, Reference Wendling and Santin2015) and environment (Hartmann et al., Reference Hartmann, Kerster, Davies and Geneve2014); among these, seasonality is an important factor to be studied, because it influences directly on the rooting process (Stuepp et al., Reference Stuepp, Bitencourt, Wendling, Koehler and Zuffellato-Ribas2015; Sá et al., Reference Sá, Portes, Wendling and Zuffellato-Ribas2018) and on the chemical composition of yerba mate (Coelho et al., Reference Coelho, Athayde and Schenkel2001; Schubert et al., Reference Schubert, Zanin, Pereira and Athayde2006). Despite the numerous chemical compounds found in yerba mate leaves, little is known about their variation over the year in different genotypes and their influence on rhizogenic process. Hence, the genotype selection with a particular chemical profile, coupled with cloning techniques, allows new plantations to provide raw material with high potential, resulting in selected products and higher added value (Wendling, Reference Wendling2004; Duarte et al., Reference Duarte, Tomasi, Helm, Amano, Lazzarotto, Godoy, Nogueira and Wendling2020).
Thus, we aimed to evaluate seasonal variation in cuttings rooting and in bioactive compound contents of I. paraguariensis genotypes and the influence of these chemical compounds on the rhizogenic process. Our study is the broadest found in the literature so far, characterizing 19 yerba mate genotypes in the four seasons of the year, contributing as a new and important step of the yerba mate breeding programme.
Materials and methods
Selection and harvest of plant material
We studied nineteen 23-years-old genotypes, established in a provenance and progenies trial stablished in Ivaí, Paraná, Brazil (Wendling et al., Reference Wendling, Sturion, Reis, Stuepp and Peña2016). Genotypes were selected based on previous studies on productivity and bioactive compound contents (Helm et al., Reference Helm, Ruiz, Hansel, Stuepp and Wendling2015; Wendling et al., Reference Wendling, Sturion, Reis, Stuepp and Peña2016; Schuhli et al., Reference Schuhli, Penteado Junior and Wendling2019). The genotypes were submitted to pruning every 2 years, keeping the crown lowered and providing more juvenile propagules. The plant material for vegetative propagation and bioactive compound evaluation was collected at the end of four seasons of the year, between September 2018 and June 2019.
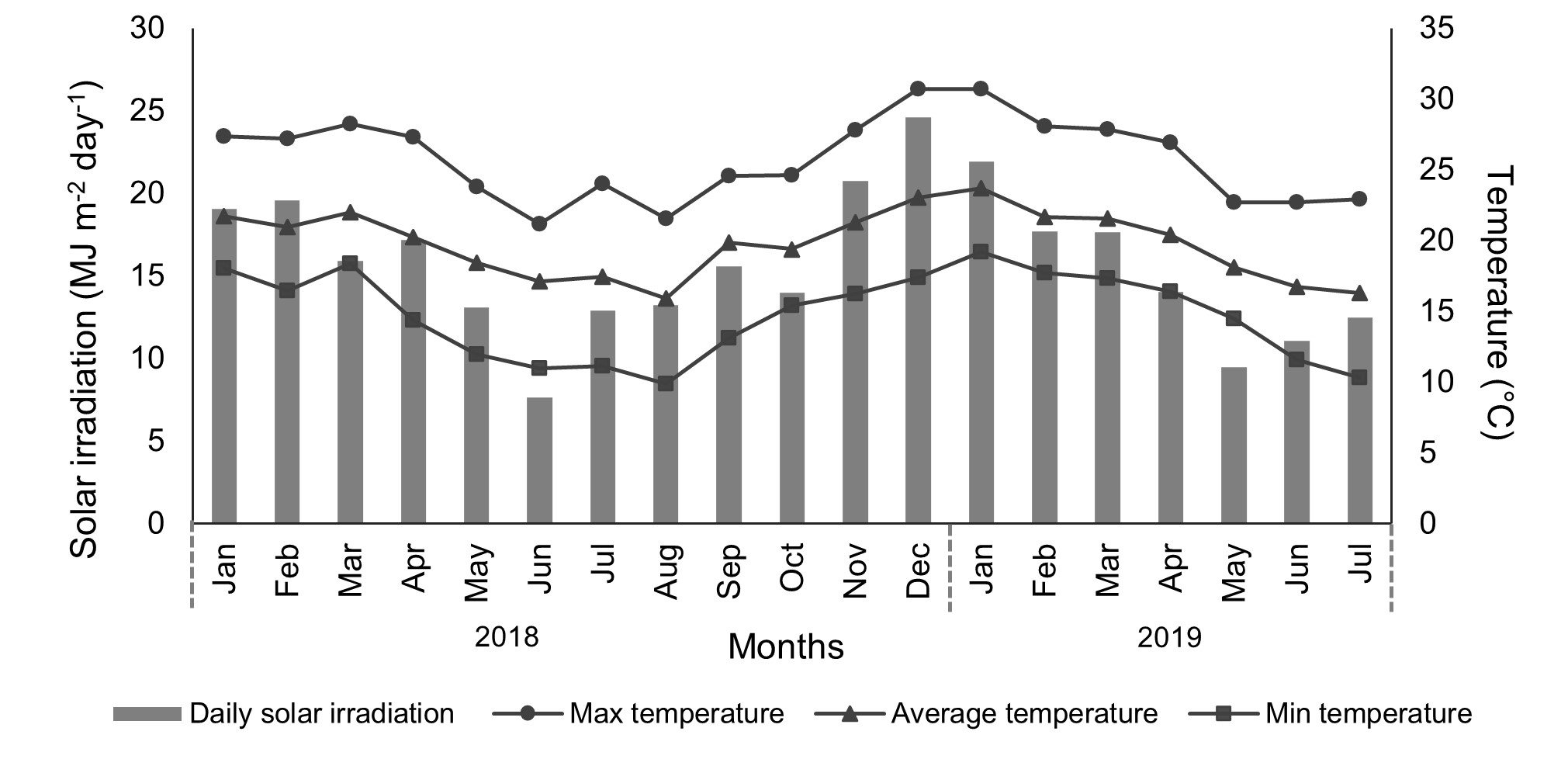
