1 Introduction
1.1 With or Without the Idea of Organic Progress?
The word ‘progress’ comes from the Latin word pro-gradi, meaning to walk forward. In English, the term ‘progress’ was, until the sixteenth century, used in the axiologically neutral sense of walking, that is, travel and movement forward in space. However, starting from the seventeenth century, the word gained an additional meaning that is still in use today: change has a positive value, referring to something leading towards a better, more elevated state (Simpson and Wiener Reference Simpson and Wiener1989). Thus, in the notion of progress, there is both a descriptive element (change in time) and an axiological element – from the Greek word axios, valued – related to what is considered to be ‘good’ and ‘better’ (Goudge Reference Goudge1961).
The idea of progress has several domains of application. Applied to human civilization as a whole, the idea was central to the Enlightenment movement of the late seventeenth and eighteenth centuries, before seeing a decline during the twentieth century (Wagar Reference Wagar1972). The idea of progress can also be applied more specifically to the technological domain (change towards better technical artefacts) and to the scientific domain (change towards better scientific theories). Another field of application of the idea of progress is the organic domain, where this change towards the ‘better’ refers to the evolutionary history of living beings. The application of the idea of progress to the organic domain raises questions about the complex relationship between organic progress and evolutionary theory, which is the main focus of this Element.
The historian of science John C. Greene sums up this question nicely: ‘Evolutionary biologists, it seems, can neither live with nor live without the idea of progress’ (Greene Reference Greene1990). The reasons why evolutionary biologists should not ‘live with’ organic progress are quite easy to understand. At first glance, ‘progress’ sounds like a non-scientific term: the axiological element seems to imply a subjective judgement, which should have no place in a scientific discipline. Indeed, in the past, organic progress underpinned a clearly anthropocentric view: humanity was considered the ‘best’ organism at the top of the progressive evolution (Ruse Reference 82Ruse1996), as in the representation of the ‘chain of being’ by Charles Bonnet (Bonnet Reference Bonnet1745).Footnote 1
The Darwinian revolution, however, should have once and for all rejected the anthropocentric perspective: Homo sapiens is just one branch of the tree of life, rather than the final product of the ‘chain of being’. Thus, we may think that detachment from the ‘men at the top’ perspective should also entail the idea of organic progress being abandoned starting with Darwin. Yet the history of biological thought shows that this is not the case. In his book Monad to Man: The Concept of Progress in Evolutionary Biology, Michael Ruse shows that the idea is present in Darwin’s thought and in the work of the authors of the Modern Synthesis across the twentieth century (Ruse Reference 82Ruse1996).
It is thus pertinent to ask why evolutionary biologists seem unable to ‘live without’ some idea of organic progress. Several options can be considered:
Because of some anthropocentric bias. There may be some influence of metaphysical remnants of the idea that man is at the top of the chain of being (despite the detachment that we should have today from these beliefs).
Because of some cultural bias. For example, the idea of ‘optimization’ as applied in human occidental societies (e.g., to production or technical artefacts) could lead biologists to overestimate the relevance of this notion as applied to organisms – see for example the arguments in Hamant (Reference Hamant2022).
Because there are good theoretical arguments that some ‘change towards the better’ is legitimate within evolutionary theory, for example, some conceptual implication of the theory. In that case, it should be possible to clearly identify these arguments and clarify this notion of ‘change towards the better’.
In contemporary evolutionary biology, the mainstream position seems to be that there are no good theoretical arguments for organic progress (Desmond Reference Desmond2021): I refer to the intuition against organic progress.Footnote 2 This intuition is often related to the arguments of Stephen Jay Gould, the main critic of organic progress in the twentieth century.
However, there are also some renowned biologists who have an intuition for a notion of organic progress, that is, believing there are good theoretical arguments in favour of a progress notion (e.g., George Gaylord Simpson, Richard Dawkins, and Geerat Vermeij). Evolutionary theory would conceptually imply something that can be labelled as a ‘change towards the better’. This discussion has been reopened recently and is ongoing (Rosenberg and McShea Reference Rosenberg and McShea2008, Desmond Reference Desmond2021).
This Element contributes to the debate by focussing on two main questions:
1) There is no unanimity on discarding organic progress from evolutionary biology. Why? Are there any theoretical arguments for using the normative terms ‘good’ and ‘better’ within evolutionary theory?
2) If there are, can we clarify the idea that some ‘change towards the better’ would be conceptually implied by evolutionary theory?
Since the conceptual question of organic progress is elusive, I will begin by presenting a tripartite distinction, before explaining the strategy to tackle these two main questions.
1.2 Cause, Criterion and Scope of Progress
This distinction between cause, criterion and scope of progress is inspired by the historian Warren Wagar (Reference Wagar1972), who proposed it for the progress of human civilization: the belief that human civilization has moved, is moving and will move in a desirable direction (Bury Reference Bury1920). Wagar distinguishes three key elements when considering the progress of civilization: agency, content and subject of progress (Wagar Reference Wagar1972, pp. 4–5). ‘Agency’ is the cause of this improvement, for example divine providence, collective human effort or ‘great men’. ‘Content’ is the criterion by which we judge whether there has been an improvement, for example material health, knowledge or wealth. What Wagar calls ‘subject’ corresponds to the extent or scope of progress, for example the whole cosmos, humanity or a single nation.
Let us now consider organic progress: the idea that during the history of life, there has been a change towards organic forms that are, in some respects, ‘better’ than preceding forms. Here it is possible to apply the distinction proposed by Wagar and raise three corresponding questions:
Cause: What is driving the improvement? As we will see, the mechanism of natural selection is generally considered by evolutionary biologists to be the cause of organic progress.
Criterion: Which properties are we referring to when talking about progress? It could be argued that complexity, intelligence or perception of the environment have improved during the history of life: these are criteria by which to judge a possible improvement. It could also be argued that ‘something’ is improving, but it is not necessarily related to a single property.
Scope: To which ‘range of life’ does the improvement apply? For example, organic progress might concern life as a whole (thereby having a global scope), or only certain specific lineages (thereby having a local scope). Thus, the scope concerns the ‘framing’ of organic progress in time and space.
1.3 Structure of the Element
In this Element, I start by tackling the question of natural selection as a potential cause of organic progress. In Section 2, I present the positions of two authors who have opposed intuition on this issue: Charles Darwin and Stephen Jay Gould. While Darwin’s intuition is that the mechanism of natural selection may cause some kind of change towards the better, Gould has devoted several works to claiming that the notion of progress has no legitimacy within evolutionary theory. Gould (Reference Gould1996) is well known for arguing that, at a global scale, the increase in complexity across evolutionary history might be due to a stochastic process rather than natural selection (for Gould’s argument of the ‘left wall’, see Section 8). Then, in Section 2, I present relevant arguments put forward by Gould against organic progress. In particular, I focus on the argument that any possible ‘change towards the better’ can, across evolution, be destroyed by mass extinctions (Gould Reference Gould1985, Reference Gould1989).
In Section 3, I consider Darwin’s intuition on organic progress and discuss whether it can be made more precise concerning the criterion and scope. I do so by drawing on Richard Dawkins. For Dawkins, the concept of adaptation by natural selection conceptually implies organic progress on small and medium timescales. This notion only allows us restricted comparisons between ancestors and descendants within a lineage. Even if this progress might be destroyed, Dawkins argues that it still counts as a betterment in the organisms’ ‘equipment for survival’. Although Dawkins offers more precise arguments for organic progress than Darwin, a significant objection remains unaddressed: are ‘normative’ terms (good, better) acceptable in the context of a scientific theory?
Section 4 is devoted to the previous question. Drawing on Rosenberg and McShea (Reference Rosenberg and McShea2008), I claim that there are theoretical arguments for defending a specific kind of instrumental value in evolutionary biology, which can be called ‘organic value’. Organic value has nothing to do with moral issues or with the notion of ‘intrinsic value’. It just implies the notion of relative value with respect to a means: ‘X is instrumentally valuable for the purpose of P’. After presenting this framework, I examine an objection to organic value: can we speak of purposes for organic entities other than human beings? Countering this objection, I claim that purposes can be understood with reference to the concept of ‘function’. I further argue that this concept implies an axiological polarity, while having a proper and legitimate place within biology. I answer with an affirmative to the first question (Are there any theoretical arguments for using the normative terms ‘good’ and ‘better’ within evolutionary theory?), provided that normative terms are clearly understood as referring to organic value.
I then move to question two: can we further clarify Dawkins’ notion of local progress? I tackle this question in successive steps. In Section 5, I present two empirical cases from the fossil record for which there is a strong intuition that something is getting better. In Section 6, I draw on these two cases to propose a refinement of Dawkins’ notion. Based on this, I propose that adaptation conceptually implies that ‘something gets better’ in this specific sense: in the adaptive process, organic traits in a population get better with respect to a biological function, while the condition of ‘all other things being equal’ holds. I label this notion ‘Functional Improvement of Organic Traits’ (FIOT). This analysis leads me to propose a new way of looking at some classical issues in philosophy of biology, such as the relationship between organic structures, functions and the environment. Once the notion is refined, I consider whether there is more direct evidence for FIOT by looking at experimental evolution. In Section 7, I examine the results of the Long-Term Evolutionary Experiment by Richard Lenski and interpret these results based on my account.
In Section 8, it is finally time to turn the discussion towards organic progress on a global scale. Despite Gould’s powerful criticism related to mass extinctions, I ask whether there may be good theoretical arguments to defend global progress. I start by clarifying what macroevolutionary trends are and that only natural selection allows us to speak of progressive trends. After briefly presenting the main candidates for progressive trends, I focus on complexity and I show, following the palaeobiologist Daniel McShea, that it is not a tenable criterion. This analysis allows me to highlight the requirements for a satisfactory criterion for global progress and thereby try another direction of inquiry. Following an insight of the French philosopher Georges Canguilhem, and based on the recent framework of Bourrat, Deaven et al. (Reference Bourrat, Deaven and Villegas2024), I suggest that robustness of adaptedness and evolvability are promising criteria for global progress, at least from a theoretical standpoint.
2 Rival Intuitions about Organic Progress
2.1 Darwin’s Intuition on ‘Competitive Highness’
Darwin’s attitude towards the question of organic progress is not straightforward: the father of evolutionary theory continually struggled with this ‘inextricable’ subject (Darwin Reference Darwin1858) and indeed he was quite embarrassed by it. The literature provides several interpretations of Darwin’s attitude; here I mainly follow the reconstructions of Ospovat (Reference Ospovat1981), Radick (Reference Radick2000) and Shanahan (Reference Shanahan2004). The main point that I focus on is that Darwin tried to characterize what linked the mechanism of natural selection with some kind of organic improvement, setting out for the first time the idea of progress in terms of competitive advantage.
On several occasions, Darwin wonders what naturalists mean by ‘higher’ and ‘lower’ organisms. In a passage in On the Origin of Species, he says that naturalists have not yet defined to each other’s satisfaction what is meant by high and low forms (Darwin Reference Darwin1859, p. 337). However, he adds, there seems to be one sense in which it is possible to talk about ‘higher’ organic forms:
But in one particular sense the more recent forms must, on my theory, be higher than the more ancient; for each new species is formed by having had some advantage in the struggle for life over other and preceding forms. If under a nearly similar climate, the Eocene inhabitants of one quarter of the world were put into competition with the existing inhabitants of the same or some other quarter, the Eocene fauna or flora would certainly be beaten and exterminated; as would a secondary fauna by an Eocene, and a palæozoic fauna by a secondary fauna.
Following the logic of his theory, the more recent forms have beaten the preceding forms in the struggle for survival, and thus they are ‘in one particular sense’ higher. In Darwin’s thought experiment, if Eocene forms were placed in competition with present forms, the latter would exterminate the former. It is relevant to note here the importance of the condition ‘under a nearly similar climate’, referring to the supposition of some constancy of the environment (see further discussion that follows). He concludes:
I do not doubt that this process of improvement has affected in a marked and sensible manner the organisation of the more recent and victorious forms of life, in comparison with the ancient and beaten forms; but I can see no way of testing this sort of progress.
Darwin believes that a kind of ‘improvement’ has taken place, but he acknowledges that this kind of progress is impossible to test as we do not have a ‘time machine’ to perform the experiment. Indeed, the word ‘improvement’ is present in the definition of natural selection itself from the second edition of On the Origin of Species:
This principle of preservation, I have called, for the sake of brevity, Natural Selection; and it leads to the improvement of each creature in relation to its organic and inorganic conditions of life.
In the basic Darwinian scheme, natural selection is the result of (1) organisms presenting variations (rendering them more or less fitted to their life conditions) and (2) a limitation of resources (Malthusian principle). Some organisms are fitter than others in relation to life conditions and, resources being limited, they therefore have a competitive advantage. This mechanism takes place over time. Let us consider, in a ‘quarter of the world’, an abstract organic community at time 1 and at time 1000. Can the organic community at t1000 be considered as somehow ‘better’ than the more ancient? Or, rather, should we say that the organic community t1000 is adapted to its life conditions exactly as the community at t1 was adapted to their own life conditions?
Darwin seems to think (despite the acknowledged difficulty of empirically testing) that the recent organic community should be considered somehow ‘better’. In the attempt to justify this position, he introduces the expression ‘competitive highness’ in a letter to his colleague and friend Joseph Dalton Hooker (Darwin Reference Darwin1858). With this, Darwin refers to forms whose superiority is expressed by the fact that they have won in the competition with other forms. This is true of recent forms compared to ancient forms in the same habitat, as in the thought experiment cited earlier. This is also true, for Darwin, for species inhabiting a very large area when compared to species in small areas. As explained to Hooker, Darwin believes that the larger the area, the larger the number of organic forms that are present and can interact, resulting in more severe competition (Darwin Reference Darwin1858). Organic forms that have withstood more severe competition are ‘superior’: the plants of Eurasia (large area) are the most ‘improved’ and thus are able to withstand the ‘less perfected’ Australian plants (Australia being a smaller area, with less intense competition).
It should be noted that, in Darwin’s thought experiment, the criterion ‘under a nearly similar climate’ is crucial. The intuitive idea is that, if the change in ‘life conditions’ is too dramatic between t1 and t1000, the axiological comparison between organisms cannot have any meaning: we should rather think that the t1000 organic community is adapted to its life conditions exactly as the t1 community was adapted to its own life conditions. The ‘life conditions’ of an organism may be inorganic (e.g., temperature and light exposure), or organic (e.g., symbionts, parasites, competing species, and predators). With the expression ‘under a nearly similar climate’, Darwin seems to presuppose a relative constancy of inorganic conditions of life. Inorganic conditions would be somehow ‘fixed’ and selective pressure mostly related to competition between organisms. Following Darwin’s reasoning, t1000 organic forms may be thought to be competitively better than those at t1, where ‘better’ is relative to the fixed abiotic conditions. However, it is unclear which kind of organic forms Darwin is considering here: is he comparing a whole extant organic community in a quarter of the world (e.g., the whole extant Australian fauna) with a whole Eocene organic community (e.g., the whole Eocene Australian fauna)? Before returning to the problems in Darwin’s reasoning exposed so far, it is important to note that ‘competitive highness’ is not the only notion of organic progress that is considered by Darwin, as will emerge in the next section.
2.2 Darwin on ‘Competitive Highness’ and Complexity
From the third edition of Origin (Darwin Reference Darwin1861), Darwin elucidates another sense in which it can be said that one organic form is superior to another: the morphological differentiation of parts and specialization to different functions. This is generally what is meant when biologists speak about ‘complexity’. Darwin explicitly refers to Karl Ernst Von Baer and Henri Milne-Edwards:
Von Baer’s standard seems the most widely applicable and the best, namely, the amount of differentiation of the different parts (in the adult state, as I should be inclined to add) and their specialisation for different functions; or, as Milne Edwards would express it, the completeness of the division of physiological labour.
Thus, there are two senses of progress in Darwin’s reasoning, which are clarified in a letter to Hooker (Darwin Reference Darwin1858). ‘Highness in the common acceptation of the word’ refers to complexity, while ‘competitive highness’ is related to success in the struggle for life and might also be related to ‘degradation of organization’. Darwin takes the example of the blind, degraded, worm-like snake that might supplant more complex forms of earthworms: the example shows that morphologically simple organisms can be competitively ‘better’ than complex ones.
Yet while Darwin clearly sees that the two senses of ‘highness’ can be in principle decoupled, he seems to claim that they usually go hand in hand. This claim depends on accepting a premise: that complexity is generally ‘an advantage to each being’ (Darwin Reference Darwin1861, p. 364). Once this premise is accepted, it can be affirmed that modern forms should be ‘higher’ than ancient forms with respect to both competitive success and morphological complexity. The ‘coupling’ of the two senses of ‘highness’ is clear in this passage of the third edition of Origin:
As the specialisation of parts and organs is an advantage to each being [complexity], so natural selection will constantly tend thus to render the organisation of each being more specialised and perfect, and in this sense higher; not but that it may and will leave many creatures with simple and unimproved structures fitted for simple conditions of life, and in some cases will even degrade or simplify the organisation, yet leaving such degraded beings better fitted for their new walks of life. In another and more general manner [competitive highness], we can see that on the theory of natural selection the more recent forms will tend to be higher than their progenitors; for each new species is formed by having had some advantage in the struggle for life over other and preceding forms. (…) So that by this fundamental test of victory in the battle for life [competitive highness], as well as by the standard of the specialisation of organs [complexity], modern forms ought on the theory of natural selection to stand higher than ancient forms.
It is interesting to note that, after this passage, Darwin raises the question: ‘is this the case?’ In the same edition from 1861, he writes:
Is this the case? A large majority of palaeontologists would certainly answer in the affirmative; but in my judgment I can, after having read the discussions on this subject by Lyell, and Hooker’s views in regard to plants, concur only to a limited extent. Nevertheless it may be anticipated that the evidence will be rendered more decisive by future geological research.
Darwin then changes this passage in the successive editions of Origin and seems to have increasing confidence in the reasoning. In 1866, he writes that he cannot look at the conclusion ‘as fully proved, though highly probable’ (Darwin Reference Darwin1866, p. 402). Three years later, he supposes that ‘the answer must be admitted as true, though difficult of full proof’ (Darwin Reference Darwin1869, p. 410).
Here I have briefly exposed Darwin’s reasoning about organic progress, which rests on two senses of ‘highness’: competitive highness and complexity. For my argument, I prefer to start by focussing on Darwin’s idea of competitive highness decoupled from the notion of complexity, which I will treat in Section 8. For the moment, let us focus on competitive highness and consider the opposite intuition about progress, put forward by Stephen Jay Gould.
