In the early twentieth century, Aleš Hrdlíčka and others established three requirements necessary to prove a Pleistocene-aged human presence in the New World: (1) artifacts or remains that were clearly human, (2) artifacts in indisputable context (direct association), (3) control over chronology (undisturbed stratigraphy; Adovasio and Page Reference Adovasio and Page2009:99; Haynes Reference Haynes1969; Meltzer Reference Meltzer2015a). These stringent, yet reasonable, requirements were met in 1927 when prominent scientists A. V. Kidder and Frank H. H. Roberts confirmed the association of artifacts with extinct bison remains in Folsom, New Mexico. Soon after, archaeologists discovered what would later be classified as Clovis points associated with mammoth bones at the Dent site in Colorado (Figgins Reference Figgins1933). Starting with these discoveries of fluted points and extinct fauna, the hallmark of the Clovis culture has become the hunting of now-extinct Pleistocene fauna (Kornfeld Reference Kornfeld, Walker and Driskell2007; Meltzer Reference Meltzer2015a). While many archaeologists question the view of Clovis as big-game specialists (Cannon and Meltzer Reference Cannon and Meltzer2004, Reference Cannon and Meltzer2008), there is universal acceptance in the field that Clovis hunters did, at least occasionally, hunt megafauna (Haynes and Hutson Reference Haynes, Hutson, Graf, Ketron and Waters2013; Kornfeld Reference Kornfeld, Walker and Driskell2007; Surovell and Waguespack Reference Surovell, Waguespack and Haynes2009). Understanding the number and nature of Pleistocene megafauna kills/scavenge sites is therefore vital to understanding North America's earliest widespread inhabitants.
Archaeologists have subjected Paleoindian kill/scavenge sites to intense scrutiny, especially so for sites containing proboscidean (Mammuthus sp., Mammut americanum, Cuvieronius sp.) remains (Cannon and Meltzer Reference Cannon and Meltzer2004; Frison and Todd Reference Frison and Todd1986; Grayson and Meltzer Reference Grayson and Meltzer2002, Reference Grayson and Meltzer2015; Haynes Reference Haynes, Willig, Aikens and Fagan1988, Reference Haynes1991, Reference Haynes2002, Reference Haynes, Corona M and Arroyo-Cabrales2007a; Haynes and Huckell Reference Haynes and Huckell2007; Haynes and Krasinski Reference Haynes and Krasinski2010; Haynes and Stanford Reference Haynes and Stanford1984; Hofman Reference Hofman and West2001; Kreutzer Reference Kreutzer1988; Meltzer and Mead Reference Meltzer, Mead, Mead and Meltzer1985; Saunders and Daeschler Reference Saunders and Daeschler1994; Waguespack and Surovell Reference Waguespack and Surovell2003). The skepticism of a human presence at bone bed sites is warranted, since natural death sites can be difficult to distinguish from kill/scavenge sites (e.g., Haynes Reference Haynes1991, Reference Haynes, Pickering, Schick and Toth2007b; Haynes and Klimowicz Reference Haynes and Klimowicz2015; Lyman Reference Lyman1994; Todd Reference Todd, Frison and Todd1987; Todd and Rapson Reference Todd, Rapson, Brugal, David, Enloe and Jaubert1999). This high bar for acceptance, however, potentially rejects true associations and consequently affects both interpretations of Clovis lifeways (Cannon and Meltzer Reference Cannon and Meltzer2004, Reference Cannon and Meltzer2008; Haynes Reference Haynes2002; Surovell and Waguespack Reference Surovell, Waguespack and Haynes2009) and the debate surrounding the cause of megafaunal extinctions in the Americas (for review, see Meltzer Reference Meltzer2015b; Waguespack Reference Waguespack, Graf, Ketron and Waters2013). Accordingly, sites containing the remains of extinct megafauna require close attention to taphonomy and site formation to determine human involvement.
