
We construct two new classes of topological dynamical systems; one is a factor of a one-sided shift of finite type while the second is a factor of the two-sided shift. The data are a finite graph which presents the shift of finite type, a second finite directed graph and a pair of embeddings of it into the first, satisfying certain conditions. The factor is then obtained from a simple idea based on binary expansion of real numbers. In both cases, we construct natural metrics on the factors and, in the second case, this makes the system a Smale space, in the sense of Ruelle. We compute various algebraic invariants for these systems, including the homology for Smale space developed by the author and the K-theory of various  $C^{*}$-algebras associated to them, in terms of the pair of original graphs.
$C^{*}$-algebras associated to them, in terms of the pair of original graphs.

Let $G$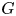 be a compact connected simple Lie group of type $(n_{1},\,\ldots,\,n_{l})$
be a compact connected simple Lie group of type $(n_{1},\,\ldots,\,n_{l})$ , where $n_{1}<\cdots < n_{l}$
, where $n_{1}<\cdots < n_{l}$ . Let $\mathcal {G}_k$
. Let $\mathcal {G}_k$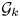 be the gauge group of the principal $G$
be the gauge group of the principal $G$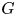 -bundle over $S^{4}$
-bundle over $S^{4}$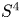 corresponding to $k\in \pi _3(G)\cong \mathbb {Z}$
corresponding to $k\in \pi _3(G)\cong \mathbb {Z}$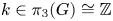 . We calculate the mod-$p$
. We calculate the mod-$p$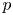 homology of the classifying space $B\mathcal {G}_k$
homology of the classifying space $B\mathcal {G}_k$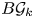 provided that $n_{l}< p-1$
provided that $n_{l}< p-1$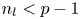 .
.