Daily solar irradiation and average, maximum and minimum temperatures data were obtained from meteorological station of Ivaí recorded by the National Institute of Meteorology – INMET (online Supplementary Fig. S1).
Cuttings rooting
We collected semi-woody shoots of the current year from the crown (average height of 1.5 m) from each 19-stock plant. Collections were carried out in the morning and propagules were packed in polystyrene boxes, moistened and transported to the place of cutting preparation. Cuttings were prepared to a length of 12 ± 1 cm, keeping two leaves reduced by half. They were disinfected with sodium hypochlorite (0.5%) for 10 min, washed in running water for 5 min and then immersed in fungicide Bendazol® (1 ml/l) for 10 min. The base of each cutting was treated with a hydroalcoholic solution (50% v/v) of 6000 mg/l of indole-3-butyric acid (IBA) for 10 s.
Cuttings were planted in polyethylene boxes (14.5 × 43.5 × 34.0 cm – height, width and depth, respectively) filled with medium granulometry vermiculite as a substrate. The boxes were kept in a greenhouse with intermittent fogging for 15 s every 6 to 10 min (relative humidity above 80% and temperature between 20 and 30°C). After 100 days in the rooting bed, the rooting percentage (cuttings with roots at least 2 mm in length), the number of roots per cutting, the leaf retention (cuttings that kept the original leaves) and the sprouting percentage (new sprouts of at least 5 mm) were measured. Five replicates of 10 cuttings per genotype were evaluated in each season, in a completely randomized design.
Bioactive compounds
Extract preparation
In each season, about 200 g of ripe mature leaves free of injuries were collected directly around the circumference from the crown of each of the 19 genotypes (performed at the same time as the harvests for cuttings experiments). The leaves were packed in kraft bags and transported in thermal boxes to the laboratory and stored in an ultra-freezer (−80°C) until the analysis was carried out. The leaves were lyophilized at −40°C for 72 h and crushed to obtain a fine and homogeneous powder. For aqueous extract, 10 mg of sample was added 2 ml of ultrapure water and vortexed for 30 s. The extractions were performed in a thermomixer equipment at 60°C and 300 rpm for one hour and then filtered through a 0.22 μm nylon filter. Mass correction to the chemical analysis was performed by the moisture content (evaluated in triplicate by drying in an oven at 105°C until constant weight) (Duarte et al., Reference Duarte, Tomasi, Helm, Amano, Lazzarotto, Godoy, Nogueira and Wendling2020).