Following Hrdlíčka and William Henry Holmes, archaeologists ask that megafauna kill/scavenge sites meet three requirements: (1) contemporaneity of human and megafauna remains, (2) direct association between human activity and megafauna remains, and (3) evidence for use of megafauna remains (e.g., cutmarks, breakage; Grayson and Meltzer Reference Grayson and Meltzer2002, Reference Grayson and Meltzer2015; Haynes and Stanford Reference Haynes and Stanford1984). Researchers have used these criteria to scrutinize possible megafauna kill/scavenge sites (Cannon and Meltzer Reference Cannon and Meltzer2004; Grayson Reference Grayson1984; Grayson and Meltzer Reference Grayson and Meltzer2002, Reference Grayson and Meltzer2015; Haynes Reference Haynes, Willig, Aikens and Fagan1988, Reference Haynes2002; Haynes and Stanford Reference Haynes and Stanford1984; Surovell and Waguespack Reference Surovell, Waguespack and Haynes2009).
Here we apply these criteria to a previously questioned mammoth kill site, the La Prele Mammoth site. The site's initial excavations (Walker et al. Reference Walker, Frison, Darlington, Reider, Latady and Miller1988) left doubt regarding the association of artifacts with the mammoth remains (Byers Reference Byers2002; Grayson and Meltzer Reference Grayson and Meltzer2002). However, our four seasons of investigations show that the La Prele site meets the three criteria, resolves previous doubts about the site, and contains an undeniable association between evidence of human activity and mammoth remains.
The La Prele Mammoth Site
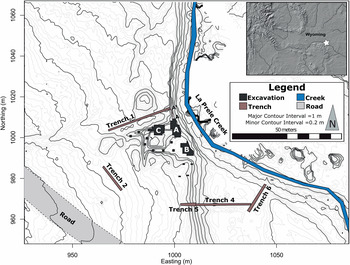
The La Prele Mammoth site (48CO1401) is buried in an alluvial terrace of La Prele Creek, a tributary of the North Platte River, near the town of Douglas in Converse County, Wyoming (Figures 1–2). Initially called the Hinrichs and later the Fetterman Mammoth, the site was discovered in fall 1986 by two Douglas locals, Mike Earnst and William Hinrichs. The following spring a crew led by Dr. George Frison tested the site with an approximately 3 × 4 m block. Frison and crew recovered the partial remains of a mammoth (Mammuthus sp.) and a small artifact assemblage that included a stone tool, a possible hammerstone, and two in situ flakes (Walker et al. Reference Walker, Frison, Darlington, Reider, Latady and Miller1988). The mammoth remains consisted predominantly of axial elements, the other portions having perhaps washed down the creek before the site was identified (Byers Reference Byers2002:Table 1). The artifact assemblage grew when seven flakes were found in the sediments surrounding the mammoth bones in their plaster casts (Byers Reference Byers2002; Walker et al. Reference Walker, Frison, Darlington, Reider, Latady and Miller1988).Footnote 1 The initial excavators concluded that “these flakes would have been deposited at the same stratigraphic level as the ribs, lending credence to the association of the stone biface [flake tool] and flake found during excavation and the mammoth bone” (Walker et al. Reference Walker, Frison, Darlington, Reider, Latady and Miller1988:159). Unfortunately, for the next 30 years the site sat untouched because of a dispute with the landowner.