2.3 Gould’s Intuition against Organic Progress
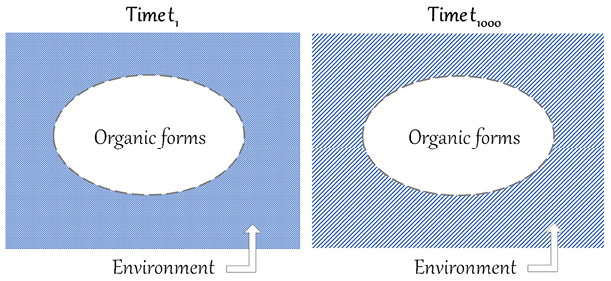
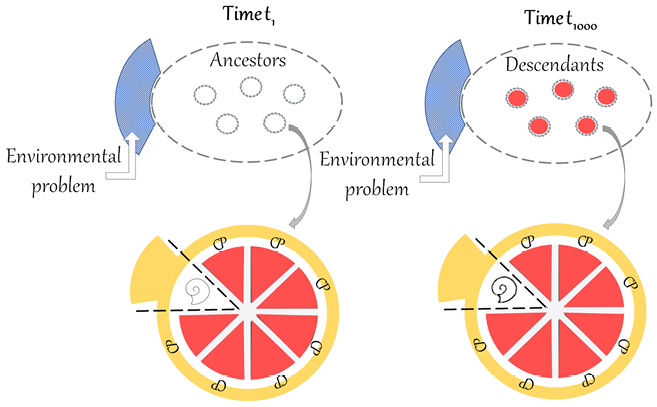
The American palaeontologist Stephen Jay Gould has questioned Darwin’s reasoning on organic progress. The core of his criticism can be summed up by saying that natural selection is a mechanism that results in organisms that are adapted to changing local environments. Environments change in a non-directional and unpredictable manner, in line with the principles of Charles Lyell’s geology. Organisms that are not adapted to the new environments are eliminated, and organisms that stay alive are better adapted to local conditions, which can change again and again. In this ‘continuous dance of local adjustment’ (Gould Reference Gould2002, p. 468), the fitness of each organic form is always relative to particular conditions. Let us imagine that we can schematically represent organic forms with ovals and their environment with a rectangle enveloping them, and let us suppose that we can therefore compare the situation at time t1 with the situation at time t1000, as represented in Figure 1.
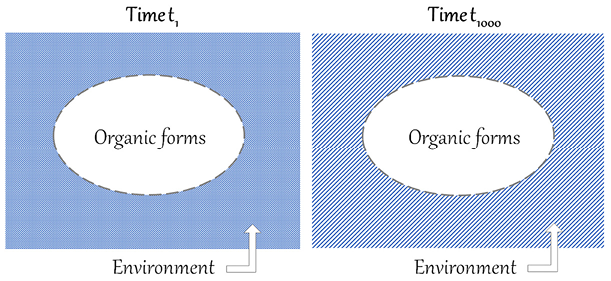
Figure 1 Comparison of t1 and t1000, illustrating Gould’s criticism of the incommensurability of organic forms in heterogenous environments. The oval represents organic forms and the rectangle the environment, which is heterogenous from t1 to t1000.
If the rectangle at t1000 is completely heterogenous from the rectangle at t1 (as represented by the different patterns), there is no common background against which organic forms can be compared. If the two situations are incommensurable, it is not legitimate to claim that recent forms are better than ancient ones. With the core of this criticism in mind, let us now examine Gould’s arguments against progress, starting with his texts from the 1980s.
2.4 The Paradox of the First Tier
The theory of punctuated equilibria was first formulated by Gould and Niles Eldredge in Reference Eldredge and Gould1972 (Eldredge and Gould Reference Eldredge and Gould1972) and further developed in the following years (Gould and Eldredge Reference Gould and Eldredge1977, Gould Reference Gould1982). The ways in which this theory dismantles the intuition for progress are made explicit in a text published by Gould in 1985, ‘The Paradox of the First Tier: An Agenda for Paleobiology’ (Gould Reference Gould1985). In this text, Gould proposes considering evolutionary time as structured following three levels or ‘tiers’.
The first tier includes evolutionary events ‘of the ecological moment’ (Gould Reference Gould1985, p. 2): this relates to very short timescales, generally referred to as microevolution.
The second tier refers to ‘trends within lineages and clades that occur during millions of years in ‘normal’ geological time between events of mass extinctions’ (Gould Reference Gould1985, pp. 2–3). For this tier, it is possible to use the term macroevolution.
The third tier is represented by the phenomenon of mass extinctions, whose importance had recently been discovered by palaeobiologists at the time when Gould was writing in 1985. Sometimes the term megaevolution is used to refer to this level (Huneman Reference Huneman, Huneman and Bouton2017).
Gould suggests that the Darwinian tradition – and namely the architects of the Modern Synthesis – deny this hierarchical structure of time by extrapolating from the first tier to the second and third tier. The extrapolation rests on the assumption that causal processes of the first tier (the mechanism of natural selection) can be used to explain what happens at the other tiers. Gould argues that this extrapolation is impossible because each tier has its own causal processes. Palaeobiology must be conceived as an autonomous discipline that studies the second and third tier, decoupling them from the first.
But in what way does this concern progress? Gould claims that the intuition for organic progress is related to the first tier. Darwin expects competitive highness because he is convinced that competition is a main driver of evolutionary change and because he believes that a biomechanical improvement is a way in which organisms adapt to local environments (Gould Reference Gould1985, p. 4). The paradox of the first tier can be described as follows: despite the fact that in theory we should expect organic progress (thus, despite Darwin’s intuition of competitive highness), no clear vector of cumulative progress can be found at larger timescales. In Gould (Reference Gould1985), two solutions are considered to solve this paradox, which I will label respectively ‘no theoretical reasons for progress at the first tier’ and ‘progress is reversed at the second and third tier’.
No theoretical reasons for progress at the first tier. At the short timescales related to the first tier, there are no good theoretical reasons to think that natural selection might cause progress. Darwin’s intuition has no internal foundation in evolutionary theory (it rather comes from a cultural bias).
Progress is reversed at the second and third tier. The theoretical justification for organic progress is valid at the first tier: at short timescales, natural selection may cause something that can meaningfully be called progress. However, this progress is reversed (diluted, undone) by specific processes taking place at the second and, more significantly, third tier.
Gould (Reference Gould1985) explicitly states that he wants to defend the second solution, even though he seems to be sympathetic to the first. Let us consider the two arguments by which Gould argues for the second solution, one relating to the macroevolutionary level, the other to the megaevolutionary level.
The first argument refers to the theory of punctuated equilibria. It is a central theme in Gould’s work that the importance of natural selection has been overestimated by the Darwinian tradition: ‘immediate adaptations of organisms within populations are not the only stuff of long-term evolution’ (Gould Reference Gould1985, p. 6). Gould’s claim is that at the macroevolutionary level there are some distinct causal processes, for example, species selection. The basic idea is that species are not sorted because of competitive success, but rather for properties at the species level, like propensity towards speciation. These complex issues, which I can only evoke here, are still debated today in palaeobiology (Vrba and Lieberman Reference Vrba and Lieberman2005).
The second argument is, as Gould himself admits, the most important one for rejecting the idea of organic progress (Gould Reference Gould1985, p. 7). The phenomenon of mass extinction was well documented from 1980, with the publication of the article ‘Extraterrestrial Cause for the Cretaceous-Tertiary Extinction’ by Alvarez, Alvarez et al. (Reference Alvarez, Alvarez, Asaro and Michel1980). This new evidence supported the view that mass extinctions are frequent and rapid phenomena, qualitatively different in rates and effect from other phenomena of the history of life. From this perspective, extinctions are not always competitive replacements, as in a gradualist view of evolution; when mass extinctions occur, species survive or die by chance. In a catastrophic event like the Earth being hit by a meteorite, progress (as defined by adaptations to local environments) is of no utility for organisms.
2.5 Wheels and Wedges
Four years after the publication of ‘The Paradox of the First Tier: An Agenda for Paleobiology’, Gould returned to the question of the relationship between tiers of time and progress in the paper ‘The Wheel of Fortune and the Wedge of Progress’ (Gould Reference Gould1989). Here he insists still further on the megaevolutionary argument: mass extinctions are ‘the wheel of fortune’ that destroy ‘the wedge of progress’. The reference here is to the Darwinian metaphor of the wedge (Stauffer Reference Stauffer1975, p. 208) that would justify the idea of competitive highness. However, Gould’s position with respect to the macroevolutionary argument seems to be different in this paper. Gould appears here more sympathetic to the idea that a few cases of progressive trends can be observed at the macroevolutionary level. This appears clearly in the following passage:
I am persuaded that some cases in Darwin’s preferred mode of organic competition have been documented. Biologist Geerat Vermeij (…), for example, has demonstrated a geological trend for thicker and stronger crab claws matched by ever more efficient defenses (spines, knobs, and thick shells) in the snails that crabs love to eat. I accept the interpretation of this lock-step escalation as an ‘arms race.’
Here Gould refers to the researches of his colleague Vermeij and to his book Evolution and Escalation (Vermeij Reference Vermeij1987). By studying the fossil documentation in marine environments, Vermeij describes an ‘arms race’ between gastropods (prey) and shell-crushing crustaceans (predator). Gould admits that this example can count as a progressive trend, but he argues that we should not over-extrapolate from this because ‘one swallow does not make a spring’:
But (…) a case or two in the fossil record does not establish a pattern. Directional trends produced by wedging do occur, but they scarcely cry for recognition from every quarry and hillslope. The overwhelming majority of paleontological trends tell no obvious story of conquest in competition.
Thus, although a few progressive trends may exist, they do not represent a common pattern. And, even if they exist, mass extinctions could always end them. The history of life is thus reminiscent of the destiny of Sisyphus. Punished in Hades, the king of Corinth is forced to eternally roll a painfully heavy stone up a hill (moment of the wedge of progress through competition), the stone slipping back to the bottom every time he reaches the top (moment of the wheel of fortune that reverses progress) (Gould Reference Gould1989, p. 20).
2.6 No Progress in the Bare Bone Mechanics of Darwinian Theory
Gould returns relentlessly to the issue of progress in two of his books: his widely read Full House: The Spread of Excellence from Plato to Darwin, and in his opus magnum, The Structure of Evolutionary Theory (Gould Reference Gould2002). As Full House is rather focussed on complexity as a criterion for global progress – an issue that I will discuss in Section 8 – I will here focus on his arguments in the 2002 book and in particular on the interpretation that Gould gives of Darwin’s thought.
According to Gould’s interpretation, Darwin does not have any theoretical basis for defending his reasoning about progress. Darwin ‘stuck’ to progress solely for cultural reasons, related to the importance of the idea (especially when applied to human civilization) in the Victorian society (Gould Reference Gould2002, ch. 6). To do so, Darwin added some supplementary hypotheses to the ‘bare bones mechanics’ of his theory (Gould Reference Gould2002, p. 467). The two hypotheses mutually reinforce each other and can be summed up following the reconstruction of Radick (Reference Radick2000):
Selective factors related to the biotic environments are the most relevant ones in evolutionary processes when compared to causes related to abiotic environments. Here Gould refers to the metaphor of ‘wedges’ used by Darwin (Gould Reference Gould2002, ch. 6).
Independently of the oscillations related to abiotic factors, the Earth is more and more ‘crowded’ over time, thus competition between living forms intensifies.
Two main disagreements can be spotted between Darwin and Gould. First, for Gould, the Darwinian supposition of a stable abiotic environment is unacceptable. For Gould, the history of life undergoes drastic changes in abiotic environments, and species extinctions are mainly related to these events rather than to competitive replacements, as Darwin believes. Second, concerning biotic factors, Gould is sceptical about the idea that competition represents a major phenomenon in evolution. He cites, for example, the ideas of the Russian naturalist and anarchist Peter Kropotkin, according to whom environmental pressures favour intra-specific cooperation rather than competition (Gould Reference Gould2002, p. 471). For Gould, Darwin’s insistence on competition (the intensity of which would be directly proportional to the degree of kinship between individuals) does not have a scientific foundation. It would rather be related to an illegitimate extra-scientific influence connected with societal beliefs.
It should be noted that, in this interpretation, the solution to the paradox of the first tier seems to be ‘no theoretical reasons for progress at the first tier’, rather than ‘progress is reversed or undone at the second and third tier’. In fact, Gould argues here that Darwin’s reference to biotic competition as a main factor in evolutionary processes has an extra-scientific rationale rather than one that is ‘internal’ to evolutionary theory. This would mean that, even at short timescales, the mechanism of natural selection does not cause something that can be called progress.
Considering the different texts, we can say that Gould seems to oscillate between two positions. On the one hand, Gould (Reference Gould1985) and Gould (Reference Gould1989) favour the position ‘progress is reversed or undone at the second and third tier’ (even admitting in 1989 the possibility of some cases of progress at the second tier). On the other hand, although not explicitly argued, the position that there are ‘no theoretical reasons for progress at the first tier’ seems implicit in the interpretation of Darwin’s reasoning in Gould (Reference Gould2002).
2.7 Weighing Up the Two Rival Intuitions
What should we think about Gould’s intuition against organic progress? Should we accept it and consider Darwin’s reasoning on competitive highness as entirely unsound on a theoretical basis? In order to weigh up the two rival intuitions, I will make three remarks as follows.
First, it seems that the most convincing argument proposed by Gould is the one concerning mass extinctions. Since the first proposals on this in the 1980s (Alvarez, Alvarez et al. Reference Alvarez, Alvarez, Asaro and Michel1980), the significance of these phenomena for the history of life has not been questioned. If mass extinctions regularly affect the history of life, then organic traits locally improved by natural selection are useless at the moment of the catastrophe. Hence, with the mass extinctions argument, Gould has shown that it is not possible to argue for organic progress having a global scope (concerning the whole history of life). In the same vein, Huneman (Reference Huneman, Huneman and Bouton2017) has recently suggested that the argument of mass extinctions is related to the contingency thesis (Beatty Reference Beatty1994), proposing that the randomness implied in mass extinctions goes against the possibility of extrapolating from microevolution to macroevolution and megaevolution (‘extrapolationist’ thesis, Huneman Reference Huneman, Huneman and Bouton2017). To sum up, I provisionally concede to Gould that, because of mass extinctions it is not straightforward to argue for progress at a global scale (but see Section 8 for a proposal of a promising candidate for global progress).
Second, a major question is opened by Gould’s reasoning. If organic progress having a global scope seems untenable, what about progress having a local scope, concerning short timescales? After all, we could consider that an improvement, even though it is undone at a later time, would still significantly count as an improvement in a given temporal interval. To modify Gould’s metaphor, we could think that Sisyphus has somehow advanced into a ramp phase, although his efforts are then subsequently nullified.
Third, Darwin’s intuition seems to rely on the fact that natural selection conceptually implies some kind of improvement. However, as formulated by Darwin, the reasoning for this raises various difficulties. As already mentioned, it is unclear which kind of organic forms Darwin is considering in the comparison between extant and ancient forms. In fact, it seems quite odd to compare a whole extant organic community (e.g., the whole extant Australian fauna) with a whole Eocene organic community (e.g., the whole Eocene Australian fauna). If this is what Darwin means, this is subject to the following objection: the extant organic community is adapted to its life conditions just as the ancient organic community was adapted to their own life conditions. As they are incommensurable, it is impossible to say that the recent one is better than the ancient. This leads us back to the problem raised with Figure 1. Is postulating the clause ‘under a nearly similar climate’ sufficient for defending his position?
In view of these three remarks, my proposal is as follows. Let us provisionally concede to Gould that, because of mass extinctions, it is not straightforward to argue for progress at a global scale. I thus suspend the judgement – applying a sort of sceptical epoché – and return to this issue in Section 8. For now, I propose giving Darwin’s intuition a chance, at least as far as organic progress with a local scope is concerned. Although Darwin’s reasoning presents several difficulties, let us consider the idea that drives his intuition: evolutionary theory conceptually implies that – under some circumstances – some kind of organic improvement can occur. Further, I attempt to give a clear and sound meaning to this idea as far as small timescales are concerned, drawing on the ideas of Richard Dawkins and then refining his notion.
3 Dawkins’ Account of Local Progress
3.1 Giving Darwin’s Intuition a Chance
To give Darwin’s intuition a chance, at least two issues need to be clarified.
First, clarification is needed concerning the scope of progress. What does local progress mean exactly? On the one hand, we could ask whether it is legitimate to compare Eocene forms with recent forms, as Darwin does in his thought experiment. This question refers to the restriction of temporal scale. On the other hand, we could ask which organisms can be compared at t1 and t1000. For example, are we making intra-specific comparisons, for example, comparing ancestors and descendants within the same lineage? Or are we rather referring to inter-specific comparisons, which would allow us to claim that a particular organic form X is better than a very different organic form Y?
Second, the criterion (or criteria) of improvement needs to be clarified. Admitting that ‘better’ organisms have a competitive advantage, do we think that this advantage is related to any specific organic property (e.g., complexity or intelligence) or might it be related to different properties? In this section, I try to clarify these issues by drawing on the ideas of Dawkins.
Richard Dawkins is a British biologist and ethologist. Along with Gould, he significantly contributed to popularizing evolutionary theory in the twentieth century. While the two biologists are on the same side in the defence of evolutionary theory with respect to creationism, they disagree on several major points regarding the interpretation of Darwinism (Shanahan Reference Shanahan and Delisle2017). After the publication of Full House: The Spread of Excellence from Plato to Darwin (Gould Reference Gould1996), Dawkins wrote a review of Gould’s book, titled ‘Human Chauvinism’ (Dawkins Reference Dawkins1997). Dawkins states that Gould’s definition of progress in this book is a human-chauvinistic one, which makes it ‘too easy to deny progress in evolution’ (Dawkins Reference Dawkins1997, p. 1016). In fact, Gould’s treatment of progress in this large-audience book is quite different from the arguments that I have outlined in the previous section. In Full House, Gould mainly refers to progress as a tendency to increase relative to some criterion (e.g., anatomical complexity and neurological elaboration) concocted by human beings to place themselves ‘atop a supposed heap’ (Gould Reference Gould1996, p. 20). Dawkins proposes an alternative definition of progress, which he claims to be ‘less anthropocentric’ and ‘more biologically sensible’ (Dawkins Reference Dawkins1997, p. 1016); I will examine this further.
3.2 The Adaptationist Criterion of Local Progress
First, Dawkins suggests that the criterion for organic progress should not be related to the same property for all lineages, like for example complexity of intelligence. Improvements could occur in different lineages, but they might not be related to the same property in all of them. As Dawkins states:
This [my adaptationist definition of progress] (…) takes progress to mean an increase, not in complexity, intelligence or some other anthropocentric value, but in the accumulating number of features contributing towards whatever adaptation the lineage in question exemplifies.
Hence, for Dawkins, there is no single property related to Darwinian ‘competitive highness’. Organisms can accumulate ‘features’ that contribute to adaptation, but these do not have to be the same for the different lineages. Dawkins’ intuition presents some similarities with Darwin’s intuition. In both cases, the biotic aspect of selection is considered to be predominant over the abiotic aspect: Dawkins refers to the idea of an ‘arms race’ occurring among organisms (Dawkins and Krebs Reference Dawkins and Krebs1979). However, Dawkins also adds some important points to Darwin, which I think can be summed up as three main points:
The ‘equipment for survival’ gets better.
Comparisons are limited to ancestors-descendants within a lineage.
Organic progress takes place from short to medium term.
Let us start with the first point about the equipment for survival: Dawkins’ claim is not that more recent organisms ‘survive better’. As Dawkins puts it:
The participants in the race do not necessarily survive more successfully as time goes by – their “partners” in the coevolutionary spiral see to that (the familiar Red Queen Effect). But the equipment for survival, on both sides, is improving as judged by engineering criteria. In hard-fought examples we may notice a progressive shift in resources from other parts of the animal’s economy to service the arms race (Dawkins and Krebs Reference Dawkins and Krebs1979). And in any case the improvement in equipment will normally be progressive.