Methylxanthines and caffeoylquinic acids
The methylxanthines and caffeoylquinic acids were determined using high performance liquid chromatography (HPLC – Shimadzu®). For analyses, 20 μl of aqueous extract was injected onto a Shim-Pack CLC-ODS (M) C18 column (250.0 × 4.6 mm i.d., 5 μm particle size), protected by a Shim-Pack CLC G-ODS pre-column (100.0 × 4.0 mm i.d.), both Shimadzu®. The separation of compounds present in the aqueous extract was conducted at 30°C with a flow rate of 0.5 ml/min, with mobile phase A eluent (H2O: acetic acid – 99.9:0.1 v/v) and phase B eluent (acetonitrile 100%). The wavelength used to detect the compound was 280 nm (fixed). The gradient elution programme was (minutes): 0–15 (3% B), 15–20 (3–20% B), 20–40 (20% B), 40–45 (20–30% B), 45–55 (30–100% B), 55–75 (100% B), 75–80 (100–3% B) and 80–95 (3% B) (Duarte et al., Reference Duarte, Tomasi, Helm, Amano, Lazzarotto, Godoy, Nogueira and Wendling2020).
The identification and quantification of methylxanthines caffeine (1,3,7-trimethylxanthine) and theobromine (3,7-dimethylxanthine) were performed from analytical curves obtained with standards, between concentrations 0 to 1.0 mg/ml (r 2 = 0.9933) and from 0 to 0.5 mg/ml (r 2 = 0.9827), respectively. The identification of monocaffeoylquinic acids (3-caffeoiliquinic acid (CQA3), 4-caffeoylquinic acid (CQA4), 5-caffeoylquinic acid (CQA5)) was also performed with standards and semi-quantification was obtained from an analytical curve between concentrations of 0 and 10 mg/ml of CQA3 (r 2 = 0.9914) (Duarte et al., Reference Duarte, Tomasi, Helm, Amano, Lazzarotto, Godoy, Nogueira and Wendling2020). The results were expressed in mg of compound per gram of dry sample (mg/g). The analysis was carried out in triplicates for the 19 genotypes over four seasons.
Statistical analysis
Pearson's correlation (P < 0.05) was performed to verify the relationships between rooting variables and bioactive compounds for the 19 genotypes. Genotypes with rooting equal to or above 70% in at least one season of the year were submitted to subsequent statistical analyses; that is a satisfactory cutting rooted percentage for yerba mate (Stuepp et al., Reference Stuepp, Bitencourt, Wendling, Koehler and Zuffellato-Ribas2017; Duarte et al., Reference Duarte, Mireski, Oliszeski, Wendling and Stuepp2019). Rooting variables showed non-normal distribution (Shapiro Wilk test, P < 0.05) and non-homogeneous variances (Bartlett test, P < 0.05), being analysed by generalized linear models with Gaussian distribution and averages compared by multiple comparisons by Tukey test (P < 0.05), considering the factors genotypes and seasons. For methylxanthines and monocaffeoylquinic acids contents, we used multivariate statistics by principal component analysis (PCA) to determine the compounds with the greatest contribution to the results. The analysed compounds were evaluated for normal distribution and homogeneous variances using analysis of variance and Tukey test (P < 0.05).
Results
Pearson's correlation matrix (Fig. 1) demonstrated no strong correlation between two groups of variables studied (rooting and bioactive compounds); however, there are significant correlations between variables in each group. Rooting percentage was positively correlated with number of roots, leaf retention and new sprouts (r = 0.7). Chemical compounds showed significant correlations between them, especially among caffeoylquinic acid isomers.

Figure 1. Pearson's correlation between cuttings rooting (R), number of roots (NR), leaf retention (LR), new sprouts (NS), caffeine (C), theobromine (T), 3-caffeoylquinic acid (CQA3), 4-caffeoylquinic acid (CQA4) and 5-caffeoylquinic acid (CQA5) for 19 Ilex paraguariensis genotypes. *Significant at P < 0.05.