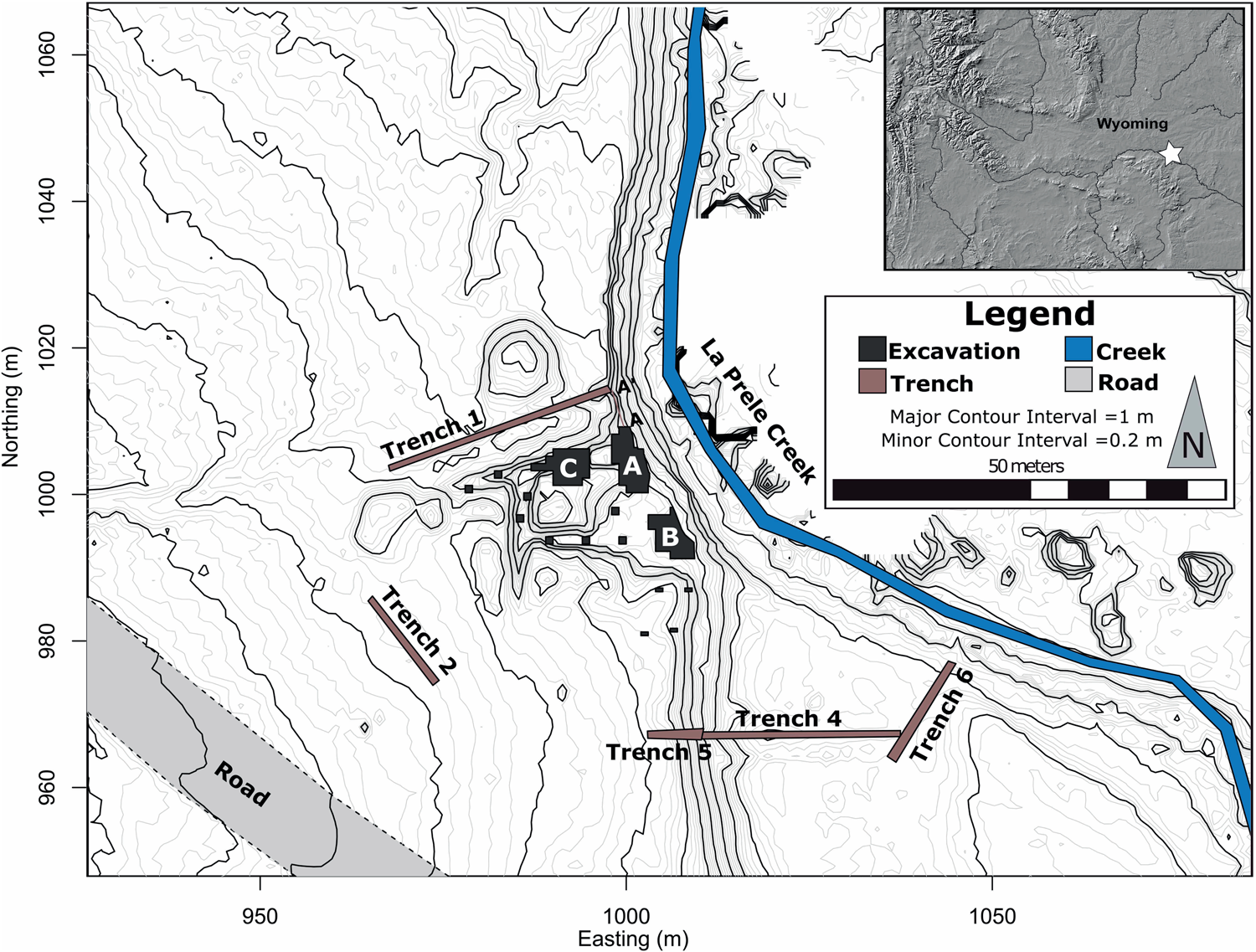
Figure 1. Local topographic map of the La Prele Mammoth site identifying geologic trenches and excavation areas. Major contours are at 1 m intervals, while minor contours are at 20 cm. Note that some of the topography in the immediate area of the excavation is due to backhoe work to expose archaeological strata, and topographic lines were not rendered in the northeast portion of the map due to heavy brush (topographic map created by Randy Haas). (Color online)

Figure 2. Overview of the 1987 and 2015 excavations at the La Prele Mammoth site. The site has been actively eroding since its discovery, and approximately 1.5 m of Block A has been lost. (Color online)
In the intervening years, Byers (Reference Byers2002) cast doubt on the original interpretation of cultural association based on analysis of the 1987 field notes and assemblage. The lack of butchery marks and the close anatomical position of the bones were consistent with a natural death event. Bone weathering patterns suggested that the mammoth remains were partially buried at least twice before being completely covered, leaving the door open for the subsequent deposition of unassociated artifacts (Byers Reference Byers2002:432–435). Finally, slight elevation differences between the bone bed and in situ artifacts found during excavation pointed to separate depositional events for the artifacts and mammoth remains (Byers Reference Byers2002:Figure 11; Walker et al. Reference Walker, Frison, Darlington, Reider, Latady and Miller1988). Byers hypothesized that during one of the partial burial events, artifacts from a later or nearby occupation were washed into the bone bed. This hypothesis was also consistent with the small size and quantity of the artifact assemblage. While none of these analyses is damning on its own, together they cast doubt on human involvement with the mammoth (Byers Reference Byers2002).