For Dawkins, what gets better is the ‘equipment for survival’, as judged by ‘engineering criteria’.
Second, Dawkins clarifies the Darwinian thought experiment regarding progress. In fact, the father of evolutionary theory does not clearly state which kinds of organisms are considered in the comparison. An example by Dawkins suggests that the comparisons concern traits of the same lineage, for example, the eyes in vertebrates. In Dawkins’ example, the initial eyes of earlier vertebrate ancestors are compared with the better-performing eyes of vertebrate descendants (Dawkins Reference Dawkins1997, p. 1018). By considering this branch of the tree of life, we can say that an improvement took place relative to the function of vision. However, continuing Dawkins’ line of reasoning, it seems less straightforward to compare the vertebrate eyes and the invertebrate eyes (e.g., bivalves’ ocelli, structures for perceiving light).
Third, Dawkins is more explicit than Darwin about the temporal scale of the comparison: he admits that progress only makes sense in the short to medium term.
Coevolutionary arms races may last for millions of years but probably not hundreds of millions. Over the very long timescale, asteroids and other catastrophes bring evolution to a dead stop, major taxa and entire radiations go extinct (…) The several arms races between carnivorous dinosaurs and their prey were later mirrored by a succession of analogous arms races between carnivorous mammals and their prey. Each of these successive and separate arms races powered sequences of evolution which were progressive in my sense. But there was no global progress over the hundreds of millions of years, only a sawtooth succession of small progresses terminated by extinctions. Nonetheless, the ramp phase of each sawtooth was properly and significantly progressive.
Thus, Dawkins concedes that Gould is right in his argument that mass extinctions undo progress at long temporal scales. However, he claims that we can meaningfully speak of organic progress at short and medium timescales, taking adaptation as a criterion (Dawkins Reference Dawkins1997, p. 1016).Footnote 4
These three points make it clear that the notion of organic progress that Dawkins defends is narrow and only applicable at a local scale. I will refer to this notion as the Dawkins notion of local progress.
3.3 Clarifying Dawkins’ Notion of Local Progress
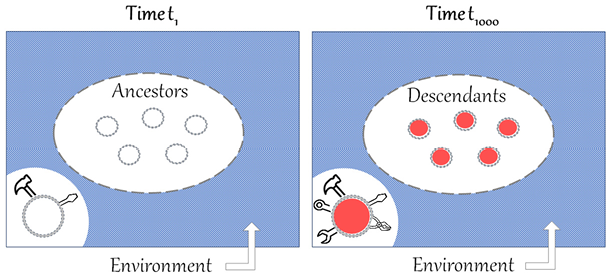
It can be useful to visualize Dawkins’ notion of local progress, as in Figure 2, and compare it to Figure 1.
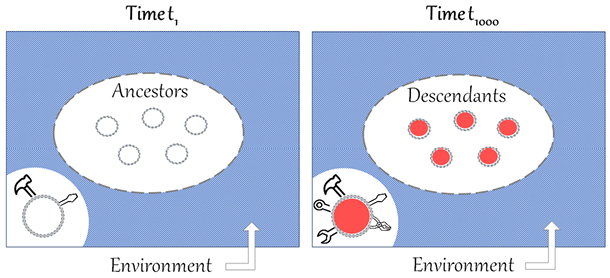
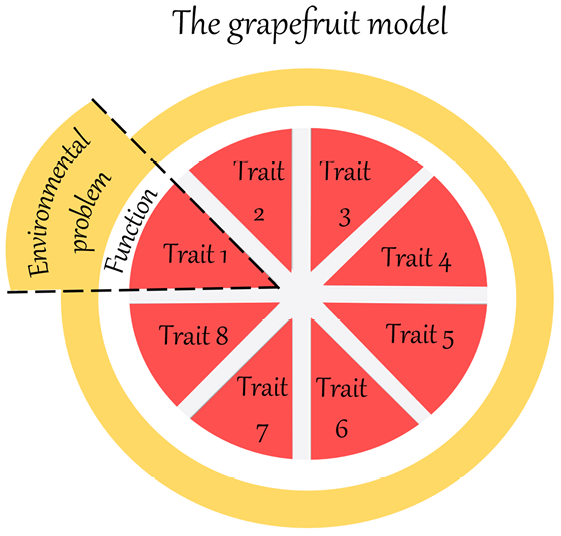
Figure 2 Comparison of t1 and t1000, illustrating Dawkins’ account of local progress. The ovals represent populations of ancestors and descendants of the same lineage, the circles represent individual organisms of the populations and the rectangles represent the environment. The environment is supposed to be homogenous because the timespan between t1 and t1000 only represents short to medium timescales. Due to competition, the population of descendants would present a better ‘equipment for survival’ than the population of ancestors, here represented by a richer ‘toolkit’.
First, in Figure 2, the meaning of the ovals is clarified. They no longer represent generic organic forms, but rather an ancestor population at t1 and a descendant population at t1000, belonging to the same lineage. I suggest further representing individual organisms of the two populations as circles within the ovals. Second, another important difference in Figure 2 is that the environment is supposed to be homogenous at t1 and t1000 because the timespan only represents short to medium timescales. Finally, for Dawkins, what differentiates organisms at the two different times is the ‘equipment for survival’. For the moment, let us visually represent this difference with a richer ‘toolbox’ in the descendant population (see Section 6 for further clarification).
It should be stressed that, for Dawkins, evolutionary theory conceptually implies local progress. His reasoning can be described as consisting of two premises and a conclusion:
P1: Evolutionary theory conceptually implies the possibility that the process of adaptation by natural selection takes place.
P2: The process of adaptation by natural selection conceptually implies local progress.
C: Evolutionary theory conceptually implies the possibility of local progress.
The first premise is commonly accepted among evolutionary biologists. But what kind of explanation do we have for the existence of specialised morphological structures? Dawkins has in mind here the kind of biological cases that had already surprised Darwin, where biological structures give a strong impression of ‘tight correspondence’ or ‘fine tuning’ with the surrounding environment, as with the vertebrate eyes, the echolocation in bats and dolphins, and the sophisticated adaptations of orchids (Dawkins Reference Dawkins1997, p. 1018). If these specialized morphological structures are our explanandum, the most plausible explanans is that the process of adaptation by natural selection took place. Otherwise, we should assume that these structures appeared suddenly after one large mutation, which seems counterintuitive (and leaves space for creationist objectionsFootnote 5). Dawkins claims that even Gould should admit this is the only possible explanans.
According to the second premise, the process of adaptation by natural selection conceptually implies that some improvement occurs. It does not matter if this improvement is local and is ‘undone’ at a long temporal scale; it can nonetheless be labelled improvement, thus legitimating discussions of progress. As Dawkins puts it, we should admit this adaptationist notion of progress ‘without stirring from our armchair’ (Dawkins Reference Dawkins1997, p. 1018).
By this definition, adaptive evolution is not just incidentally progressive, it is deeply, dyed-in-the-wool, indispensably progressive. It is fundamentally necessary that it should be progressive if Darwinian natural selection is to perform the explanatory role in our world view that we require of it, and that it alone can perform.
Yet, the second premise of the reasoning (P2) is much more controversial, as I will now explain.
3.4 An Unanswered Question in Dawkins’ Account
Dawkins presents some ways of making Darwin’s intuition about progress much more precise, that is, the three main points discussed earlier. However, his reasoning relies on the acceptance of the second premise, according to which the process of adaptation by natural selection conceptually implies local progress.
The controversial point is as follows: local progress refers to the notion of ‘betterment’, which in turn presupposes the notion of ‘good’. But are these normative terms acceptable within a scientific theory? Are we not supposed to remain in the factual or descriptive realm, instead of the normative realm? Defenders of the value-free ideal of science might raise the objection that these terms are unacceptable in a scientific discipline. They may claim that evolutionary biology should be purged from terms such as ‘progress’ (as well as ‘teleology’ and ‘goal-directedness’) because they could damage the professional image of the discipline (McShea Reference McShea2023). I will try to argue that, if the precise meaning of normative terms is made explicit, they do not represent a problem in this sense. The next section will be devoted to tackling this point.
4 Normative Terms within Evolutionary Biology
4.1 Rosenberg and McShea’s Framework
In philosophy, it is common to distinguish what is normative from what is non-normative (i.e., natural or descriptive). Normative terms include good, bad, right, wrong, and mandatory. By contrast, examples of non-normative terms are blue, red, round, small, big, light, and heavy. The domain of philosophy that is especially concerned with normative terms is moral philosophy, dealing with what is generally categorized as axiological (referring to values) or deontic (referring to norms, e.g., what is permitted and required). So how could there possibly be normative terms in evolutionary biology?
In the context of their discussion of organic progress, Rosenberg and McShea (Reference Rosenberg and McShea2008) provide a useful framework for understanding in what sense value attributions are relevant to evolutionary biology. The authors start by identifying two ways in which the normative component of organic progress could be understood:
Now there are two ways in which an increase in some feature of organisms might be valuable. The feature could be valuable to us, to human beings. Or it could be valued by the evolutionary process, so to speak, valued in the sense of preserved or enhanced owing to its adaptive value, the contribution it makes to survival and reproduction. The more progressive organism could be the one that is better at surviving and reproducing, the one that is more fit, so that progress would be an increase in whatever features of organisms underlie increased fitness, over the history of life.
The first option is to consider the feature in question as valuable for human beings. This is considered problematic by virtue of the value-free ideal: human values should not be relevant ‘to the findings of science or to the process of determining what is true’ (Rosenberg and McShea Reference Rosenberg and McShea2008, p. 129). Thus, the authors propose examining the second option, ‘valued in the sense of preserved or enhanced owing to its adaptive value’. Here they introduce the distinction between ‘intrinsic’ and ‘instrumental’ value. Typically, the distinction refers to something to which one could attribute a value ‘in itself’ (e.g., friendship, happiness), as opposed to something that is only valuable as a means to something else (e.g., money). The opposition between intrinsic and instrumental value is classically introduced while discussing moral issues, although it has been noted – mainly by Korsgaard (Reference Korsgaard1983) – that the distinction can be misleading.Footnote 6 Intrinsic value is an especially important concept in axiology, although the questions ‘what is intrinsic value’ and ‘what has intrinsic value’ are far from having simple and uncontested answers. Some authors have even raised the question of whether intrinsic value exists at all (Zimmerman and Bradley Reference Zimmerman, Bradley, Zalta and Nodelman2019, Section 2).
Rosenberg and McShea claim that the value referred to in evolutionary biology is instrumental. If one argues that one item is better than another for achieving a purpose, the term ‘better’ is being used instrumentally. This is first illustrated by a technical example: ‘an electron microscope is better than a light microscope for the purpose of seeing the very small details of certain objects’ (Rosenberg and McShea Reference Rosenberg and McShea2008, p. 129). The axiological term ‘better’ here refers to the specific purpose of seeing small details. This has no implication for the question of whether an electron microscope is valuable in itself, in absolute terms. For Rosenberg and McShea, this would be a matter of moral or ethical value.
Then, by analogy, the authors apply the same distinction to a biological case:
Switching to a biological example, the theory of natural selection identifies the thick coat of the polar bear as instrumentally valuable for insulation, and insulation as instrumentally good for survival and reproduction. But science stops at this point. It does not identify some further good for which survival and reproduction are instrumentally valuable. Nor does it suggest that survival and reproduction are intrinsically good in themselves. It is silent on this matter owing to its value neutrality.
The authors claim that statements about instrumental value are ‘essential’ in evolutionary biology, while they make no reference to intrinsic value. They specify that the difference between the two can be made by virtue of the fact that instrumental value is subject to empirical testing once we stipulate the criteria of goodness we have in mind (e.g., efficiency and speed). By contrast, there is no test or experiment that would enable us to settle disputes over intrinsic value (Rosenberg and McShea Reference Rosenberg and McShea2008, p. 130).
By analysing the example used by Rosenberg and McShea, we can identify two kinds of axiological statements (see De Cesare Reference De Cesare2019):
(i) A thick coat is instrumentally good for the polar bear to achieve the particular organic purpose of keeping its body warm.
(ii) Keeping the body warm is instrumentally good for the polar bear to achieve the general organic purpose of survival and reproduction.
By generalizing the example, we obtain:
(i) An organic trait (x) is instrumentally good for an organic entity (O) to achieve the particular organic purpose P.
(ii) The particular organic purpose (P) is instrumentally good for an organic entity (O) to achieve the general organic purpose of survival and reproduction (P*).
I suggest calling this kind of value ‘organic value’. This requires further clarification comprising two points, which are presented as follows.
4.2 Two Remarks Concerning the Framework
First, the attribution of the instrumental value of a means does not imply the value of the end itself. Referring to P*, Rosenberg and McShea explicitly state that there is no need to attribute value to survival and reproduction. In fact, science ‘does not identify some further good for which survival and reproduction are instrumentally valuable’ (Rosenberg and McShea Reference Rosenberg and McShea2008, p. 129). Indeed, this position would lead to the strong metaphysical thesis that ‘life’ and ‘reproducing life’ are better than death. However, it should be noted that this end P*, even if it does not need to be value-laden, has a particular status of being the ‘endpoint’ of instrumental value attributions. One could argue that there are good reasons why biologists are particularly interested in what contributes to organisms’ survival and reproduction. In fact, organisms that are good with respect to survival and reproduction are expected to be (except in the case of unlucky catastrophic events) those that leave descendants and ‘go through’ evolutionary history. To explain why extant organisms are there and present certain characteristics, we might refer to the fact that ancestors in the lineage must have been ‘good enough’ at surviving and reproducing.
Second, with organic value, we find ourselves in a specific part of the normative realm, where objective assessments about what is better or worse relative to a specific purpose are possible. Once again, the notion of organic value does not imply any ethical considerations. The distinction between the goodness that is implied in the case of organic value versus goodness ‘as an end’ might be clarified by drawing on an insight from Ludwig Wittgenstein. In his Lecture on Ethics, Wittgenstein (Reference Wittgenstein1929) presents two kinds of goodness and discusses what is valued as a means under the label of ‘relative good’. He describes it as follows:
If for instance I say that this is a good chair this means that the chair serves a certain predetermined purpose and the word good here has only meaning so far as this purpose has been previously fixed upon. In fact the word good in the relative sense simply means coming up to a certain predetermined standard. Thus when we say that this man is a good pianist we mean that he can play pieces of a certain degree of difficulty with a certain degree of dexterity. (…) Used in this way these expressions don’t present any difficult or deep problems.
However, Wittgenstein argues that the kind of ‘good’ that is interesting in ethics is not this relative good, but rather absolute good. For example, in the statement ‘this is the right way to Granchester’ the reference is to relative good. Instead, if we speak of the ‘right road’, this would be ‘the road which everybody on seeing it would, with logical necessity, have to go, or be ashamed for not going’ (Wittgenstein Reference Wittgenstein1929). The difference can be spotted because, contrary to judgements about absolute value, judgements about relative value can be translated in descriptive terms:
Every judgment of relative value is a mere statement of facts and can therefore be put in such a form that it loses all the appearance of a judgment of value: Instead of saying “This is the right way to Granchester,” I could equally well have said, “This is the right way you have to go if you want to get to Granchester in the shortest time”; “This man is a good runner” simply means that he runs a certain number of miles in a certain number of minutes, etc.
Wittgenstein’s idea is that it is possible to translate statements implying relative value into factual terms. I think that this insight nicely complements a point by Rosenberg and McShea concerning organic value, according to which it is possible to empirically test statements about instrumental value. In brief, it is possible – at least in principle – to objectively assess that one jaguar is better than another jaguar if I decide to focus on the activity of running fast. In the same way, it is possible to assess which is the best road to get to Granchester with respect to the purpose of reaching Granchester in the shortest time. Note that, in both cases, the result of the test might change if I ‘set’ the purpose differently: for example, for jaguars, by focussing on the most effective technique for capturing prey; or, for roads, reaching Granchester while enjoying the nicest view. However, once the purpose is clearly identified, the relative good in the statements becomes empirically testable.
Thus, this notion of relative good refers to a specific domain within the normative realm, where it is possible to assess that ‘x is better than y for P’ without appealing to subjective appreciations. Despite the use of normative terms, it is possible at least in principle to refer to something objective, for example, assess the better jaguar with respect to running fast, or assess the better road with respect to arriving in Granchester as soon as possible.
4.3 An Objection: Can We Speak of Means for a Purpose within Biology?
I have defended the idea that, with the framework of Rosenberg and McShea, we have a coherent notion of organic value. However, an objection might be raised here. As I have presented it so far, the notion of instrumental organic value speaks of good/bad means towards some ‘end’. However, the terminology referring to purposes within biology might be accused of being finalistic, and thus unacceptable. This objection, which could be labelled the anti-teleological objection to organic value, has some strong historical roots, which are worth sketching briefly, partly following Colin and Neal (Reference 77Colin, Neal and Zalta2020).
It is a cornerstone idea of modern science that final causes should be entirely banished. With respect to the study of living beings, this position comes in opposition to ancient authors such as Aristotle and Galen, for whom biological explanation cannot be purely mechanistic. For example, it is impossible to explain an organ (e.g., the heart) without considering its organic purpose (‘telos’) or function within the organism. However, these influential views were strongly challenged during the scientific revolution starting from the sixteenth century. The use of mechanical analogies to explain living entities has become dominant (e.g., in the work of Harvey and Descartes) and Francis Bacon clearly banished final causes from scientific methodology.
At the end of the eighteenth century, this issue was famously discussed in Immanuel Kant’s Critique of Judgment (Kant Reference Kant1790 [2000]). Kant claims that humans unavoidably understand living beings, with their unique capacity to grow and reproduce, as being purposive. Can this purposiveness be reduced to causal-mechanistic explanations? Commenting on Kant, Ginsborg (Reference Ginsborg, Zalta and Nodelman2022) nicely explains the intricacy of this issue:
The mechanical inexplicability of organisms leads to an apparent conflict, which Kant refers to as an ‘antinomy of judgment’, between two principles governing empirical scientific enquiry. On the one hand, we must seek to explain everything in nature in mechanical terms; on the other, some objects in nature resist mechanical explanation and we need to appeal to teleology in order to understand them (§70, 387).
Some authors have interpreted Kant’s position on teleology as denying the possibility of biology as a causal science,Footnote 7 but see Teufel (Reference Teufel, Smith and Nachtomy2013) for a criticism of this interpretation. It should also be noted that the debate on how to interpret Kant’s legacy on the issue of teleology and biology is far from closed. A recent paper by Gambarotto and Nahas (Reference Gambarotto and Nahas2022) presents the debate in the following terms. Kant claims that we need to appeal to teleology to understand organic entities. But should we consider teleological explanations simply as heuristic tools? Or should we rather think that purposiveness is an intrinsic feature of biological systems? Gambarotto and Nahas (Reference Gambarotto and Nahas2022) label these approaches as heuristic and naturalistic, respectively.Footnote 8
However, let us go back to the brief historical sketch, and particularly to the context into which Darwin builds his view of evolution. Although his direct knowledge of Kantian philosophy was not ‘necessarily great’ (Stowell Reference Stowell2024, p. 523), Darwin probably knew about Francis Bacon’s negative assessment of final causes via the British philosopher William Whewell (Ruse Reference Ruse1975). Darwin was aware that, in a Baconian perspective, the standards of scientific methodology set by modern physics would require the banishment of final causes. However, interestingly, Darwin sometimes refers to purposes and ‘final causes’, while admitting in a personal note: ‘it is anomaly in me to talk of Final Causes: consider this!’ (cited in Lennox Reference Lennox1993, p. 411).
