We consider the long-time behaviour of a West Nile virus (WNv) model consisting of a reaction–diffusion system with free boundaries. Such a model describes the spreading of WNv with the free boundary representing the expanding front of the infected region, which is a time-dependent interval  $[g(t), h(t)]$ in the model (Lin and Zhu, Spatial spreading model and dynamics of West Nile virus in birds and mosquitoes with free boundary. J. Math. Biol. 75, 1381–1409, 2017). The asymptotic spreading speed of the front has been determined in Wang et al. (Spreading speed for a West Nile virus model with free boundary. J. Math. Biol. 79, 433–466, 2019) by making use of the associated semi-wave solution, namely
$[g(t), h(t)]$ in the model (Lin and Zhu, Spatial spreading model and dynamics of West Nile virus in birds and mosquitoes with free boundary. J. Math. Biol. 75, 1381–1409, 2017). The asymptotic spreading speed of the front has been determined in Wang et al. (Spreading speed for a West Nile virus model with free boundary. J. Math. Biol. 79, 433–466, 2019) by making use of the associated semi-wave solution, namely  $\lim _{t\to \infty } h(t)/t=\lim _{t\to \infty }[\!-g(t)/t]=c_\nu$, with
$\lim _{t\to \infty } h(t)/t=\lim _{t\to \infty }[\!-g(t)/t]=c_\nu$, with  $c_\nu$ the speed of the semi-wave solution. In this paper, by employing new techniques, we significantly improve the estimate in Wang et al. (Spreading speed for a West Nile virus model with free boundary. J. Math. Biol. 79, 433–466, 2019): we show that
$c_\nu$ the speed of the semi-wave solution. In this paper, by employing new techniques, we significantly improve the estimate in Wang et al. (Spreading speed for a West Nile virus model with free boundary. J. Math. Biol. 79, 433–466, 2019): we show that  $h(t)-c_\nu t$ and
$h(t)-c_\nu t$ and  $g(t)+c_\nu t$ converge to some constants as
$g(t)+c_\nu t$ converge to some constants as  $t\to \infty$, and the solution of the model converges to the semi-wave solution. The results also apply to a wide class of analogous Ross–MacDonold epidemic models.
$t\to \infty$, and the solution of the model converges to the semi-wave solution. The results also apply to a wide class of analogous Ross–MacDonold epidemic models.
We focus on the population dynamics driven by two classes of truncated  $\alpha$-stable processes with Markovian switching. Almost necessary and sufficient conditions for the ergodicity of the proposed models are provided. Also, these results illustrate the impact on ergodicity and extinct conditions as the parameter
$\alpha$-stable processes with Markovian switching. Almost necessary and sufficient conditions for the ergodicity of the proposed models are provided. Also, these results illustrate the impact on ergodicity and extinct conditions as the parameter  $\alpha$ tends to 2.
$\alpha$ tends to 2.
In this paper, we study the existence of a solution for the following class of non-local problems:
P
where N ≥ 3, λ > 0, γ ∈ [1, 2), f : ℝ → ℝ is a positive continuous function and K : ℝN × ℝN → ℝ is a non-negative function. The functions f and K satisfy some conditions that permit us to use bifurcation theory to prove the existence of a solution for (P).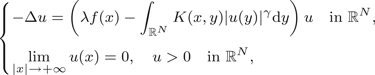 $$\eqalign{\big\{&-\Delta u=\left(\lambda f(x)-\int_{{open R}^N}K(x,y)\vert u(y)\vert ^{\gamma}\hbox{d}y\right)u\quad \mbox{in } \R^{N}, \cr &\lim_{\vert x\vert \to +\infty}u(x)=0,\quad u \gt 0 \quad \text{in } {open R}^{N},}$$
$$\eqalign{\big\{&-\Delta u=\left(\lambda f(x)-\int_{{open R}^N}K(x,y)\vert u(y)\vert ^{\gamma}\hbox{d}y\right)u\quad \mbox{in } \R^{N}, \cr &\lim_{\vert x\vert \to +\infty}u(x)=0,\quad u \gt 0 \quad \text{in } {open R}^{N},}$$