A change in landowners allowed us to return to the site in 2014 to address the possibility of a cultural presence there. After relocating the 1987 block, we expanded the original excavations and tested additional portions of the site. Units were excavated in 50 × 50 cm quadrants in 5 cm levels, and all materials recovered in situ, regardless of size, were point plotted using a total station. All sediments were wet screened with 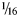 ${1 / {16}}$-inch mesh, and screen matrix was picked in the laboratory after initial picking in the field. Portions of the site contained calcium carbonate concretions large enough to obscure artifacts. These carbonate-rich sediments were treated with acetic acid and picked for artifacts, and gravels were weighed by size. Since an understanding of the site's formation processes was critical to addressing the previous questions, we excavated several geologic trenches that were profiled and dated using radiocarbon and optically stimulated luminescence.
${1 / {16}}$-inch mesh, and screen matrix was picked in the laboratory after initial picking in the field. Portions of the site contained calcium carbonate concretions large enough to obscure artifacts. These carbonate-rich sediments were treated with acetic acid and picked for artifacts, and gravels were weighed by size. Since an understanding of the site's formation processes was critical to addressing the previous questions, we excavated several geologic trenches that were profiled and dated using radiocarbon and optically stimulated luminescence.
Encouraged by the discovery of additional cultural materials and faunal remains, we completed four seasons of excavations at the site (2014 to 2017). In total we have excavated just under 100 m2 along with six geologic trenches (Figure 1). New excavations include the expansion of the 1987 bone bed (Block A), two associated artifact concentrations (Blocks B and C), and 11 test units. We have identified multiple activity areas surrounding the mammoth and recovered more than 1,700 pieces of chipped stone, 500+ bone fragments, and 1,600+ ocher pieces. Here we focus on the evidence supporting the cultural association with the mammoth remains, while future publications will present the details and interpretations of the new artifact assemblages found at the La Prele Mammoth site.
Geologic and Chronological Setting
The La Prele Mammoth is buried within a 7 m alluvial terrace of the La Prele Creek (Figure 2). Terrace sediments span circa 20,000–45,000 cal BP and include alluvial, colluvial, and eolian deposits with intermittent episodes of soil formation (Supplemental Text 1; Supplemental Tables 1–2). The only known archaeological deposits at the locality are in the same stratigraphic horizon as the mammoth, at the lower margin of a buried A-horizon (S-1) within the overbank sediments in which it formed (F-1; Figure 3). Based on texture, sorting, and bedding, we determined that the sediments underlying (F-1) and immediately overlying (F-2) are low-energy overbank flood deposits, not colluvial as previously hypothesized (Albanese Reference Albanese2000; Byers Reference Byers2002). While the S-1 paleosol presents as a pedogenic unit within the excavated portion of the site, it is likely the product of three A-horizons that weld just north of Block A (Figure 3). This is consistent with the mammoth being buried in three stages as hypothesized by Byers (Reference Byers2002) based on bone weathering patterns.