Does Darwinian theory reject teleology or not? When Origin was published in 1859, Darwin’s contemporaries considered him either as a teleologist or a non-teleologist, either with praise or blame (Lennox Reference Lennox1995). It is well known that Darwin’s intention was to explain organic features without appealing to the design of a benevolent creator, who would have intentionally created a feature to fit a purpose. This kind of divine purpose, at the core of natural theology of William Paley, is banished by Darwin. Still, does this mean that Darwin barred all kinds of teleology from evolutionary theory? Scholars disagree on this point. Micheal Ghiselin answers with an affirmative, claiming that Darwin purged biology from teleology (Ghiselin Reference Ghiselin1994), while James G. Lennox defends the idea that Darwin ‘re-invented teleology’, a selection-based teleology (Lennox Reference Lennox1993).
Alongside these divergencies on the interpretation of Darwin’s thought, the contemporary debate about the legitimacy of teleological notions in evolutionary biology is far from closed (Colin and Neal Reference 77Colin, Neal and Zalta2020). One of the main things that bothers the ‘anti-teleologists’ is that referring to purposes would be anthropomorphic. As human beings are purposive, they would be led to attribute purpose where there is none (Colin and Neal Reference 77Colin, Neal and Zalta2020). Moreover, they claim that there is no need to appeal to purposes if we can explain biological organisms in the same way that we explain physical objects: by identifying causes and effects. Rather than saying that X achieves a purpose, we can simply say that X causes an effect.
After having outlined the historical roots of the anti-teleological objection, I will try to respond to the criticism. The aim is to show that the objection does not dismantle the notion of organic value that I have previously presented.
4.4 First Step to Answering the Objection: Clarifying Teleology within Biology
First, let us clarify what teleology within biology is not. It is possible to exclude from the outset any notions of intentional design, spirit or entelechy, which would carry a strong metaphysical element, widely rejected in modern science (McShea Reference McShea2023). Drawing on McShea’s reflection, it is possible to distinguish two ways in which teleology comes into play in biology:
Concerning the evolutionary process. This question is about large-scale evolutionary trends. At the scale of lineages, is there a sense in which evolution is somehow goal-directed? This is the main issue discussed by McShea (Reference McShea2023).
Concerning the results or outcomes of the evolutionary process, that is, characteristics of organismal structures or behaviours. Consider the example of a bird’s behaviour of migrating south for the winter. Is the bird’s behaviour goal-directed?
Orthogonal to this distinction, I propose identifying several attitudes towards teleology within biology:
Teleology eliminativism. In looking at both processes and outcomes, biology should be teleology-free and only deal with mechanisms.
Weak defence of teleology – heuristic approach. In the biological disciplines, from an epistemic perspective, we need teleological explanations. This position, corresponding to the heuristic interpretation of Kant on teleology (Gambarotto and Nahas Reference Gambarotto and Nahas2022), equates to saying that we need to appeal to teleology in our way of understanding the organic world. This might include processes and/or outcomes. However, this position would not necessarily imply the ontological affirmation according to which there is purposiveness outside human minds.
Strong defence of teleology – naturalistic approach. Biological objects, whether processes or outcomes, are ontologically teleological. This idea corresponds to what Gambarotto and Nahas (Reference Gambarotto and Nahas2022) call the naturalistic interpretation of Kant on teleology. This can be applied either to the processes (there is some kind of goal-directedness in large-scale evolution) or to results (there is some kind of goal-directedness in organic features, be this in structure, organization or behaviours). According to this view, there is purposiveness independent of the human mind.
It would be interesting to analyse these three possible attitudes and confront them with the effective positions that can be found in the literature on teleology in biology, but this is not the aim of the current work. The question here is rather: what is the minimal notion of teleology (Pérez-Escobar Reference Pérez-Escobar2024) that we need to defend in order to secure the notion of organic value?
Recall the two kinds of statements implying organic value:
(i) An organic trait (x) is instrumentally good for an organic entity (O) to achieve the particular organic purpose P.
(ii) The particular organic purpose (P) is instrumentally good for an organic entity (O) to achieve the general organic purpose of survival and reproduction (P*)
To secure the notion of organic value, it is necessary to make sense of ‘organic purpose’ with reference to organismal activities, for example, flying or surviving. Thus, I am interested here in defending the idea that it is meaningful to speak of purposes concerning the outcomes of the evolutionary process, that is, living beings.
This being said, making sense of purposes can be done in several ways. It is possible to criticise teleology eliminativism, for example, by claiming that appealing to mechanisms is as anthropomorphic as appealing to purposes (Canguilhem Reference Canguilhem1952). It is also possible to adopt a strong defence and argue that, ontologically, organisms are goal-directed. This characteristic could be claimed to have internal sources, for example by drawing on an organizational account of teleology (Mossio and Bich Reference Mossio and Bich2017, Bich Reference Bich2024). Or it could be argued that teleology has external sources, as put forward by McShea (Reference McShea2023) in his novel account based on field theory.
Although I think there are good arguments for both criticising eliminativism and for defending the strong option of teleology, I will here stick to the weak option and argue for it by drawing on Lennox and Kampourakis (Reference Lennox, Kampourakis and Kampourakis2013).
4.5 Second Step in Answering the Objection: Functional Explanations for Organisms Imply an Axiological Polarity
A mountain’s slope can have a causal impact on rainfall. A bird’s wing, and an aeroplane’s wing, can have a causal action on flight (Lennox and Kampourakis Reference Lennox, Kampourakis and Kampourakis2013). What is the difference between these cases?
All of the cases can be understood as identifying a cause and an effect. However, the case of the bird and the aeroplane share a commonality that is not present in the case of the mountain. In both the organic and technical cases, the effect of ‘flying’ plays a particular role. Flying contributes to explaining why, over history, birds and aeroplanes came to present the structural feature of having wings. By contrast, in the case of inorganic entities, we do not appeal to the rainfall to explain the mountain’s slope.
However, as Lennox and Kampourakis (Reference Lennox, Kampourakis and Kampourakis2013) emphasize, we should also note an important difference between organisms and artefacts. In the case of artefacts, it is a conscious agent who identifies the most appropriate wing design, while in the biological case it is the unconscious process of natural selection. In biology, design-based teleological explanations are inappropriate because they appeal to the intention of a supernatural creator. On the other hand, natural selection-based teleological explanations are appropriate, referring to an unintentional process (the mechanistic ‘filtering’ by natural selection).
Despite this relevant difference, we are in the same situation for organisms and artefacts: we need teleological explanations in the sense of Larry Wright’s conception of teleological explanation, that is, consequence aetiology (Wright Reference Wright1976). According to this conception, a teleological explanation is one in which the presence of x (a property, process or entity) can be explained by appealing to a result or consequence that x brings about. Such teleological explanations can be presented by referring to the concept of function. Bird and aeroplane wings are said to have the function of flight, but a mountain’s slope does not have the function of producing rainfall, just the effect of producing rainfall.
In the past half century, the literature on teleology in biology has focussed on the concept of function (McShea Reference McShea2023). The debate has been structured around two contributions from the 1970s by the philosophers Larry Wright and Robert Cummins (Gayon and Petit Reference Gayon and Petit2018, p. 155). Wright (Reference Wright1973) provides the origin of etiological approaches of function, proposing a focus on the history of traits – whether they are organic traits or technological artefacts. Starting from Wright’s proposition, Karen Neander (Neander Reference Neander1991) proposes the following definition: a trait’s function is the effect for which this trait has been selected over evolutionary history. Opposing the idea that historical considerations are pertinent in defining function, Cummins (Reference Cummins1975) proposes the thesis that something has a function if it has a causal role within a system. Alongside the etiological and systemic accounts of function, a third approach can be distinguished (Wouters Reference Wouters2003): the life chances approach (e.g., Bigelow and Pargetter Reference Bigelow and Pargetter1987). According to this approach, functions are considered as the effects that enhance the chances of survival in those that bear them.
Regardless of the specific account that one prefers, there is a general feature that is common in functional language: claiming that x has a function implies that x can dysfunction. This normative dimension gives the functional discourse an axiological polarity. Trait x is supposed to do something. If it can do that effectively, it is functional; if not, it is dysfunctional. George Gaylord Simpson expresses a similar view about the relationships between function and organic goodness in Simpson (Reference Simpson and Tax1960) and (Reference Simpson1974) (see also De Cesare Reference De Cesare2019).
Thus, let us return to our statements about organic value and transform them in the following way:
(iii) An organic trait (x) is instrumentally good for an organic entity (O) to achieve the particular organic function F.
(iv) The particular organic purpose (F) is instrumentally good for an organic entity (O) to achieve the general organic purpose of survival and reproduction (F*).
In the organic context, instrumental good or bad makes sense with respect to biological functions, whether particular or general. I have argued that, given the specificities of organisms (and artefacts), it is legitimate to use functional explanations, which are characterized by the fact that the effect contributes to explaining the cause. As the concept of function comes with an axiological polarity, this suggests that appealing to the notion of organic value is epistemically justified, rather than related to some suspicious anthropocentric or cultural bias.
4.6 Summing Up and Going Forward
At the end of Section 3, I was left with the following question: Dawkins’ notion of local progress refers to a ‘betterment’, which in turn presupposes the notion of ‘goodness’, but are these normative terms acceptable within a scientific theory such as evolutionary theory?
In this section, I have claimed that it is possible to defend a specific notion of instrumental organic value, drawing on the framework of Rosenberg and McShea (Reference Rosenberg and McShea2008). I have argued that organic value should be clearly distinguished from other kinds of value, such as moral or aesthetic. In this notion, there is no need to attribute value to survival and reproduction, which nonetheless represent the endpoint of value attributions. Moreover, drawing on Wittgenstein, I have highlighted that organic value can (at least in principle) be objectively assessed, provided that the end is clearly identified.
I have then presented an objection to this view, which I have called the anti-teleological objection to organic value. To respond to this objection, I have proposed the following reasoning:
We are epistemically justified in appealing to functional explanations for living beings (given the difference from explanations of inorganic entities).
Functional explanations imply an axiological polarity.
We are epistemically justified in appealing to an axiological polarity, which can be described in terms of organic value.
If this reasoning is correct, it affirmatively answers the question about the acceptability of normative terms: it is possible to accept the notion of organic value – and thus speak of goodness – within evolutionary biology. This boils down to answering question 1 (Are there any theoretical arguments for using the normative terms ‘good’ and ‘better’ within evolutionary theory?) with an affirmative.
Let us now return to the second premise put forward by Dawkins: the process of adaptation by natural selection conceptually implies local progress. Now we know that betterment can be spelled out in terms of organic value, and we are able to clarify what exactly this second premise means. The first step is to examine a few concrete examples where the process of adaptation by natural selection comes into play.
5 Empirical Cases from the Fossil Record
5.1 Looking for Local Progress in the Fossil Record
In presenting his notion of local progress, Dawkins claims that it holds from a purely theoretical standpoint, that we should accept it ‘without stirring from the armchair’ (Dawkins Reference Dawkins1997, p. 1018). However, I think that the adequacy of a theoretical notion should also be evaluated based on its application to empirical examples. Interesting cases in this context include those for which we have the strong intuition that something ‘gets better’ over evolutionary history. To present such empirical cases, we need to look at the fossil record. Although ‘often frustrating in its incompleteness and inadequacy’ (Vermeij Reference Vermeij1987, p. 359), the fossil documentation represents the main source of evidence for reasoning about organic progress. Here I will discuss two concrete cases relating to mollusc shells and seal swimming appendages. This will provide the basis for further examining the notion of local progress in Section 6.
5.2 Vermeij’s Molluscs
I have already pointed out that even Gould admits that a few cases of organic progress have been documented (Gould Reference Gould1989, p. 16); in doing so, the palaeontologist refers to his colleague Vermeij, the author of the book Evolution and Escalation. An Ecological History of Life (Vermeij Reference Vermeij1987). Along with Dawkins, Vermeij is another important champion of organic progress, sharing the Darwinian idea of ‘competitive highness’. I cannot investigate the details of Vermeij’s position on organic progress here. However, I will say a few words about his idea of escalation and his methodology before presenting the specific case of marine molluscs.
For Vermeij, competition for resources and predation are the most common and important selective factors acting on organisms, while physical calamities play a secondary role (Vermeij Reference Vermeij1987, pp. 19 and 24). Vermeij considers the Phanerozoic, the eon covering the last 542 million years, and claims that during this eon there has been no radical change in abiotic factors, excluding a few short-term catastrophic events. Thus, organisms have evolved during the Phanerozoic against the common background of physically similar habitats.Footnote 9
With this presupposition, Vermeij formulates a first hypothesis: adaptive traits favouring competitive and anti-predatory capacities should have seen an increase during the Phanerozoic. Moreover, the fact that an organism is better adapted in a given ecological context would also affect other organisms, resulting in an increase in adaptations in the latter. Vermeij refers to the Red Queen Hypothesis by Leigh Van Valen, according to which species must be kept ‘running in place’ by incessant adaptations just to stay alive (Vermeij Reference Vermeij1987, p. 4). According to Vermeij, this mechanism increases the frequency and severity of environmental hazards. As a consequence, organisms able to cope with risks should express more and more adaptations. This is the hypothesis of escalation (Vermeij Reference Vermeij1987, p. 49).
Interestingly, Vermeij notes that adaptation is not the only possible consequence of the increase of environmental hazards. Two alternative consequences are more common than adaptation: (1) ‘restriction or displacement of individuals to sites where conditions are more compatible with existing traits’ and (2) ‘death or extinction’ (Vermeij Reference Vermeij1987, p. 15).
But how could we possibly test the hypothesis of escalation? Vermeij suggests appealing to the uniformity principle: we should first examine the correspondence between the morphology, the habits and the environment in living organisms. Then, we can apply this information to the fossil record (Vermeij Reference Vermeij1987, pp. 54 and 61). Nevertheless, to apply this methodology, we have to narrow down the investigation to adaptations that leave traces in the fossil record and ‘whose history can be reconstructed with some confidence’ (Vermeij Reference Vermeij1987, p. 61).
According to Vermeij, the most suitable adaptations are related to resistance to predation in prey, especially when selective pressures affecting them leave traces in the preservable hard parts. This is why mollusc shells, devoted to the biological function of defence from predators (e.g., crustaceans), represent an excellent model. The idea is that the study of wounds and perforations in the shell makes it possible to infer the performance of these organic traits. Following the hypothesis of evolutionary escalation, the level of environmental hazard should increase, implying that molluscs are increasingly exposed to predation over evolutionary time.
Vermeij claims that the fossil documentation supports such a trend because we observe an improvement in the shell from a biomechanical standpoint:
Traits that prevent access to the aperture, strengthen the shell, and enable the foot to withdraw deeply into the shell increased in incidence and in degree of expression over time in assemblages of warm-water gastropods from unconsolidated bottoms, whereas traits associated with mechanical weakness, unobstructed openings, and limited retractability declined.
Let us simplify Vermeij’s results and consider only one of the traits mentioned here: the shell’s thickness. Figure 3 schematically depicts how the shell’s thickness differs in an organism of the ancestor population and in an organism of the descendant population.
Figure 3 Schematic representation a mollusc trait studied by Vermeij: the shell’s thickness. On average, descendants present thicker shells.
Vermeij’s example of marine molluscs is particularly suited to illustrating a case where we have the intuition that some improvement has taken place. With a thicker shell, descendants will have higher probabilities than ancestors to resist shell-crushing predators. Before proceeding further, let us consider another empirical case of a similar kind.
5.3 Seal Swimming Appendages
Swimming appendages in seals are a canonical example of adaptation: ‘the seal’s flipper has been perfected by natural selection as a means of swimming’ (Orzack Reference 81Orzack and Ruse2009, p. 6). Observing the goodness of fit between the seal flipper and the aquatic environment, evolutionary theory supposes that this organic trait must have been different in the seal’s terrestrial ancestors. However, until recently, ‘fossil evidence of the morphological steps leading from a terrestrial ancestor to the modern marine forms has been weak or contentious’ (Rybczynski, Dawson et al. Reference Rybczynski, Dawson and Tedford2009).
The fossil evidence was provided by an impressive palaeontological discovery described in Rybczynski, Dawson et al. (Reference Rybczynski, Dawson and Tedford2009). In the summer of 2007, in Nunavut (Canada), the research team stumbled upon a nearly complete skeleton of a semi-aquatic carnivore from the Miocene epoch. The new taxon – which was called Pujila darwini – does not have flippers, presents a long tail and its overall form is otter-like. Its morphology seems to be adapted to freshwater conditions. Pujila darwini cannot be considered as the pinniped’s common ancestor; it is rather one of the extinct cousins of modern pinnipeds. However, we have good reason to think that Pujila darwini presents some morphological resemblances to the pinniped common ancestor, namely in the fact that appendages are not flippers but are rather similar to the limbs of modern terrestrial carnivores. Figure 4 shows the reconstructed silhouette of Pujila darwini and the silhouette of a modern seal.

Figure 4 Left: reconstructed silhouette of Pujila darwini, a cousin of modern pinnipeds whose limbs are similar to modern terrestrial carnivores. Right: silhouette of a modern seal with flippers as swimming appendages (drawings by Clémence Moulard, reproduced with the consent of the artist).
The scenario would thus be as follows. When the first populations of the pinniped lineage started to colonize oceanic waters, they faced a new problem: they needed to swim longer distances than in freshwater environments. With respect to this problem, trait variations towards the ‘flipper morphology’ happened to advantage some individuals. By virtue of their better performance in swimming, these individuals did ‘relatively better’ in terms of survival and reproduction. This adaptive process went on over a timespan, while the challenge of swimming in oceanic waters stayed the same. After a given amount of time, we reach the morphology that we observe today: the flipper is widespread in seal populations.
In both the mollusc and seal cases, there are all the elements highlighted by Dawkins: a lineage of ancestors-descendants, a precise timespan in which the environment is supposed to stay homogeneous, and a betterment of the ‘equipment for survival’ from a mechanical standpoint. Let us now see how the idea that local progress occurs in these lineages can be further clarified.
6 Refining Local Progress: Functional Improvement of Organic Traits
6.1 Definition of Functional Improvement of Organic Traits
We now have all the elements to propose a clear definition of local progress: Dawkins’ reasoning on local progress (Section 3), the clarification of the notion of organic value (Section 4), and two empirical cases from the fossil record (Section 5). To move forward, let us recall the statements (iii) and (iv) at the end of Section 4:
(iii) An organic trait (x) is instrumentally good for an organic entity (O) to achieve the particular organic function F.
(iv) The particular organic purpose (F) is instrumentally good for an organic entity (O) to achieve the general organic purpose of survival and reproduction (F*).