There was variation for rooting results between and within genotypes in all periods (Fig. 2). Rooting percentage ranged from 0 to 98%. Genotypes EC18, EC26, EC32 and EC33 obtained the lowest rooting means, not reaching 30% in any of the evaluations. Only EC21 genotype had more than 70% rooting in all evaluated seasons. For subsequent analysis, we used just genotypes with rooting above 70% in at least one of the seasons, totalizing eight genotypes.

Figure 2. Rooting percentage (average and standard deviation) throughout the four seasons of the year in 19 I. paraguariensis genotypes.
For rooting variables, there was a significant interaction between the factors for seasons of the year and genotypes (online Supplementary Table S1). EC21 genotype was the only one that did not differ in rooting percentage according to seasons, which was above 82% in all seasons (Table 1). For other genotypes, summer and/or spring showed highest rooting percentages. In general, autumn resulted in lowest mean root number, and spring provided cuttings with a greater number of roots, except for EC28 and EC45. For sprouting percentage and leaf retention, summer stood out with highest means. For these two variables, genotypes did not differ in spring and summer seasons, except for EC53 and EC40.
Table 1. Rooting percentage, number of roots per cutting, leaf retention percentage and new sprouts percentage in cuttings of eight genotypes of Ilex paraguariensis in four seasons of the year

Capital letters for comparison between genotypes and lowercase letters for comparison between the seasons.
The PCA performed with contents of methylxanthines and monocaffeoylquinic acids indicated that CQA5 presented the highest contribution for principal component 1, and caffeine for principal component 2 (Fig. 3). Analysing distribution by groups, we observed that spring data all shifted to the right, towards CQA5, indicating a trend towards greater accumulation of this compound in this season (Fig. 4a). Performing PCA by groups of genotypes, we found a clear separation of EC22, located in the opposite quadrant of caffeine, indicating that this genotype has low levels of this compound (Fig. 4b). The EC21 and EC22 genotypes were in the right graph quadrant, possibly with high levels of CQA5 (Fig. 4b).

Figure 3. Principal component analysis (PCA) for contents of methylxanthines and monocaffeoylquinic acids.

Figure 4. Principal component analysis (PCA) for groups of seasons of the year (a) and genotypes (b). The ellipses were defined by a 95% confidence interval.
Compounds with the greatest contribution to PCA were caffeine and CQA5; thus, we evaluated these compounds in eight genotypes and throughout four seasons of the year. There was a significant interaction between factors for both compounds (online Supplementary Table S2). As indicated by PCA, spring showed highest levels of CQA5 (Fig. 5a); this behaviour was observed in six of eight genotypes. Only for EC21 and EC22 genotypes, spring did not differ significantly from other seasons. EC21 presented highest means of CQA5 between evaluated genotypes in every season, except in spring. In winter and autumn, EC22 genotype did not differ significantly from EC21, with both showing high levels of CQA5. These results confirm the trend of highest levels of CQA5 for EC21 and EC22 genotypes indicated by PCA. EC65 genotype showed low levels in all seasons, except on spring.

Figure 5. 5-caffeoylquinic acid (a) and caffeine (b) content (dry basis) for eight I. paraguariensis genotypes over four seasons of the year, compared for Tukey test (P < 0.05). Capital letters for comparison between genotypes and lowercase letters for comparison between the seasons.
Mean caffeine contents ranged from 0.02 (EC22) to 34.48 mg/g (EC53), both in autumn (Fig. 5b). EC22 genotype showed low levels of caffeine in all seasons, differing significantly from others. The genotype with the highest caffeine content in winter was EC40; for spring and autumn, EC53 genotype was superior. In summer, genotypes EC24, EC40 and EC53 showed the highest levels of this compound. We also observed that all genotypes showed significant variations in caffeine levels throughout the year, however, there is no clear trend according to the season.
Discussion
Rooting variables did not present stable correlations with bioactive compounds, indicating that compound contents probably did not influence on rhizogenic process of yerba mate genotypes. Some studies indicate that phenolic compounds can promote or inhibit the rooting process (Rout, Reference Rout2006; Porfirio et al., Reference Porfirio, Calado, Noceda, Cabrita, Silva, Azadie and Peixe2016; Shang et al., Reference Shang, Wang, He, He, Liu and Fu2017) which is dependent on the species; in our founds, phenolics did not influence the rooting, showing null correlation. Sá (Reference Sá2018) obtained similar results for yerba mate minicuttings.