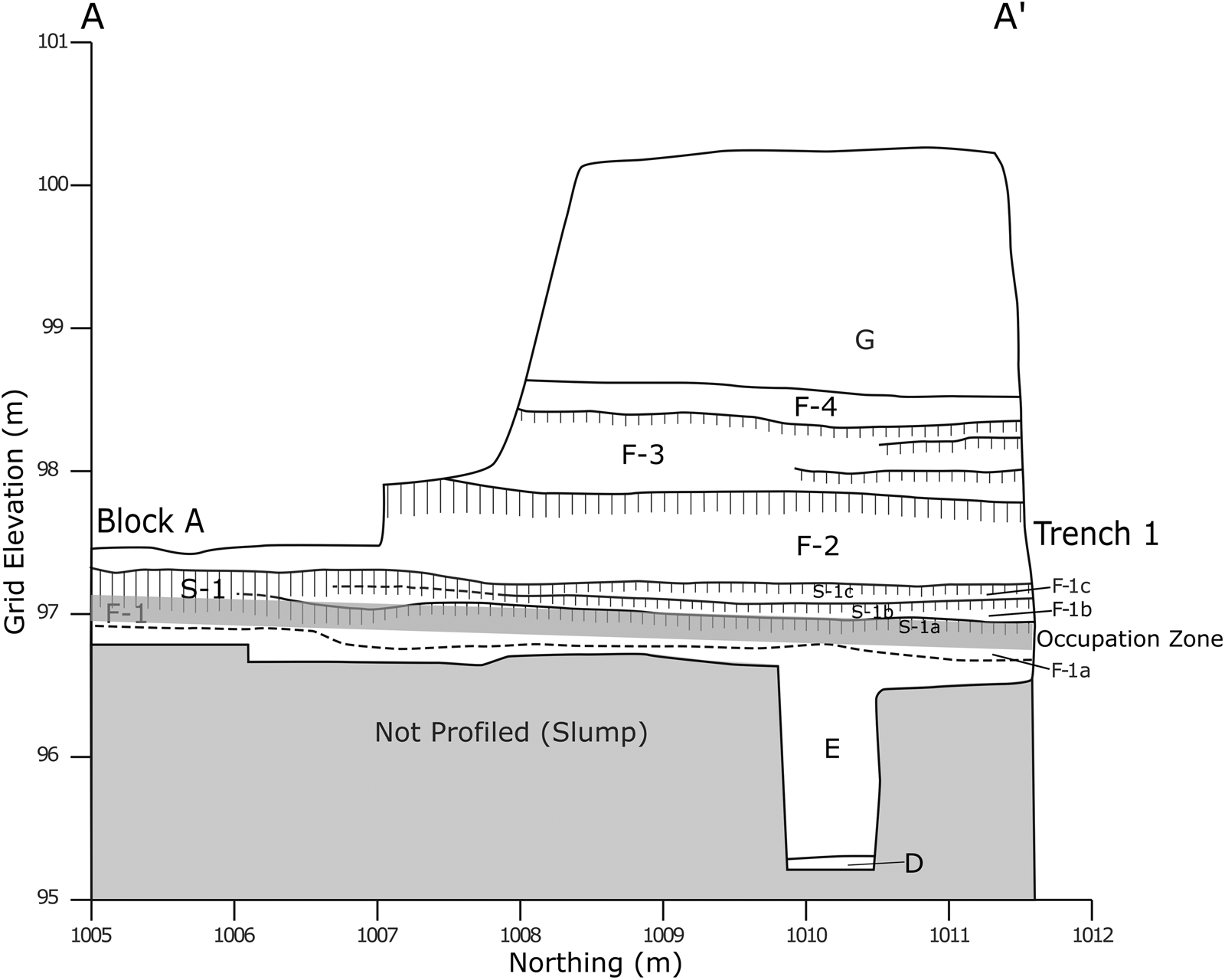
Figure 3. Profile of cleaned terrace face between Block A and Trench 1 in 2015. The archaeological deposits sit at the base of S-1, which welds into a single soil in the center of this profile just north of Block A. Strata descriptions can be found in Supplemental Text 1.