These two statements refer to the kind of goodness described by the notion of organic value. Yet, we have not made explicit how this can lead to statements concerning betterment over time. In this case, such statements would concern different traits, for example x and y. By reasoning in gradable terms, it is possible to compare traits and assess, for example, that trait ‘y’ is better than ‘x’ for a function. The betterment over time can then be characterized as follows:
Functional Improvement of Organic Traits (FIOT)
y is instrumentally better than x at performing F.
At the beginning of the considered timespan (‘ancestors’), we observe more instances of x.
At end of the considered timespan (‘descendants’), we observe more instances of y.
Given these observations, we conclude that over the considered timespan there has been a functional improvement with respect to F.
In the mollusc case, F is the function of defence from shell-crushing predators, while, in the seal case, F is the function of locomotion in oceanic waters. In both cases, organisms bearing relatively better traits with respect to F have more potential to survive and leave descendants who can inherit good traits (F*).
6.2 Three Missing Clarifications
At first sight, the FIOT notion that I have presented is easily applicable to these two empirical cases from the fossil record considered in the previous section. Yet, with more careful examination, we can see that the analysis of local progress provided so far does not explicitly answer three questions:
In what way does the environment act as a causal factor modifying the population over time?
What precisely is getting better?
How often does this betterment occur?
This section focusses on clarifying these issues, which is needed to achieve a coherent and clear notion of local progress as FIOT.
6.3 First Clarification: The Improvement is Related to Specific Environmental Problems
I have pointed out that, in Dawkins and Vermeij’s approach, the homogeneity of the environment over the timespan is the assumption needed to avoid the problem of ‘incommensurability’ raised by Gould. In fact, if t1 and t1000 are characterized by completely heterogeneous environments (as is the case in Figure 1), it is impossible to fairly compare the organic forms at the respective times. By contrast, the homogeneity of the environment would guarantee the common background against which it is possible to compare functional performances of the ancestor and descendant population, which is a precondition for claiming that local progress occurred.
However, is it plausible to assume that over the timespan the whole environment stays homogenous? If we think about the intricate complexities of the environment of a population (e.g., parasites, predators, competitors, and resources), this assumption sounds very unrealistic.
Yet, based on examining the two empirical cases, I think that a more modest assumption would be sufficient to avoid the incommensurability problem. In the mollusc case, what stays constant over time is not the whole environment but rather (borrowing terminology from Richard Lewontin) a specific ‘environmental problem’ encountered by the population (Lewontin Reference Lewontin1978). For molluscs, the problem is related to the biotic component of the environment: the presence of shell-crushing predators. For seals, the environmental problem is the physical conditions of the environment, that is, oceanic waters. In both cases, the specific environmental problem influences the intra-population competition, thus being part of the selective environment (Brandon Reference Brandon1990).
From this perspective, the suggestion would be as follows: instead of making an unrealistic assumption about the homogeneity of the whole environment at t1 and t1000, it is possible to narrow down the assumption to a specific environmental problem. Figure 2 should thus be modified to obtain the representation in Figure 5. In this, ‘natural selection’ is the name of an environmental problem remaining stable over a timespan and being responsible for differential fitness among individuals of the population.Footnote 11
Figure 5 Representation of two populations (t1 and t1000) with a specific environmental problem as a cause of betterment. This avoids the incommensurability problem without assuming the homogeneity of the whole environment at the two times.
6.4 Second Clarification: The Focus is on the Improvement of Isolated Traits Ceteris Paribus
Another point still needs to be clarified in Dawkins’ account of local progress. What does it mean that the organism’s ‘equipment for survival’ gets better when local progress takes place from t1 to t1000? To see this, we must recall that, in the empirical cases introduced earlier, the focus is on isolated and specific organic traits. This would suggest that the ontological substrate of the betterment consists of parts rather than whole organisms. However, is it possible to ‘atomize’ organisms and consider parts in isolation?
To settle this question, I will proceed in two steps. First, I will present an original conceptual model concerning the way in which biologists conceive organisms in relation to the environment. I will call this the ‘hesperidium model’ or, more simply, the ‘grapefruit model’. Second, I will present a relevant insight by Richard Lewontin that, combined with the conceptual model, will be useful to answer the question.
The Grapefruit Model
‘Hesperidium’ is the botanical name for the kind of berry typical of the genus Citrus, containing, for example oranges, lemons and grapefruits. A hesperidium is made of three components. Starting from the inside, there is the edible flesh of the fruit (called endocarp), which is divided into segments. Covering this is a white layer called mesocarp or albedo, that is, the white part of the grapefruit’s peel. Finally, there is the outer pigmented layer called exocarp or flavedo, that is, the yellow-orange outer part of the grapefruit’s peel. Now imagine the organism as a grapefruit cut across the middle, as presented in Figure 6.
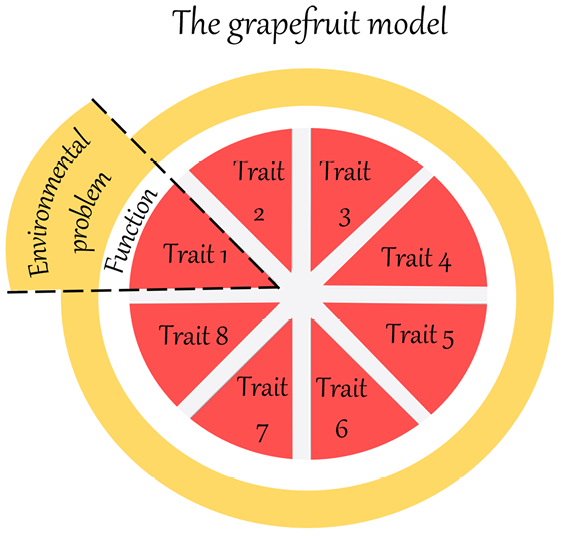
Figure 6 The grapefruit (or hesperidium) model illustrates the relationships between organic traits, biological functions and environmental problems. The inner part corresponds to the structure of the organism, divided into segments representing organic traits. The white layer represents the functionalities of the organism, and the outer layer corresponds to the ensemble of environmental problems encountered by the organism.
Starting from the inside, there is the endocarp (the grapefruit’s flesh). This would correspond to the morphological structure of the organism, which can be decomposed into organic traits. Biologists are aware of the complex interactions of organic traits within the organism, but it is common practice to ‘segment’ the parts of the organism in traits. While doing this, the hope is that we ‘carve nature at its joints’ (to employ Plato’s metaphor), so that our epistemological carving is not too artificial.Footnote 12 Thus, we can imagine that biologists separating the wedges of a grapefruit are distinguishing different organic traits of a whole organism (e.g., for a mollusc: the shape of the aperture, thickness of the shell, etc.).
If Figure 6 represents an individual organism, note that at the population level organic traits are characterized by countless small morphological variations, which are the basis upon which natural selection takes place. For example, measuring the thickness of a shell within a mollusc population, we will obtain a huge number of different values (from which, of course, we can obtain the average shell thickness of the population).
Then we come to the grapefruit’s peel. The second layer corresponds to the global functioning of the organism and the outer layer to the ensemble of environmental problems encountered by the organism. In the real grapefruit, the mesocarp and exocarp form the peel, which does not show any segmentation. However, if I want to eat a grapefruit, it is possible to section the peel by cutting the mesocarp and the exocarp at the same time. In an analogous way, biologists can conceptually carve out both a biological function (within the global functioning of the organism) and a corresponding environmental problem (within the ensemble of the environmental problems encountered by the organism) in the same move. In the same cut, there is a correspondence between a function and a problem.
It is relevant to note that, between the grapefruit’s wedges and the cutting of the peel, there is not necessarily a biunivocal correspondence. Outside the metaphor, it is possible that several organic traits contribute to one function, and it is also possible that the same organic trait takes place in several organic functions.
Lewontin’s Insight about the Ceteris Paribus Clause
How is this botanical detour helpful? I was left with an open question: is it possible to atomize organic traits and consider them in isolation? I think that it is now possible to answer this by combining the grapefruit model with some relevant remarks by Lewontin (Reference Lewontin1978). In his paper ‘Adaptation’, Lewontin identifies a feature that is required in explaining organic adaptations: the ceteris paribus assumption of ‘all other things being equal’:
In order to make an argument that a trait is an optimal solution to a particular problem, it must be possible to view the trait and the problem in isolation, all other things being equal.
The ceteris paribus clause is widespread in the field of economics. Its use can be traced back to the thirteenth century and it was popularized in the nineteenth century by Alfred Marshall (Reutlinger, Schurz et al. Reference Reutlinger, Schurz, Hüttemann, Jaag, Zalta and Nodelman2024), author of the influential book Principles of Economics (Marshall Reference Marshall1890). Marshall explains that, in answering complex questions of economics, causes have to be considered in isolation:
The forces to be dealt with are, however so numerous, that it is best to take a few at a time; and to work out a number of partial solutions as auxiliaries to our main study. Thus we begin by isolating the primary relations of supply, demand and price in regard to a particular commodity. We reduce to inaction all other forces by the phrase “other things being equal”: We do not suppose that they are inert, but for the time we ignore their activity.
Interestingly, Lewontin’s claim seems to be that the biologist who wants to explain adaptations has to use the same methodology. He argues:
If all other things are not equal, if a change in a trait as a solution to one problem changes the organism’s relation to other problems of the environment, it becomes impossible to carry out the analysis part by part, and we are left in the hopeless position of seeing the whole organism as being adapted to the whole environment.
Lewontin remarks that it is a ‘hopeless position’ to consider the whole organism as adapted to the whole environment. This is the same as the idea of the incommensurability criticism that has already been encountered with Gould (Figure 1). Adaptation requires us to ‘carry out the analysis part by part’ and to be able to isolate a constant environmental problem as a common background for organic comparisons. It is interesting to note that, although joining Gould in the criticism of the excesses of the adaptationist programme (Gould and Lewontin Reference Gould and Lewontin1979), Lewontin emphasizes that we cannot abandon the notion of adaptation:
On the other hand, to abandon the notion of adaptation entirely, to simply observe historical change and describe its mechanisms wholly in terms of the different reproductive success of different types, with no functional explanation, would be to throw out the baby with the bathwater. Adaptation is a real phenomenon. It is no accident that fish have fins, that seals and whales have flippers and flukes, that penguins have paddles and that even sea snakes have become laterally flattened. The problem of locomotion in an aquatic environment is a real problem that has been solved by many totally unrelated evolutionary lines in much the same way. Therefore it must be feasible to make adaptive arguments about swimming appendages. And this in turn means that in nature the ceteris paribus assumption must be workable.
This presents a line of defence against the accusation of excessive atomization of living beings: analysing organisms ‘part by part’ (and supposing the ceteris paribus clause) is a theoretical prerequisite for the notion of adaptation. We would be throwing out ‘the baby with the bathwater’ in dismissing this notion. Without it, we could not explain the existence of fins, flippers and countless other organic features that we observe. Thus, we have to accept the theoretical need to isolate both traits and problems.
Lewontin claims that this epistemological choice has an ontological counterpart in nature: ‘in nature the ceteris paribus assumption must be workable’ (Lewontin Reference Lewontin1978, p. 230). In the last passage of the paper, Lewontin clarifies that the assumption can only be workable ‘if both the selection between character states and reproductive fitness have two characteristics: continuity and quasi-independence’ (Lewontin Reference Lewontin1978, p. 230). Continuity means that a small change in one trait results in a small (and not dramatic) change in another. For example, if a small change in fin shape had the effect of making the organism suddenly attractive to a new predator, the adaptive process concerning the fin’s shape could not occur. By quasi-independence, Lewontin seems to mean that it must be possible that selection acts on an isolated characteristic ‘without altering other characteristics of the organism in a countervailing fashion: pleiotropic and allometric relations must be changeable’ (Lewontin Reference Lewontin1978, p. 230). For Lewontin, ‘continuity and quasi-independence are the most fundamental characteristics of the evolutionary process’ (Lewontin Reference Lewontin1978, p. 230) because, without them, adaptive evolution would have been impossible.
Let us try to make explicit Lewontin’s argument with the mollusc example. Suppose that our aim is to explain an adaptive trait of extant molluscs: the thickness of the shell. The explanandum is the process of adaptation by natural selection occurring over an evolutionary timespan. However, this explanandum requires two things: considering the trait in isolation and applying the ceteris paribus clause. In the timespan, organisms with the organic trait ‘thicker shell’ have an advantage over organisms with a ‘thinner shell’ relative to the problem of defence against predators. However, besides this, it has to be supposed that there is no interference from other environmental problems regarding the evolution of the trait. Things can of course change, but not in such a way as to interfere with the adaptive process concerning the thickness of the shell. To visualise the need for this assumption, imagine the counter-scenario: at some point in the timespan under consideration, new parasites arrive in the population of gastropods, representing a novel environmental problem. Then, imagine that (for some reason) the new parasites preferentially attack gastropods with thicker shells rather than gastropods with thinner shells. The result in terms of differential reproduction would now be in favour of thinner-shelled gastropods. But this is not what we actually observe. Thus, we can infer that this counter-scenario did not take place. Instead, we can legitimately argue that over this timespan the adaptive process concerning the thickness of the shell occurred while the ceteris paribus condition was holding with respect to other environmental problems.
Combining the Grapefruit Model with Lewontin’s Insight
By combining Lewontin’s insight with the grapefruit model, I can finally present a refined picture in Figure 7.
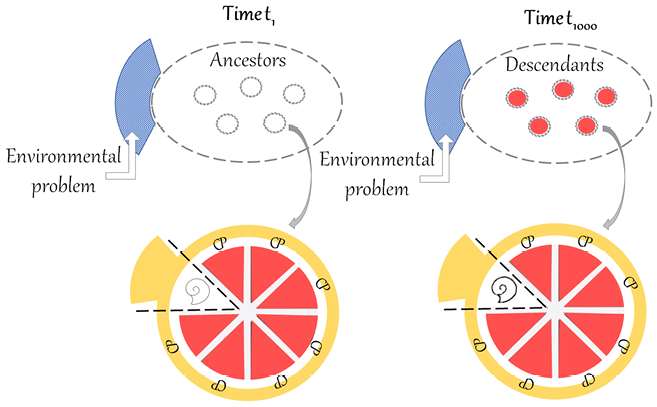
Figure 7 A refined picture of local progress, accounting for the grapefruit model and Lewontin’s insight about the ‘part by part’ analysis and the ceteris paribus assumption.
In this reconstruction of the adaptive process, we must assume that several conditions are given over the timespan:
A specific environmental problem that stays (relatively) constant over the timespan.
Correspondence of the environmental problem with functional aspects (related to structural traits).
Intra-population variations, resulting in more or less good functional performances with respect to the environmental problem.
The ceteris paribus assumption.
When all these conditions are satisfied, the process of adaptation by natural selection can occur, that is, variations of traits presenting better functional performances (with respect to the environmental problem) are accumulated over the relevant timespan, while we suppose that the ceteris paribus condition is holding. Thus, it must be possible to say that what improves at a populational level are organic traits, while all other things are equal. It is thus possible to modify the structure of Dawkins’ reasoning in this way.
P1: Evolutionary theory conceptually implies the possibility that the process of adaptation by natural selection takes place.
P2: The process of adaptation by natural selection can be characterized as FIOT.
C: Evolutionary theory conceptually implies the possibility of FIOT.
Thus, in my view, whenever an adaptive process occurs, FIOT occurs. It is important to note, however, that this in no way means that adaptive processes/FIOT occur all the time. This will be the object of the third clarification.
6.5 Third Clarification: The Frequency of Functional Improvement of Organic Traits
The process of adaptation by natural selection is only one of the processes, though an important one, that characterize organic evolution. As Vermeij interestingly notes, when a population experiences an increase of environmental hazards, adaptation is far from being the only thing that can occur. Two alternative consequences are more common than adaptation: (1) ‘restriction or displacement of individuals to sites where conditions are more compatible with existing traits’ and (2) ‘death or extinction’ (Vermeij Reference Vermeij1987, p. 15). This is coherent with what has been said so far: evolutionary theory conceptually implies the possibility (and in no way the necessity) of adaptive processes/FIOT. However, this is compatible with three different claims concerning the frequency of this phenomenon over evolutionary history:
It is possible that the adaptative process/FIOT occurs and this happens sometimes.
It is possible that the adaptative process/FIOT occurs and this happens most of the time.
It is possible that the adaptative process/FIOT occurs and this happens rarely.
In refining the FIOT notion, I had the first claim in mind. Within evolutionary theory there is a need for an explanandum for at least some cases of adaptations. This explanandum is the process of adaptation by natural selection, which – if one accepts the reasons grounding organic value – implies FIOT. All that is required for this reasoning is that at least some cases of adaptative process/FIOT exist; nothing more is said about the frequency of this phenomenon.
However, what should we think about the two other claims? The endorsement of the second corresponds with what is known as ‘adaptationism’ and implies that optimal organic traits are widespread in the biological realm.Footnote 13 On the other hand, according to the third claim, optimal traits rarely occur in evolutionary history.
Are there any good reasons to favour one of the two claims? I think that Orzack (Reference 81Orzack and Ruse2009) grasps a relevant point when he suggests that the position taken by biologists about adaptationism amounts to an a priori judgement. Some biologists, such as John Maynard Smith, think that adaptationism is a priori true: optimal traits are everywhere in nature (Orzack Reference 81Orzack and Ruse2009, p. 19). Others, in Gould’s vein, think that adaptationism is a priori false, that is, optimal traits are rare in nature.
But should we accept a priori judgements about adaptationism? A suggestion by Orzack (Reference 81Orzack and Ruse2009) is particularly interesting in this regard: rather than sticking to a priori judgements, the question should be investigated in empirical terms by studying the local optimality of traits (Orzack and Sober Reference Orzack and Sober1996). Several outcomes would be possible based on this kind of investigation. It could turn out that, among the cases considered, 90% count as locally optimal traits, and this would support the second claim about the frequency of the adaptative process/FIOT. However, it could also turn out that the percentage is only 20%, which would be rather in favour of the third claim. This kind of empirical study would be ideal for settling the issue. However, it is not straightforward to perform such a study at a large scale and with a homogeneous protocol. As a result, it is hard to draw final conclusions on this issue.
Thus, the safer epistemic option would seem to be to stick to the first claim: an adaptative process/FIOT occurs sometimes. However, as Gould and Lewontin (Reference Gould and Lewontin1979) point out, the adaptationist claim had some success over the history of evolutionary biology in the twentieth century. Why might evolutionary biologists be led to easily jump from ‘FIOT happens sometimes’ to ‘FIOT happens most of the time’?
On this point, I agree with Gould’s intuition concerning the fact that some extra-scientific bias is coming into play in the biological thought. The biases can be of several kinds. For example, they might be due to the influence of the analogy between organisms and technological artefacts. As FIOT is more widespread in artefacts and as the structure of the reasoning is similar, biologists overestimate the frequency of FIOT in the organic realm (De Cesare Reference De Cesare2019). Also, the cult of performance and optimality, dominant in occidental cultures, might lead biologists to think that the adaptive process/FIOT is everywhere (Hamant Reference Hamant2022).