Contrarily, Vieira et al. (Reference Vieira, Maggioni, Tomasi, Gomes, Wendling, Helm, Koehler and Zuffellato-Ribas2021) found a negative correlation between total phenolic compounds and rooting percentage on yerba mate minicuttings and recommended carrying out quantitative analyses of each compound separately, which we did in the present work. Thus, we can state that monocaffeoylquinic acids, which is a phenolic compound, had no negative influence on the rooting of yerba mate cuttings; but this result does not exclude the possibility that other phenolic compounds present in the species, such as dicaffeoylquinic acids and others monophenols, may influence the rhizogenic process (Shang et al., Reference Shang, Wang, He, He, Liu and Fu2017; Sá, Reference Sá2018).
Different phenolic compounds, such as chlorogenic acid and caffeic acid found in yerba-mate, can interfere in adventitious rooting by acting as rooting co-factors during the root formation process (Krajnc et al., Reference Krajnc, Turinek and Ivančič2013) and protecting rooting-inducer endogenous auxin indole-3-acetic acid (IAA) from being destructed by peroxidases (Shiozaki et al., Reference Shiozaki, Makibuchi and Ogata2013; Hartmann et al., Reference Hartmann, Kerster, Davies and Geneve2014). Besides, our results indicate that methylxanthines and monocaffeoylquinic levels did not influence the rooting of yerba mate by cuttings, and the low rooting observed in some genotypes was not associated with these bioactive compounds. Thereby, the null correlation between chemical compounds and rhizogenesis can be considered a positive aspect of yerba mate silviculture because we can select genotypes with different compound contents and propagate them vegetatively.
The positive correlation between rooting percentage and leaf retention demonstrated that the yerba mate rhizogenic process is dependent of presence of leaves, an observation already made in other studies (Sá et al., Reference Sá, Portes, Wendling and Zuffellato-Ribas2018; Vieira et al., Reference Vieira, Maggioni, Tomasi, Gomes, Wendling, Helm, Koehler and Zuffellato-Ribas2021). The rhizogenic process is favoured by presence and/or maintenance of leaves on cuttings, providing carbohydrates and phytohormones essential for rooting (Hartmann et al., Reference Hartmann, Kerster, Davies and Geneve2014). Thus, the early loss of leaves stops the transportation of energy and favourable substances (Hartmann et al., Reference Hartmann, Kerster, Davies and Geneve2014), which can be related to lower rooting rates observed in winter and autumn. We also observed strong positive correlations between rooting percentage and new sprouts. Physiologically, production of new shoots may be correlated with auxin/cytokinin balance, which when in adequate concentrations favour rooting (Li et al., Reference Li, Xue, Xu, Feng and An2009).
In our study, the season influenced rooting of yerba mate cuttings; this may be related to the consistency of shoots, carbohydrate concentration and endogenous regulatory substances (Hartmann et al., Reference Hartmann, Kerster, Davies and Geneve2014). There is a high variation in solar irradiation and in minimum temperature throughout the year in the region where stock plants are located. In periods of high temperature and solar irradiation – i.e. spring and summer – occur intense vegetative growth of the species (Rakocevic and Martim, Reference Rakocevic and Martim2011; Aguiar et al., Reference Aguiar, Gabira, Tomasi, Duarte, Vieira, Lavoranti and Wendling2022) and consequently, propagules are more herbaceous, which enhances rooting capacity.
In addition, even in a greenhouse, there is variation in microclimatic conditions; temperature and the quality and quantity of the irradiance during rooting process modulate auxin availability, transport and activity, affecting adventitious root development (Almeida et al., Reference Almeida, Aumond, Costa, Schwambach, Ruedell, Correa and Fett-Neto2017). For yerba mate, other authors also observed rooting variability as a function of seasonal variation (Stuepp et al., Reference Stuepp, Bitencourt, Wendling, Koehler and Zuffellato-Ribas2017; Sá et al., Reference Sá, Portes, Wendling and Zuffellato-Ribas2018; Pimentel et al., Reference Pimentel, Lencina, Kielse, Rodrigues, Somavilla and Bisognin2019; Vieira et al., Reference Vieira, Maggioni, Tomasi, Gomes, Wendling, Helm, Koehler and Zuffellato-Ribas2021), corroborating with our results; the rooting improvement in warmer months is an important indicator for the vegetative rescue of genotypes in the field by cuttings.