Further exploration of this issue requires another work. However, I hope I have sufficiently clarified my position. Evolutionary biologists who cannot ‘live without’ organic progress are driven by the intuition that the adaptive process conceptually implies some betterment and, as we need to appeal to the adaptive process, we also have to retain the idea of betterment. I have tried to show that this intuition can be clarified: the notion of organic goodness is related to functional explanations, and the need for functional explanations within biology is epistemically grounded. Given this functionally grounded notion of organic value, it is possible to characterize the adaptive process as FIOT. However, some bias might come into play when biologists ‘make a leap’ that is not supported by empirical evidence, transforming the claim ‘FIOT happens sometimes’ to ‘FIOT happens most of the time’.
6.6 Note on Fitness
Before proceeding further, I will add a note here concerning the relationship between FIOT and the concept of fitness. To begin, it is worth noting how this concept is generally characterized in the literature of philosophy of biology.
The notion of fitness can be understood in two ways: ecological and evolutionary fitness. Ecological fitness (Rosenberg and Bouchard Reference Rosenberg, Bouchard, Zalta and Nodelman2023) refers to the correspondence between organismal traits and various aspects of the environment. This notion is related to the idea of appropriate organism ‘design’. For example, what makes a wing fit for flying, or a heart fit for pumping blood (Gayon and Petit Reference Gayon and Petit2018, p. 293)? Rosenberg and McShea seem to refer to a similar concept when speaking of fitness as solving environmental design problems (Reference Rosenberg and McShea2008, p. 55). Interestingly, they also use the terminology of environmental problems – as I borrowed from Lewontin.Footnote 14
If ecological fitness puts more emphasis on survival of the individual organism (its dealings with the environment), evolutionary fitness rather refers to the contribution that the organism makes to future generations. Thus, with evolutionary fitness, the interest shifts from the individual organism to the level of populations. This definition has been converted in measurement with population genetics and particularly with the work of Ronald Fisher. In fact, Fisher transforms the qualitative concept into a quantitative measure, including mortality and fecundity rates (Gayon and Petit Reference Gayon and Petit2018, p. 248).Footnote 15 Some authors (for example, Brandon (Reference Brandon1990)) have proposed to modify the evolutionary concept of fitness by adopting a probabilistic propensity definition. In this account, a fit organism is one that has a probabilistic propensity to leave descendants (it might happen that, ‘unluckily’, the fit organism does not reproduce).
The debate over fitness is still open in the philosophy of biology. When the term is used without specification, we should ask whether it is the ecological or the evolutionary concept that is being invoked. In my analysis, the relevant question to ask is which kind of fitness comes into play in the notion of FIOT. Can we affirm that descendants at t1000 have a better fitness than ancestors at t1? If by fitness we mean ecological fitness, the answer is yes. If FIOT has occurred, this means that a descendant at t1000 has some traits that better solve (on average) a specific environmental problem than an ancestor at t1, considering that the two are equal with respect to other problems (ceteris paribus). However, the answer is not the same if by fitness we mean evolutionary fitness. As noted by Dawkins, evolutionary absolute fitness might be the same: at t1000, an organism might leave the same absolute number of descendants (on average) as the ancestor at t1.
Thus, once FIOT has occurred, descendants at t1000 are not necessarily better than their ancestors at t1 in terms of absolute evolutionary fitness. However, let us consider relative evolutionary fitness. In the adaptive process taking place over the timespan, some organisms within the population happened to be better in terms of ecological fitness. For example, let us suppose that the first seals with a flipper morphology were better ‘problem solvers’ in oceanic environments compared to seals of the same populations lacking this trait. By virtue of the differences with respect to ecological fitness, some organisms (seals with a flipper-morphology) had a better relative evolutionary fitness with respect to other organisms in the same population (seals lacking this trait).
6.7 Is There a Way to Observe Functional Improvement of Organic Traits?
At the end of this section, I think we have a satisfactory refined notion of FIOT, which accounts for what happened in empirical cases such as those discussed in Section 5. As I have pointed out, in these palaeontological cases we infer that an adaptive process/FIOT occurred in a lineage. This inference relies on some (reasonable) hypotheses, that is, the principle of uniformity can be applied to compare descendants and ancestors and that an environmental problem is relatively constant while the ceteris paribus clause is holding.
However, it might be asked: is it possible to provide more direct evidence about the occurrence of FIOT in a lineage? For this, we would need some kind of time machine (as imagined by Darwin’s thought experiment) that allowed us to place ancestors and descendants in competition with one another. But how could this ever be possible?
It turns out that bacteria have some amazing properties and these have been used to transform Darwin’s thought experiment into a real experiment at small timescales. The next section will therefore be devoted to presenting the Long-Term Evolutionary Experiment (LTEE) by Richard Lenski to confront our characterization of FIOT with these results regarding experimental evolution.
7 Observing Functional Improvements by Time-Travelling with E. coli
7.1 The Long-Term Evolutionary Experiment
Several model organisms allow biologists to materialize Darwin’s thought experiment on competitive highness: this research field is known as ‘resurrection ecology’ (Burge, Edlund et al. Reference Burge, Edlund and Frisch2018, Weider, Jeyasingh et al. Reference Weider, Jeyasingh and Frisch2018). With his research on experimental evolution, the biologist Richard Lenski is considered a pioneer in this field. The LTEE began in 1988 on twelve populations of Escherichia coli and it is still ongoing today. The primary motivation of the experiment was precisely to address questions about evolutionary dynamics, for example: is it possible to observe adaptation by natural selection, whereby the performance of bacteria would tend to improve over time (Lenski, Mongold et al. Reference Lenski, Mongold, Sniegowski, Travisano, Vasi, Gerrish and Schmidt1998)? Would all populations improve their performance in a similar way or not? How do evolutionary novelties emerge?Footnote 16
7.2 Experimental Protocol and Competition Experiments
Lenski’s study began with a single ancestor strain of E. coli, from which individual cells were taken to found twelve populations, allowed to evolve in separate flasks (Lenski, Rose et al. Reference Lenski, Rose, Simpson and Tadler1991, Lenski and Travisano Reference 80Lenski and Travisano1994). The populations grow in a simple environment that is kept as constant as possible. They have plenty of oxygen, an incubator at 37°C and no other species in the same flask (Lenski Reference Lenski2024).Footnote 17 The substrate is mainly composed of glucose, potassium phosphate, citrate and a few other elements. The carbon source intended to feed E. coli is glucose, present in the medium in limited quantities. The reproduction of bacteria is strictly clonal (asexual) and runs through six or seven generations a day. Every day following the start of the experiment, a sample is taken from each population (1% of the volume) and transferred into a fresh medium where the population continues to propagate. This process has been carried out every day since 1988Footnote 18 and Lenski’s team reached 75,000 generations in June 2022 (Lenski Reference Lenski2024).
Every 500 generations (around 75 days), a sample from each population is stored at -80°. The frozen cells remain perfectly viable, so they can be defrosted and revived at a later time. This is precisely the property that allows the kind of ‘time travel’ that seemed impossible to Darwin. In fact, this means that Lenski and his colleagues have a ‘frozen fossil record’ and are able to put the ancestral strain and the later-generation samples into competition with one another, with genetic markers making the distinction between the two possible. As Lenski and Travisano (Reference 80Lenski and Travisano1994) put it, this approach might be called ‘experimental palaeontology’.
To perform competition experiments, Lenski and his team defrost a sample of an ancient generation (ancestors, A) and put it in the same flask as a sample of the current generation (descendants, D). A and D can be distinguished with a colour marker. A sample of the solution is plated out to look at the abundances of the two populations at the start of the experiment. The flask is incubated for a day and then plated out again. This process is repeated for the duration of the competition experiment. The colonies (each starting from a single cell) can be counted by hand to quantify the respective abundances.
7.3 Structural Aspects in Ancestors and Descendants: Comparing Cell Size
What are the main differences between A and D? With respect to the biological examples discussed earlier (molluscs and seals), here we are in a situation where distinguishing traits of the organism is much less straightforward.
However, a striking morphological difference has been noted by Lenski’s team between A and D: the D cells are bigger than the A cells. In all twelve populations that were initially the same size, there is an increase in average cell volume already in the first 10,000 generations (Lenski and Travisano Reference 80Lenski and Travisano1994). This increase in average cell volume was surprising to the researchers, who would have expected that cells would become smaller, based on surface-to-volume considerations in an environment where resources are limited (Lenski Reference Lenski2024). A recent study performed by Grant, Magid et al. (Reference Grant, Magid, Franklin, Dufour and Lenski2021) confirmed the trend towards larger cells over 50,000 generations.Footnote 19
7.4 A First Functional Aspect about Ancestors and Descendants: Comparing Growth Rates
Besides the structural feature of a bigger morphology, are there any striking differences between A and D from a functional standpoint? The results of competition experiments can elucidate this question. A first functional aspect that is studied by Lenski and his team concerns bacterial growth rates, that is, the speed at which an E. coli cell can consume resources, grow and reproduce asexually.
Growth rates are estimated by looking at the number of colonies. At different times in the competition experiment, colonies of D and A are counted. These data are used to calculate the ratio of the number of D and A colonies. A ratio of 1 means that the two populations have similar cell counts, as was the case at the beginning of the competition experiment. A ratio of 1.5 means that the population of D is 50% more abundant on the plate. In Figure 8, the vertical axis represents this ratio, which is considered by Lenski et al. as a measure of ‘relative’ or ‘competitive’ fitness. The horizontal axis represents the time from the start of the experiment, measured in bacterial generations. Note that the environment is kept stable over this competition experiment.
Figure 8 Results of head-to-head competition experiment between D and A over 10,000 generations. Relative fitness is calculated as the ratio of realized growth rate of the D and A (Lenski and Travisano Reference 80Lenski and Travisano1994, p. 6810).
The results show that, already in the first 2,000 generations, the D population density has undergone a rapid increase. After 10,000 generations, D shows a 50% greater growth rate than A (the ratio increasing at around 1.5). Moreover, the researchers have sampled the bacteria at shorter intervals during the first 2,000 generations and observed that the trajectory of the increasing relative fitness of D is step-like (Lenski and Travisano Reference 80Lenski and Travisano1994).
The fact that D cells grow and reproduce faster than A cells provides, according to Lenski, ‘a simple and compelling demonstration of the process of adaptation by natural selection’ (Lenski Reference Lenski2024, Introduction, Section 3). Here, natural selection acts in an extremely simple and constant selective environment, where the limiting factor is the amount of resources available in the medium.
Another surprising result has been obtained in the LTEE concerning growth rates. One might think that, in a constant environment, the increase in ‘competitive fitness’ would stop at some point, so that the curve attains an asymptote. However, it turns out that the mean-fitness trajectories measured in the experiment better fit a power-law model, which has no upper bound (Wiser, Ribeck et al. Reference Wiser, Ribeck and Lenski2013). Also, the power-law model – applied to a truncated dataset only including the first 20,000 generations – predicts the future dynamics in a more accurate way than a model with an asymptote.
For Lenski, this suggests that adaptation by natural selection does not only occur where there are significant environmental changes, but can also keep going in constant environments. Lenski speaks of ‘endless adaptations’: ‘the bacteria continue to become better and better adapted to the LTEE environment over time, and it appears their fitness may continue to increase indefinitely, albeit at a slower pace.’ (Lenski Reference Lenski2024, Introduction, Section 3)
7.5 A Second Functional Aspect: Energy Efficiency
The LTEE suggests that D cells are more effective than A cells with respect to the functional aspect of consuming resources quickly, growing and reproducing. However, interestingly, another functional aspect has been studied by Lenski and his colleagues: energy efficiency in terms of ‘production cost’ of the cell. Being efficient, for an E. coli cell, would mean growing by using a small amount of resources. It is important here to distinguish effectiveness from efficiency. Something is effective if it accomplishes a purpose; something is efficient if it uses the minimum amount of resources (in accomplishing a purpose).
Note that there might be a discrepancy between these two functional aspects: a cell might be very effective in quickly consuming resources, growing and reproducing, but it could at the same time be inefficient, that is, show a bad performance in terms of production cost. Microbiologists are able to obtain evidence about energy efficiency by measuring the bacterial yield. This can be estimated by counting the number of viable cells per unit of a limiting resource (Vasi, Travisano et al. Reference Vasi, Travisano and Lenski1994). Thus, to compare the performance of A and D with respect to energy efficiency, a question can be asked: if I give a unit of resource to A and exactly the same unit of resource to D, how many viable cells do I obtain, respectively? With respect to this question, I will focus here on the results of two studies performed by Lenski’s collaborators: Vasi, Travisano et al. (Reference Vasi, Travisano and Lenski1994) and a recent study by Marshall, Malerba et al. (Reference Marshall, Malerba, Lines, Sezmis, Hasan, Lenski and McDonald2022).
7.6 The Two Functional Aspects in Ancestors and Descendants: Comparing Growth Rates and Energy Efficiency
Vasi, Travisano et al. (Reference Vasi, Travisano and Lenski1994) performed experiments by putting A and D in competition. However, there was a modification with respect to the experimental protocol described so far. Namely, the environment was not kept constant: the competition assays were done by simulating a ‘seasonal’ environment, alternating periods of feast and famine over bacterial generations. What did they observe? As in the case of the competition assays in a stable environment, D cells increased in maximum growth rate and cell size over 2,000 generations. Nevertheless, results regarding numerical yields did not show a tendency to increase. The authors estimated the yield in A and D separately, giving them the same amount of resources. They found out that, given the same unit of limiting resource, (small) A cells were more numerous than (big) D cells. Thus, D cells were more effective than A at quickly consuming resource, growing and reproducing, but they were less effective than A in growing by using a small amount of resources.
These results are extremely interesting in comparison with those of Marshall, Malerba et al. (Reference Marshall, Malerba, Lines, Sezmis, Hasan, Lenski and McDonald2022), published in a paper bearing the title ‘Long-Term Experimental Evolution Decouples Size and Production Costs in Escherichia Coli’. Here, the authors had 60,000 generations of E. coli and, unlike Vasi et al., they performed the experiment with a constant environment. They measured cell metabolism (quantifying it by measuring oxygen consumption) and bacterial yields. Overall, their results are as follows. As usual, D presented larger cells, roughly doubling in size. These larger cells, having a greater surface, consumed more oxygen than smaller A cells (in absolute terms, D has higher metabolic rates). However, relative to their size, metabolic rates were lower in D than in A. In fact, while D cell size increased by around 50% with respect to their ancestors, oxygen consumption increased only by 30%. And what about yields? Due to the relatively low metabolic rates of D, the maximum biovolume yields were higher in D than in A. The authors conclude:
Thus, the larger cells are metabolically more efficient and attain higher population biomass than smaller cells from a given amount of resource. This result conforms with other LTEE studies that found that the evolved cells are larger, are more efficient, and attain higher maximum biomass yields than the ancestors.
Thus, according to Marshall, Malerba et al. (Reference Marshall, Malerba, Lines, Sezmis, Hasan, Lenski and McDonald2022), D are better than A both in terms of quickly growing and reproducing and in terms of growing by using a small amount of resources. However, these results do not go in the same direction as the previous study of Vasi, Travisano et al. (Reference Vasi, Travisano and Lenski1994), where there is a discrepancy with respect to the two functional aspects. It should be kept in mind that, in the two studies, environmental conditions were not the same, diverging in resource availability (constant resources or seasonal variation of resources).
7.7 Results Concerning the Emergence of a Novel Function
Another result obtained in the LTEE is worth mentioning, even if it does not concern the direct competition between A and D. In one population of E. coli, something unexpected happened: one of the twelve populations discovered ‘new ways of making a living’ (Lenski Reference Lenski2024). As previously stated, the carbon source supposed to feed E. coli was glucose, present in the medium in limited quantities. However, since the beginning of the experiment, citrate had been added as a chelating agent in order to stabilize the medium. One day, Lenski’s team discovered that one population in a flask ‘looked strange’, with much more turbidity than others. After excluding the hypothesis of a contamination, the researchers realized that this population had evolved the capacity for aerobic citrate utilization (Cit+). Blount, Barrick et al. (Reference Blount, Barrick, Davidson and Lenski2012) have also analysed genome sequences to investigate the history and genetic basis of this capacity, in order to identify traits related to the emergence of this new function.
7.8 Conditions for Functional Improvement of Organic Traits Are Materialized in the Long-Term Evolutionary Experiment
For our purposes, the relevance of the LTEE consists in ‘materializing’ the conditions for FIOT in the experimental protocol. Recall the conditions for FIOT that I highlighted in Section 6:
A specific environmental problem for the population, staying (relatively) constant over the timespan.
Correspondence of the environmental problem with functional aspects (related to structural traits).
Intra-population variations, resulting in more or less good functional performances with respect to the environmental problem.
The ceteris paribus assumption.
Concerning the first condition, the experimental protocol of the LTEE allowed the researchers to create an extremely simple environment, if compared to the complexities of natural conditions. In the LTEE flasks, an E. coli cell only has one environmental problem: consume resources. It is possible to keep this problem constant by keeping the availability of resources stable (but, as previously explained, it is also possible to simulate seasonal conditions).
The second condition is also fulfilled in the LTEE: Lenski and his team studied aspects of E. coli that correspond to the problem of resource consumption. E. coli cells can be studied from a structural standpoint, that is, looking at phenotypic traits. These are less straightforward to study than in macroscopic organisms, although the morphological analysis can be complemented with genetic analysis. E. coli cells can also be studied from a functional standpoint, that is, by looking at different aspects of bacterial activity related to resource consumption, like growth rate and energy efficiency.
The third condition is a common characteristic of populations, even in artificial environments: organisms of a population, even with asexual reproduction, will not present exactly the same features. Thus, they might differ in functional performance related to the (relatively) constant environmental problem.
Finally, there is the condition of the ceteris paribus assumption. In LTEE, the environmental problem is fixed: consuming resources. Concerning all the other aspects of the environment, the LTEE finds a radical solution to materialize the ceteris paribus assumption. During the course of the experiment, all other things concerning the environment are simply removed. Excluding resource consumption, other problems are artificially eliminated from the selective environment. Thus, organisms are equal by virtue of the complete absence of other environmental problems.
Given that these conditions are realized in the LTEE, the expectation is that it is possible to directly observe FIOT occurring in bacterial populations.
7.9 Main Results of the Long-Term Evolutionary Experiment in Light of My Account
Results from the LTEE show that, when the availability of resources is kept stable over the experiment, FIOT occurs with respect to two biological functions: rapid resource consumption and resource consumption with low production cost (Marshall, Malerba et al. Reference Marshall, Malerba, Lines, Sezmis, Hasan, Lenski and McDonald2022).
From a structural perspective, what are the variations of organic traits that enable this better functioning in descendants? One evident phenotypical difference is that descendants are bigger in size. However, the reason for this is not obvious: cell size might have been a target of selection, or might merely be correlated with other traits conferring a better relative fitness (Lenski and Travisano Reference 80Lenski and Travisano1994). It also cannot be ruled out that the increase in size took place due to stochastic events in the presence of a ‘left wall’ of minimal size. An analogous argument at a macroevolutionary scale has been proposed by Stanley (Reference Stanley1973) and Gould (Reference Gould1996). At a microevolutionary scale, this would mean that it is impossible for bacteria of the LTEE to become smaller due to structural constraints.
Interestingly, with respect to the two biological functions, different outcomes have been obtained by Vasi, Travisano et al. (Reference Vasi, Travisano and Lenski1994). Here, the specific environmental problem was constant in the sense of being fundamentally the same: consume resources. However, in the protocol, researchers chose to alternate feast and famine in order to simulate seasonal variations. With this protocol, FIOT occurs with respect to the function of quick resource consumption, but not with respect to the function of resource consumption with low production cost. Although these results are difficult to interpret, they seem to suggest that, with respect to energy efficiency, some condition for FIOT was lacking in the experimental protocol of Vasi, Travisano et al. (Reference Vasi, Travisano and Lenski1994).
Finally, I would like to add a note about the novel function of citrate consumption. As I have previously explained, this concerns the appearance, by random mutation, of an innovative way of solving the environmental problem of resource consumption. Should we consider this an instance of FIOT? Strictly speaking, there is no improvement in the simple fact that some organisms happen to have a new functional trait. However, if this new way of ‘making a living’ confers a relatively better fitness to some organisms, we can affirm that this trait’s emergence by random mutation is potentially the first step towards FIOT.
8 Candidates for Global Progress
8.1 Releasing the Suspension of Judgement
As seen in the previous sections, the notion of local progress presented in this Element has been grounded in organic value and then refined as FIOT. The notion has also been confronted with empirical evidence, both by considering cases from palaeontology and considering the results of experimental evolution, namely the LTEE.
At this point, one might be convinced by the refined notion of FIOT but still wonder if that is all there is to say about progress. Should we be satisfied by this narrow notion of local progress? We might want organic progress to have more grandeur, finding it unsatisfactory that organic goodness is only relative to specific organisms and local environments. Those who are disappointed by the narrow scope of FIOT may wish to find a feature that would be good for living beings in general. This would allow the axiological comparisons to become meaningful among organisms belonging to different lineages, no longer restricted to ancestors and descendants.
To see whether it is possible to satisfy the ‘global progressivist’ reader, we must release the suspension of judgement announced at the start of the Element and finally tackle the issue of global progress. At the end of Section 2, I provisionally conceded to Gould that, due to mass extinctions, it is not straightforward to argue for global progress. It is now time to question this statement. To do so, I shall begin by characterizing an issue that is tightly related to global progress: the issue of macroevolutionary trends.
8.2 Macroevolutionary Trends
When examining empirical datasets in relation to diversifying groups of lineages, palaeobiologists may talk of large-scale evolutionary trends, also called macroevolutionary trends. What are these? I shall here provide a short overview of this topic, trying not to be drawn into technicalities.
Let us consider some organic trait, say organism size in a lineage, considered as a given slice of time. The population that we are studying of course presents variations in organism size. But it is always possible to establish an average, which in statistical terms corresponds to the mean size. With respect to this variable, we can speak of an evolutionary trend towards increasing if there is an increase of the mean over the given slice of time. The same would apply with respect to a trend towards decreasing. More generally, McShea (Reference McShea2000) suggests defining a trend as a ‘change in some summary statistic for a state variable’.
I have here taken the example of organism size, known in the literature as the Cope-Depéret rule (Butterworth, Rogers et al. Reference Butterworth, Rogers and Wills2025). However, the same definition of trend can be applied to whatever variable we can observe in a lineage over a slice of time, for example, degree of differentiation of parts and rate of energy usage.
Moreover, other group statistics than the mean are also important for describing a trend, namely the behaviour of the minimum and the maximum. In the case of size, these would correspond to the minimum and maximum size values observed in a population. Thus, trends can be described by looking at the behaviour of the mean, the minimum and the maximum. To show how different trends can look, it is possible to represent the changing variable on the horizontal axis and time in the vertical axis, as shown in Figure 9.
Figure 9 Different kinds of trends following the specific behaviour of the minimum, adapted from figures kindly shared by McShea. The changing variable is represented on the horizontal axis, while time is represented on the vertical axis.
Both A and B show trends to increase in the mean and the maximum. However, the behaviour of the minimum differs between the two cases. In case A, the minimum sees a significant increase, whereas in case B the minimum, although slightly varying, stays close to the initial value. These two peculiar ‘shapes’ of the trends represent two extreme cases, where the behaviour of the minimum is particularly clear. Of course, in empirical datasets, trends might show a much more nuanced behaviour than in A and B. For example, the minimum could see an increase, but this might be much weaker than in case A.
Until now, I have voluntarily introduced trends in descriptive terms, that is, how group statistics behave. However, another matter to consider is how trends should be accounted for, which I will now discuss.
8.3 Accounting for Macroevolutionary Trends: Internal, External and Undriven Mechanisms
A simple example might help introduce the difference between describing and accounting for a trend. Imagine me telling someone that in the last year I have observed an increase in my body mass index (BMI) from 20 to 25 kg/m2. Thus, my average BMI has seen a trend towards increasing over the year. This information alone does not give any clues about what accounts for the trend in terms of causal mechanisms. The reason might be that I have added muscle mass. It might also be that I have been stressed and have eaten irregularly, or that I was pregnant, thus the BMI increase was expected. Or, maybe, the trend should be accounted for by some combination of these factors. With this example in mind, let us now go back to evolutionary issues.
Different causal mechanisms can lie behind macroevolutionary trends. I suggest conceptually distinguishing a few big ‘families’ of causal mechanisms following McShea (Reference McShea1991), who distinguishes internal, external and undriven mechanisms.
First, it is possible to appeal to some internal mechanisms to account for a trend in a variable. This would mean that the existence of a trend is ascribed to some kind of ‘force’ internal to evolution, a spontaneous tendency to increase. For example, to explain complexity increase (intended as differentiation in parts), McShea and Brandon (Reference McShea and Brandon2010) argue for such an internal mechanism, a spontaneous tendency of biological systems to become complex over time (see further for more details on the Zero-Force Evolutionary Law, ZFEL).
Second, external mechanisms refer to a specific external force driving the increase in the variable. Typically, the invoked external mechanism is natural selection, that is, the selective environment. Consider again the example of size increase. Accounting for this trend by invoking natural selection would mean claiming that the increase in size is related to some fitness advantage. As bigger organisms are advantaged with respect to the selective environment, they come to be more represented over evolutionary time. An analogous argument could be used concerning complexity increases (see, for example, Bonner Reference Bonner1988).
Finally, a third possibility is to explain a trend by appealing to undriven mechanisms, as Stanley (Reference Stanley1973) does to account for an increase in size over evolution. The key idea is that a lower bound is present at the starting point: it would not be possible for an organism to be much smaller than bacteria. Increases and decreases in size occur equally often, but there is a ‘left wall’ that blocks the spread to the left, so the trend is towards the right (bigger sizes). Inspired by Stanley, Gould (Reference Gould1996) has popularized this passive/undriven mechanism by applying it to another trend: the increase in complexity over the history of life. As it is not possible for an organism to be much simpler than bacteria, the ‘left wall’ blocks the spread to the left. Here, Gould (Reference Gould1996) introduces the model of a drunk person walking. Imagine a drunk person leaving a bar. They stand on the sidewalk in front of the bar, a wall on their left and a gutter on their right. Suppose that they move in a single line only and that they have a 50% probability of stepping towards the left wall and a 50% probability of stepping towards the gutter. Now, let the drunk person stagger at random. What will happen? As Gould puts it:
In a system of linear motion structurally constrained by a wall at one end, random movement, with no preferred directionality whatever, will inevitably propel the average position away from a starting point at the wall. The drunkard falls into the gutter every time, but his motion includes no trend whatever toward this form of perdition. Similarly, some average or extreme measure of life might move in a particular direction even if no evolutionary advantage, and no inherent trend, favor that pathway.
This model is also known as diffusion from the left wall. Gould makes clear that this model accounts for the existence of a trend without supposing that the variable brings an evolutionary advantage and without supposing an inherent tendency to increase. Therefore, in Gould’s view, no external mechanisms or internal mechanisms are needed to account for the trend in complexity, as it is sufficient to interpret it in terms of passive diffusion from a left wall.
8.4 Only Natural Selection Could Allow Us to Speak of Progressive Trends
As we have seen, a trend towards increasing in some variable can be accounted for with different causal mechanisms: internal, external and undriven mechanisms. Why is this relevant to the issue of global progress? The reason is that, following the causal mechanisms invoked to account for a macroevolutionary trend, we might be led to different positions on whether the trend should be considered as progressive or not. In fact, between the three kinds of mechanisms, only the external mechanism of natural selection would allow us to talk, strictly speaking, of progressive trends.
Consider Gould’s argument of the drunk person model. By looking at the trend towards the right (either of the drunk person or of the biological variable), we are led to see a directionality: the average and maximum seem to go in one preferential direction, that is, increase. But the cause behind this directional trend, according to Gould, has nothing to do with the fact that it is good for organisms to be complex. Keeping in mind my analysis in the previous sections of the Element, we can here read the term ‘good’ in the sense of organic value, that is, goodness with respect to the environment. Gould’s argument can thus be understood in this way: as undriven mechanisms have nothing to do with organic goodness, it makes no sense to speak of a betterment over time.Footnote 20
The same goes for trends caused by an internal mechanism. A spontaneous tendency might have nothing to do with organic goodness. It could just be an intrinsic property of biological systems, as argued in the case of complexity by McShea and Brandon (Reference McShea and Brandon2010). And again, if it is not a question of organic goodness, it makes no sense to refer to betterment.
However, if the cause of a trend is an external mechanism like natural selection, it might be possible to characterize the trend as progressive. By discussing local progress, we have seen the conceptual links between the notion of organic value and natural selection. If an organic property confers goodness relative to some characteristics of the environment, and if this property sees an increase over evolutionary time, it may make sense to refer to some organic betterment over time.
To sum up, to characterize the trend as progressive, we should be sure, without ambiguity, that the causal mechanism behind the trend is natural selection, while the other mechanisms can be set aside. But this might not be a simple matter.
8.5 Disentangling Mechanisms Behind Trends
By reading this section (and particularly looking at the extreme cases presented in Figure 9), we might be led to think that disentangling mechanisms behind macroevolutionary trends is an easy task. After all, the trend represented in A seems to be actively pushed by something, and this something might be natural selection. On the other hand, the trend represented in B might be easily accounted for with the undriven mechanism described by Stanley and Gould. However, things are, in fact, much trickier than this because at least two problems arise when we attempt to disentangle the mechanisms behind macroevolutionary trends.
First, as evoked earlier, empirical trends ‘in real life’ may present a much less clear form than cases A and B. Intermediate cases, such as trends where the minimum seems to show a slight increase, can be hard to interpret. In such cases, we might, for example, be unable to say whether the minimum is actively driven (although weakly driven) or whether the trend can be accounted for by undriven mechanisms. Palaeobiologists have tried to develop statistical tests for this aim. For example, McShea (Reference McShea1994) proposes three tests to distinguish passive from driven trends. However, despite the remarkable utility of these tests, it is hard to distinguish passive trends from weakly driven trends because of the behaviour of the minimum, which behaves similarly in both cases (McShea Reference McShea, Ruse and Travis2011).
Second, suppose that we have an empirical dataset corresponding to the clearcut shape A. Despite the first intuition, nothing guarantees that the ‘driving force’ or ‘bias’ in A corresponds to the selective environment. For example, the driving force could be some kind of developmental channelling, or even a spontaneous mechanism such as the one described by the ZFEL (McShea and Brandon Reference McShea and Brandon2010). Thus, even in the case of the clearcut trend’s shape A (which would seem to be one of the simplest and most easily tractable), it is not straightforward to draw conclusions on the mechanisms behind a trend. Just as in the example of the BMI increase, we cannot exclude the possibility that the trend should be explained by a combination of different causal mechanisms.
Of course, we can hope that the advances in palaeobiological studies might shine some light on these two problems. For example, a recent proposal was advanced by Butterworth, Rogers et al. (Reference Butterworth, Rogers and Wills2025), who elaborate a model for distinguishing driven or passive trait evolution in the presence of boundaries (for example, biomechanical constraints). The application of this model to two empirical datasets suggests that there is a complex interplay between evolutionary processes: selection (preferentially operating in one direction) and passive diffusion. Interestingly, the paper by Butterworth, Rogers et al. (Reference Butterworth, Rogers and Wills2025) suggests that these two evolutionary forces can overlap and act in opposition.
The further development of this kind of study might lead to improved methods to disentangle the causal mechanisms behind trends. Such a clarification could help in understanding in which cases we could speak of progressive trends, that is, trends where the main driving force can be clearly identified as natural selection.
8.6 Candidates as Criteria for Global Progress
Now that I have characterized macroevolutionary trends and the question of the causal mechanisms has been addressed, let us look at the main criteria of global progress discussed in the literature. A useful overview of these criteria has been provided by Rosslenbroich (Reference Rosslenbroich2006). As it is impossible to present them all, I will here briefly present six.
(I) Morphological complexity. We have already encountered this criterion while briefly presenting Darwin’s theses on organic progress. In addition to competitive highness, Darwin thought that another criterion of progress could be identified: the differentiation and specialisation of parts and organs, that is, morphological complexity. This is one of the most classic propositions of global progress, corresponding to an (ill-defined) intuition: as ‘everybody knows’ (McShea Reference McShea1991), evolution goes from simple to complex forms. The usual rationale for claiming that complexity is a criterion for global progress is the evolutionary advantage that this property would confer to organisms. Natural selection would thus be the driving force that produces the increase in degrees of differentiation.
(II) Hierarchical complexity (levels of nestedness). This criterion refers to the evolutionary steps generally labelled as ‘major transitions in evolution’ (Maynard Smith and Szathmary Reference Maynard Smith and Szathmary1995), that is, the increase in the levels of nestedness from bacteria to eukaryotic cells, then to multicellular individuals and finally to colonies and societies. In the paper we examined in Section 3, Dawkins seems to be sympathetic to this kind of criterion of global progress (Reference Dawkins1997, pp. 1019–1020).Footnote 21 From an empirical standpoint, an increase in the level of nestedness is one of the most well-documented evolutionary trends (McShea Reference McShea2023, p. 3). But, to speak of a progressive trend, we should say that there is something good about the passage from unicellular to multicellular organisms. Knoll and Bambach (Reference Knoll and Bambach2000) seem to suggest something similar when they speak of a directional pattern of increasing ecological complexity. Referring to the major evolutionary transition, they claim that ‘each megatrajectory has introduced fundamentally new evolutionary entities that garner resources in new ways, resulting in a pattern of expanding ecospace utilization’ (Knoll and Bambach Reference Knoll and Bambach2000, p. 10).
(III) Energy intensiveness. Vermeij argues for this criterion of global progress. The palaeontologist defends the idea that global progress is not disrupted by mass extinctions. For Vermeij, even when these catastrophic events occur, some organisms show good features with respect to environmental hazards. These would be opportunistic small organisms, characterized by high levels of energy in their active phase and capable of switching to a dormant phase in unfavourable conditions (Vermeij Reference Vermeij1987, pp. 394–395). According to Vermeij, the continuity of the adaptive escalation is ensured by these high-energy opportunists, in which ‘the potential of escalation is high’ (Vermeij Reference Vermeij1987, p. 417).
(IV) Autonomy from environment. This criterion has been championed by the evolutionary biologist Bernd Rosslenbroich. This author refers to the fact that, in some organisms, the direct influence of the surrounding environment is reduced so that the organisms are more stable as a system and capable of self-regulation (Rosslenbroich Reference Rosslenbroich2006, Reference Rosslenbroich2014).Footnote 22 Endothermy in vertebrates is an example of this increased relative autonomy from the environment. ‘Relative’ is an important term here, because the author acknowledges that no organism can be completely independent of its environment. Rosslenbroich does not claim that autonomy as a criterion for global progress is necessarily ‘the only solution to the problem of progress, but together with others it may be a promising candidate’ (Reference Rosslenbroich2006). Note that Rosslenbroich’s argument seems to rest on the assumption that ‘autonomy is able to increase the fitness of the individual’ (Reference Rosslenbroich2006, p. 64).
(V) Exploitation of environmental heterogeneity. This recent approach, proposed by Desmond (Reference Desmond2021), is worth mentioning. According to Desmond, we can distinguish two kinds of adaptive processes: adaptation within a local selective environment and adaptation by exploiting novel ecological opportunities (e.g., novel resources or unpopulated habitats). The second feature is labelled as exploitation of environmental heterogeneity (EH). For Desmond, it is possible to distinguish and ‘classify’ organisms based on their degrees of EH exploitation. For example, a motile organism is better than a sessile one, and an organism with a more efficient visual system is better than one with a rudimentary visual system. Even though Desmond admits the difficulty of empirically testing, he believes that the trend is robust and that EH exploitation is a good candidate for global progress.
(VI) Evolvability. The concept of evolvability emerged in the early 1990s, and since then it has become an extremely popular concept in evolutionary biology (Hansen, Houle et al. Reference Hansen, Houle, Pavlicev and Pélabon2023). The term has been used in different ways, for example, referring to the capacity to generate variations or to the ability to exhibit phenotypic plasticity (Crother and Murray Reference Crother and Murray2019). Two features seem to be characteristic of the different uses: evolvability is a dispositional concept and it is related to the notion of variability. Despite the difficulty of clearly defining evolvability, it is possible, as we shall see, to consider this notion as an interesting candidate for global progress.
Extensive discussion of all these criteria requires another work. I have thus decided to adopt the following strategy. First, I will focus on complexity criteria (I and II) and show why, despite having been widely discussed, they are not satisfactory criteria for global progress. Then, partly drawing on Desmond’s proposal, I outline the requirements that a satisfactory criterion of global progress should fulfil and I identify two possible directions of inquiry. Finally, I give a perspective on what I consider to be a promising candidate: evolvability.
8.7 Complexity and Global Progress: The Work of Daniel McShea
Daniel McShea, a palaeobiologist at Duke University, has made a significant contribution towards a coherent understanding of complexity from an evolutionary perspective. In fact, despite having been widely discussed as a criterion for global progress, the issue of complexity is far from achieving consensus. I will try to sum up McShea’s work on complexity in three questions.
First, there is the question of the operational definition. What is complexity? The notion might seem intuitive, but it is not. Second, there is the question of the effective presence of macroevolutionary complexity trends. Depending on what we mean by ‘complexity’, can we document an increase in complexity over evolutionary time? This question refers to the possibility of empirically assessing complexity trends. Third, there is the question of the cause of macroevolutionary complexity trends. Admitting that some kind of complexity has increased, can we disentangle the effective cause of the increase? Let us see how McShea tackles these three questions.