Rooting also proved to be genotype-dependent; so, each genotype must be treated individually to provide the necessary conditions for rooting and increase the quality of cuttings production (Vieira et al., Reference Vieira, Maggioni, Tomasi, Gomes, Wendling, Helm, Koehler and Zuffellato-Ribas2021). Some genotypes used in our experiment with high commercial interest, mainly based on their chemical composition, presented low rooting potential. In these cases, drastic pruning, total or branch cutting, annealing and serial grafting can be used as alternative rescuing techniques (Stuepp et al., Reference Stuepp, Bitencourt, Wendling, Koehler and Zuffellato-Ribas2015; Wendling and Santin, Reference Wendling and Santin2015), reestablishing physiological and biochemical conditions, thus improving the rooting capacity (Wendling et al., Reference Wendling, Trueman and Xavier2014).
Yerba mate is a natural source of phenolic compounds, especially chlorogenic acids and their isomers, compounds with high antioxidant activity (Riachi and De Maria, Reference Riachi and De Maria2017; Mateos et al., Reference Mateos, Baeza, Sarria and Bravo2018). In our results, the CQA5 varied between genotypes and seasons, and spring was the season with higher contents; EC21 and EC22 especially had high levels of CQA5 and are promising genotypes for industrial extraction of this compound. Furthermore, other studies demonstrate that variation in yerba mate chlorogenic acids can be related to leaf development and environmental conditions (Butiuk et al., Reference Butiuk, Martos, Adachi and Hours2016), and there is a tendency towards higher levels of these compounds in younger leaves (Blum-Silva et al., Reference Blum-Silva, Chaves, Schenkel, Coelho and Reginatto2015).
In general, higher caffeine contents in yerba mate have been observed in warmer periods and decrease in winter months (Coelho et al., Reference Coelho, Athayde and Schenkel2001; Schubert et al., Reference Schubert, Zanin, Pereira and Athayde2006; Duarte, Reference Duarte2020; Tomasi, Reference Tomasi2020). In our results, each genotype has shown different behaviour at each harvesting time. We observed caffeine content variation between seasons, but a stronger difference between genotypes; this fact can be attributed to caffeine high heritability in yerba mate plants (Cardozo Junior et al., Reference Cardozo Junior, Donaduzzi, Ferrarese-Filho, Friedrich, Gonela and Sturion2010). We highlight the EC22 genotype, which presented lower levels of caffeine in all harvests. This result demonstrates the possibility of individual selection with different contents of this compound and assists in adopting breeding strategies. Given the importance of caffeine contents for development of differentiated products (Valduga et al., Reference Valduga, Gonçalves, Magri and Finzer2019; Duarte et al., Reference Duarte, Tomasi, Helm, Amano, Lazzarotto, Godoy, Nogueira and Wendling2020), the selection of adequate genotypes for the development of markets and diverse products based on yerba mate is essential. Also, it is important to emphasize that the study evaluated the same genotypes at different times of the year regarding the content of leaves bioactive compounds, which is of great relevance for breeding programmes of the species.
Thus, we concluded that seasonality influenced the rhizogenesis of cuttings and bioactive compound contents. There was no correlation between chemical compounds and rhizogenesis of cuttings. Spring and summer provided higher percentages of rooting. The highest content of CQA5 was observed in spring; despite the significant variation in caffeine contents, genotypes did not show a clear trend according to the season. Besides, we consider this study essential to link genotype selection regarding bioactive compound contents to the maximum potential for vegetative propagation, indicating potential genotypes for breeding programmes. The scheduling of better harvest seasons for vegetative propagation or obtaining a particular chemical profile also gives crucial information for yerba mate producers.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S147926212300059X.
Acknowledgements
To Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the doctoral scholarships; To Chimarrão Bitumirim Indústria e Comércio de Erva-mate Ltda and Empresa Brasileira de Pesquisa Agropecuária (Embrapa Florestas) for technical and logistical support.