Operational definition of complexity. While discussing complexity, Darwin seems to rely on an intuitive notion of specialization of organs, but no precise and operational definition is given. The same is true for several other scholars (McShea Reference McShea1991). McShea’s work offers a remedy to this problem. A first step is to distinguish horizontal and vertical complexity. Horizontal (or non-hierarchical) complexity is obtained by counting a number of ‘elements’. At different biological levels of organisation (McShea and Brandon Reference McShea and Brandon2010), this counting can be applied to physical parts (object complexity, e.g., number of cell types, number of organelles and number of genes) or processes (process complexity, e.g., component interactions in developmental or physiological processes). We then have vertical (or hierarchical) complexity. It is possible to consider both object complexity and process complexity hierarchically. Vertical object complexity refers to the number of levels of nestedness of parts within wholes. In biology, a classical object hierarchy is the series referring to levels of organization: organelle, cell, organ and organism (McShea Reference McShea1996). Vertical process complexity is the number of levels in a causal specification hierarchy: we might think of different causal steps in the process of the development of an organism. To sum up, it is not possible to talk about an overall complexity with respect to biological entities; we should be able to say which kind of complexity we are referring to between four possible options: horizontal (non-hierarchical) object complexity, horizontal (non-hierarchical) process complexity, vertical (hierarchical) object complexity and vertical (hierarchical) process complexity. As McShea puts it: ‘With complexity defined broadly or vaguely, only rhetorical answers are possible. But with the four types clearly distinguished and operational metrics devised for each, wide avenues of research are opened’ (McShea Reference McShea1996, p. 489).
Effective presence of macroevolutionary complexity trends. McShea investigates this question by looking at the empirical data, mainly concerning Metazoans (multicellular animals) over the Phanerozoic Era. Without going into detail, the empirical evidence seems to show that at least two kinds of trends effectively occurred. First, a trend occurred in vertical or hierarchical object complexity (prokaryotic to multicellular eukaryotes), as shown, for example, by McShea and Changizi (Reference McShea and Changizi2003). Second, for some specific subgroups, trends in horizontal or non-hierarchical object complexity have been documented, as shown by the work of the palaeontologist James W. Valentine (see, for example, Valentine, Collins et al. Reference Valentine, Collins and Meyer1994).
Disentangling causal mechanisms. Should these complexity trends be ascribed to the causal mechanism of natural selection? This leads us to the problem that I have raised earlier. In particular, the work of McShea (along with Robert Brandon) puts forward the possibility that the increase in complexity (intended as differentiation of parts) should be ascribed to an internal mechanism (McShea and Brandon Reference McShea and Brandon2010). According to their Zero-Force Evolutionary Law (ZFEL), parts tend to spontaneously differentiate in the absence of selection, thus engendering an increase in complexity over evolution. ZFEL predicts more complex organisms, but it is not expected that these organisms are better adapted relative to the environment. Thus, the ZFEL perspective would turn the question upside down: complexity does not increase because it is ‘good’ for organisms with respect to coping with their selective environment; complexity increases spontaneously in evolutionary systems. However, this increase implies no betterment in the sense of organic value. On the contrary, sometimes the tendency might be counteracted because, in certain environments, it is ‘good’ for organisms to be simple. The same might hold for transitions in hierarchical complexity: according to ZFEL, it can be claimed that transitions to new levels of hierarchy involve a non-selectionist chance process (Fleming and Brandon Reference Fleming and Brandon2015).
8.8 Requirements for a Satisfactory Criterion of Global Progress
This examination of complexity as a candidate of global progress has been instructive in several ways. The two notions of complexity defined by McShea were in principle promising candidates for global progress because they have been shown to satisfy two requirements. First, they have been operationalized. Second, at least some long-term evolutionary trends have been documented, and these are robust to mass extinctions. We can generalize these two requirements for a viable criterion of global progress in the following way:
Empirical requirement (i). It should be possible to operationalize/measure the feature.
Empirical requirement (ii). It can be shown that the feature reflects a macroevolutionary trend towards increasing, despite mass extinctions.
However, these two requirements are not sufficient to obtain a viable criterion for global progress. The notion of progress requires some ‘getting better’, according to the notion of organic goodness characterized in Section 4. We could thus add a further theoretical requirement for a viable criterion of global progress:
Theoretical requirement (i). It can be argued that the feature is increasing over evolutionary history by virtue of its goodness relative to the environment. This implies that it should be possible to exclude other non-selective causes of increase.
As we have seen, it is impossible to identify natural selection as the main cause of complexity increase. In fact, we cannot exclude some viable non-selective explanations, such as diffusive mechanisms as invoked by Gould or the ZFEL.
We might even mention here the possibility of another requirement, inspired by Desmond (Reference Desmond2021). We may be able to find a feature that is good in most environments of our present world. But would it be good in all possible counterfactual replays of life’s tape? This would add another (highly demanding) metaphysical requirement: the property should confer ‘competitive highness’ in all possible counterfactual replays of life’s tape.
8.9 Identifying Two Directions of Inquiry
We have seen that two notions of complexity, described following McShea, satisfy the first two empirical requirements but not the theoretical requirement. How could we pursue this analysis to find promising candidates for global progress? I think it is useful to point out that this question can be investigated in two directions.
Bottom-up. One possibility is to ‘look at the trends first’, in the same way that McShea has investigated complexity. If the feature satisfies the two empirical requirements, then we can see whether it also satisfies the theoretical requirements (and eventually the metaphysical one).
Top-down. Here the starting point of the analysis corresponds to asking the question: what could be a long-term and general problem related to some feature of the environment that organisms must deal with? If we can find a feature related to ‘solving this problem’, then perhaps this feature could represent a satisfactory criterion for global progress, at least from a theoretical standpoint. Then, once identified, we might proceed to see whether the feature also fulfils the two empirical requirements.
Following the path of McShea, I have examined complexity in the bottom-up direction, and I ended up with the conclusion that this is not a satisfactory candidate for global progress. I suggest that we now tentatively try the top-down direction of inquiry to see whether this strategy could be more successful.
8.10 Drawing on an Insight by Georges Canguilhem
As we have seen in this Element, the local notion of FIOT makes sense only with respect to an environmental problem staying (relatively) constant over a timespan. With respect to the specific problem, organisms are more or less performant, with the supposition of the ceteris paribus clause. By contrast, from the perspective of global progress, we are looking for something different, that is, a long-term and general problem related to some feature of the environment. Yet it seems difficult to think that the countless different forms of life, from a species living in hydrothermal sources to an elephant, could share any environmental feature. And, if we wanted to make this more difficult, to satisfy the metaphysical requirement, the environmental feature should be non-contingent, that is, we can find reason to think it is present in all replays of life’s tape.
To find such a feature, I think it is interesting to refer to the biological philosophy of the French author Georges Canguilhem. Mainly known in the English-speaking world for his contribution to the philosophy of medicine, Canguilhem’s thought contains some extremely interesting insights in terms of conceiving the relationship between the organism and the environment from an evolutionary perspective (Sholl Reference Sholl, Méthot and Sholl2020, De Cesare Reference De Cesare2022). According to Canguilhem, it is important to distinguish two ways in which an organism can be said to be adaptive to the environment. On the one hand, local adaptation consists in a specialization for specific tasks (Canguilhem Reference Canguilhem1991, p. 262). In the context of this Element, we can understand this idea in light of my analysis of FIOT. On the other hand, for Canguilhem, adaptive organisms are those who can deal with the inconstancies of the environment, which are inevitable in the long run. Thus, for organic beings, it is one thing to be good with respect to a specific task, and another thing to be ready for environmental change itself. According to Canguilhem’s terminology, ‘normal’ organisms are those who can cope with a specific local environment, while ‘normative’ organisms possess a different characteristic: promptness in reacting to environmental change.
This concept of normativity is widely debated in the literature on Canguilhem’s thought, see for example the edited volume by Méthot and Sholl (Reference Méthot and Sholl2020). However, for my aim here, this simple insight is sufficient: independent of local specificities, a constant feature of the environment is that it is endlessly changing in an unpredictable way. Paradoxically, in the imaginary replays of the tape of life, something about this seems to be non-contingent: the contingency itself of environmental change.
To sum up, local adaptations (understood as FIOT) would be good organic ‘solutions’ to relatively fixed and stable environmental problems. On the other hand, normativity (in Canguilhem’s terminology) would be a good organic ‘solution’ with respect to an inescapable environmental contingency, which organisms might sooner or later meet over evolution. At this point in my reasoning, the question can now be framed in the following way: how could we better characterize Canguilhem’s concept of normativity, and is it possible to understand this in more contemporary terms?
8.11 Robustness and Evolvability in the Account of Bourrat, Deaven et al. (Reference Bourrat, Deaven and Villegas2024)
Among the criteria of global progress that I have sketched earlier, the one that I have evoked under the heading of evolvability seems to go in the same direction as Canguilhem’s insight. As previously mentioned, evolvability applies to different biological systems at different scales: microevolution, macroevolution, genomes and clades (Bourrat, Deaven et al. Reference Bourrat, Deaven and Villegas2024). Evolvability is a dispositional concept because it does not refer directly to actual features but rather to ‘the broad disposition’ to evolve (Brown Reference Brown2014) in the same way that solubility describes the expected behaviour of a substance and not its actual state (Wagner and Altenberg Reference Wagner and Altenberg1996). The concept applies to high levels of organization, that is, population and/or beyond (Ramsey and Villegas Reference Ramsey and Villegas2024). Evolvability is also necessarily related to variability, that is, the disposition to vary, which should be differentiated by the notion of ‘variation’, referring to the differences present in a population (Wagner and Altenberg Reference Wagner and Altenberg1996). Recent work on this issue deals, for example, with the question of understanding the links between the developmental properties of organisms (individual level) and evolvability as a disposition to variation at higher levels (Ramsey and Villegas Reference Ramsey and Villegas2024).
From my perspective, a recent account of evolvability is particularly interesting within the discussion of global progress, that is, the account presented by Bourrat, Deaven et al. (Reference Bourrat, Deaven and Villegas2024) in their paper ‘Evolvability: Filling the Explanatory Gap between Adaptedness and the Long-Term Mathematical Conception of Fitness’. The authors draw on the propensity interpretation of fitness by Pence and Ramsey (Reference Pence and Ramsey2013), who define fitness in a specific way, that is, the probability distribution encompassing all daughter populations to which an organic entity might give rise. Bourrat, Deaven et al.’s (Reference Bourrat, Deaven and Villegas2024) aim is to build on this framework and improve it conceptually by including considerations of evolvability. I will try to illustrate their reasoning by simplifying their terminology.
Suppose an organism Type1 (we can here read: genotype Type1) is considered in a particular environment, the environment of reference Env1. The adaptedness of this organism is defined as its capacity to survive and reproduce in its environment of reference Env1. However, two other features can be considered relative to this organism: robustness of adaptedness and evolvability.
Robustness of adaptedness is defined as the probability of organism Type1 leaving descendants (of the same type) in a different environment from the environment of reference, for example, Env2. Thus, Type1 proves to have robustness of adaptedness if it can withstand the environmental change from Env1 to Env2. On the other hand, the evolvability of organism Type1 is defined as the probability of leaving daughter populations of a different type (e.g., Type2), either in the environment of reference Env1 or in another environment, for example, Env2. In simpler terms, evolvability here refers to the capacity of Type1 to produce and maintain variation, whether in the environment of reference or in another environment.
Thus, the main point raised by the authors is that evolutionary success is not all about being adaptive with respect to a (relatively) fixed local environment. Evolutionary success should also be considered from a longer-term perspective. Such a perspective would include two features: robustness of adaptedness and evolvability.
8.12 Integrating Robustness and Evolvability within My Framework
It is now possible to try to integrate this perspective into my framework on organic progress. What Bourrat, Deaven et al. (Reference Bourrat, Deaven and Villegas2024) label as adaptedness can be related to my treatment of local progress as FIOT. Consider an organism of Type1. If at least some aspects of its environment (Env1) are relatively stable over a timespan, and if some conditions are present (see the conditions for FIOT mentioned earlier), the outcome of the selective process might be local adaptation of the organism to the specific environment. However, in a longer timespan the selective action of the environment can also present other outcomes with respect to organisms. In fact, in the case of more radically changing environments, evolutionarily successful organisms are those that prove to be robust and/or evolvable (in the sense characterized by Bourrat, Deaven et al. (Reference Bourrat, Deaven and Villegas2024)), rather than those showing local adaptedness.
This perspective on robustness and evolvability, only sketched here drawing on an insight by Canguilhem and on the recent proposal of Bourrat, Deaven et al. (Reference Bourrat, Deaven and Villegas2024), merits further development. I plan to do so in another work. For the reasoning on organic progress outlined in this Element, the key point is as follows: if we choose the top-down direction of inquiry to find a satisfying criterion of global progress, robustness of adaptedness and evolvability seem to be promising candidates from a theoretical standpoint. Do these candidates also satisfy the empirical requirements needed for a convincing notion of global progress? Let us recall the two requirements:
Empirical requirement (i). It should be possible to operationalize/measure the feature.
Empirical requirement (ii). It can be shown that the feature reflects a macroevolutionary trend towards increasing, despite mass extinctions.
At present, these two requirements do not seem to be satisfied by the criterion of robustness/evolvability. This direction of research would greatly benefit from a work similar to McShea’s work on complexity, allowing us to find a way to operationalize and measure robustness/evolvability, ideally in such a way that it is also applicable to extinct organisms. Once this is done, it should be possible to look at empirical cases and investigate whether some effective macroevolutionary trends are found. Even though we are still not there, these are likely the next steps towards the aim of establishing robustness/evolvability as convincing criteria of global progress.
9 Conclusion
This Element began with two main questions with respect to the relationships between organic progress and evolutionary theory:
1) There is no unanimity on discarding organic progress from evolutionary biology. Why? Are there any theoretical arguments for using the normative terms ‘good’ and ‘better’ within evolutionary theory?
2) If there are, can we clarify the idea that some ‘change towards the better’ would be conceptually implied by evolutionary theory?
With respect to the first question, I have claimed that the intuition for organic progress – shared by Darwin and Dawkins – can be related to the notion of organic value that I characterized in Section 4. To answer the teleology-based objection, I argued that it is epistemically legitimate to refer to functional/teleological explanations of living beings. The concept of function implies a polarity, insofar as function only makes sense with respect to dysfunction. I claimed that it is with respect to this polarity that organic value is meaningful. Thus, I answered the first question with an affirmative: there are theoretical arguments to defend the notion of organic value, which opens the possibility of a meaningful notion of organic progress.
Concerning the second question, I began by clarifying what ‘change towards the better’ could mean at a local scale. Drawing on two cases from the palaeontological record, I claimed that the process of adaptation by natural selection can be understood as FIOT, with the assumption of the ceteris paribus clause. In Section 7 I then explored how FIOT can be directly observed in experimental evolution, namely in the LTEE.
Then, in Section 8, I further tackled the second question applied at a global scale. By discussing the causal mechanisms that can lie behind macroevolutionary trends, I made clear that only a mechanism would allow us to label a trend as progressive: the mechanism of natural selection. However, I also underlined how difficult it is to pursue the bottom-up direction of inquiry, that is, to identify an increasing property over evolution, examine the causal mechanisms behind the increase and then identify without ambiguity the action of natural selection. I thus suggested trying out the reverse direction of inquiry, that is, the top-down direction. By adopting this strategy, I turned the question around by asking what a general and long-term environmental problem for organisms would be. This problem may even be present in all imaginary replays of life’s tape (Desmond Reference Desmond2021). After framing the question in these terms, I followed an insight by Canguilhem and finally suggested the criterion of robustness of adaptedness and evolvability, drawing on the framework of Bourrat, Deaven et al. (Reference Bourrat, Deaven and Villegas2024). This criterion, I argued, is promising from a theoretical standpoint and is worth further development to satisfy the empirical requirements, that is, operationalization and documentation of a macroevolutionary trend.
As we have seen, progress is an extremely historically charged notion. It is a confusing notion because it applies to such different domains as the whole of human civilization, scientific theories, technical artefacts, human morality and organic beings. By focussing on organic progress, I have tried to identify theoretical arguments that are untouched by biases related to non-epistemic issues (e.g., subjective evaluations/moral considerations). I have identified the notion of organic value as relating to the polarity function/dysfunction. The legitimacy of organic value can be anchored to the heuristic relevance of the notion of function within biology. Speaking of what is instrumentally good or bad for organisms dealing with an environment does not, I think, threaten the scientific legitimacy of evolutionary biology. And once the pertinence of organic value is accepted, it is possible to draw the consequences in terms of understanding organic progress within evolutionary theory.
Note that I have not claimed that we should all necessarily speak in terms of organic goodness/betterment while dealing with evolutionary issues. Some people are reluctant to do so and prefer to avoid this language – probably for the same reasons that they are uneasy with teleology in biology. However, I have tried to show that, if carefully defined, the notion of organic value provides theoretical arguments to speak of local progress intended as FIOT. In the same vein, at a global scale, it is possible to meaningfully discuss promising criteria of global progress, such as robustness of adaptedness and evolvability.
To conclude, I have tried to make sense of Greene’s perplexity about evolutionary biologists’ attitude towards organic progress. Some evolutionary biologists feel that they cannot live with organic progress because they are afraid that the value component of the notion discredits their scientific legitimacy. This fear may be legitimate given the historically charged notion of progress. But, at the same time, some evolutionary biologists feel that they cannot live without progress. I have tried to show that this need can be unrelated to non-epistemic biases: once we grant the epistemic legitimacy of the notion of organic goodness, local and global organic progress can be discussed without fear of discrediting the scientific status of evolutionary biology. And, in my view, this discussion represents one of the most intriguing conceptual issues relative to evolutionary theory.
Acknowledgements
This Element is the fruit of around a decade of struggling with the notion of progress and evolutionary theory, and especially during my last six years as a Teaching Assistant at the University of Geneva. Given its long genesis, this work has benefitted from the feedback of several colleagues who I wish to thank – although I remain responsible for any error this manuscript may contain. I am indebted to Jean Gayon for having started work on this topic and grateful to Donato Bergandi for pushing me to persevere on the objective. Daniel McShea has been a constant and outstanding source of encouragement and feedback on this work and Marcel Weber played a great role in this story (despite his scepticism towards any progress language!). I warmly thank both for our extremely fruitful exchanges. My colleagues in Geneva and elsewhere have also contributed with excellent feedback and I wish to offer special thanks to Raphel Sholl, Lucie Cauwet, Michal Hadlky, Christian Sachse, Michele Luchetti, Fabrice Teroni, Roberto Keller, Alberto Vianelli, Laurent Reynet, Vincent Bontems, Nicola Bertoldi and Margherita Arcangeli. Warm thanks to Clémence Royer for kindly agreeing to do the drawings for Figure 4 and to Enrica Vigliano for her help with the final version of the figures. I am very grateful to Grant Ramsey for supporting this project and for his patience during the genesis of the Element. Two reviewers have given very constructive and helpful comments to improve the manuscript, and I wish to express my gratitude to them. All my thanks to Laura Witz for agreeing to proofread the final version. And finally, I wish to thank my friends, my family and Thomas for their love and support, without whom this intellectual effort would have been impossible.
Grant Ramsey
KU Leuven
Grant Ramsey is a BOFZAP research professor at the Institute of Philosophy, KU Leuven, Belgium. His work centers on philosophical problems at the foundation of evolutionary biology. He has been awarded the Popper Prize twice for his work in this area. He also publishes in the philosophy of animal behavior, human nature and the moral emotions. He runs the Ramsey Lab (theramseylab.org), a highly collaborative research group focused on issues in the philosophy of the life sciences.
About the Series
This Cambridge Elements series provides concise and structured introductions to all of the central topics in the philosophy of biology. Contributors to the series are cutting-edge researchers who offer balanced, comprehensive coverage of multiple perspectives, while also developing new ideas and arguments from a unique viewpoint.