Site dating has been complicated by the presence of young contaminants. Originally, Byers (Reference Byers2002:421) reported two early Holocene dates on the mammoth bone, 8890 ± 60 BP (CAMS-74661) and 9060 ± 50 BP (CAMS-72350; Table 1). While the close range of the dates allowed for some confidence, Byers acknowledged that these dates were young for a mammoth site and could have been affected by contamination. Since 2014, an additional 10 dates from two different portions of the site directly or indirectly date the occupation (Table 1), two of which were recently published by Devièse and colleagues (Reference Devièse, Stafford, Waters, Wathen, Comeskey, Becerra-Valdivia and Higham2018). Dated materials include four bone fragments (two mammoth, two large mammal), two calcined bone samples, two pieces of charcoal, and two samples of soil organic matter. The distribution of the dates confirms the presence of contaminants, as the six dates on mammoth bone produced the youngest as well as some of the oldest dates on the site. We also have one date on calcined bone, 13,997 ± 90 BP (AA109297), that is anomalously old, for which we do not have a good explanation of its age.
Table 1. Summary of Radiocarbon Dates That Directly or Indirectly Date the Occupation.
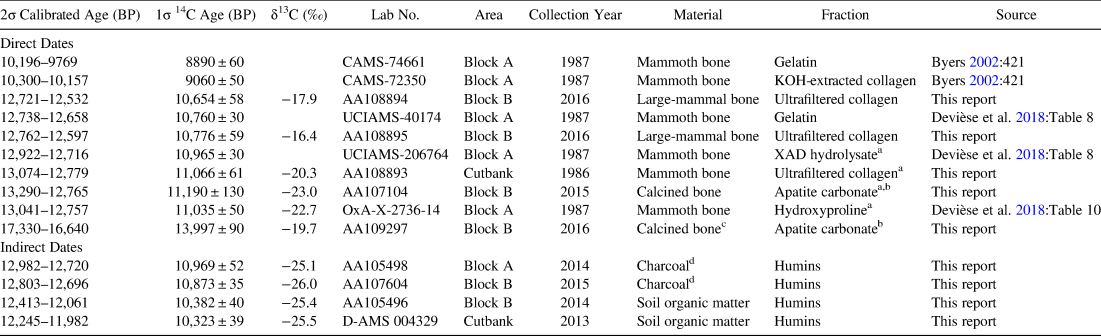
Note: Dates were calibrated using OxCal with the IntCal 13 calibration curve (Reimer et al. Reference Reimer, Bard, Bayliss, Warren Beck, Blackwell, Ramsey, Buck, Cheng, Lawrence Edwards and Friedrich2013). All radiocarbon dates from the site can be found in Supplemental Table 1.
a Dates considered most reliable by the authors.
b Accuracy established by Chatters et al. Reference Chatters, Brown, Hackenberger, McCutcheon and Adler2017.
c Anomalous date.
d Indicates the age of the stratigraphic unit, as charcoal fragments cannot be directly associated with the occupation.
Because contamination of bone collagen almost always produces radiocarbon dates that are too young, we argue that the oldest mammoth bone collagen dates and a calcined bone date from Block B are closest to representing the true age of the site. These four oldest direct dates produce a statistically homogeneous subset determined by pairwise t-tests (Long and Rippeteau Reference Long and Rippeteau1974) after calibration. Those include two dates first reported herein on ultrafiltered collagen from a recovered mammoth rib (11,066 ± 61 BP, AA108893) and a calcined bone date from Block B (11,190 ± 130 BP, AA107104) from a hearth-centered activity area (Mackie et al. Reference Mackie, Surovell, Pelton, O'Brien, Kelly, Frison, Yohe, Teteak, Shapiro and Kapp2020). We also include two dates published by Devièse and others (Reference Devièse, Stafford, Waters, Wathen, Comeskey, Becerra-Valdivia and Higham2018), one hydroxyproline date on collagen from a mammoth vertebra from the 1987 excavation (11,035 ± 50 BP, OxA-X-2736-14), and a sample of XAD-purified protein hydrolysate from the same vertebra (10,965 ± 30, UCIAMS-206764). After calibration and averaging by the Long and Rippeteau (Reference Long and Rippeteau1974) method, our best estimate for the age of the site is 12,846 ± 29 cal BP, placing the La Prele Mammoth site within the Clovis time period as defined by Waters and Stafford (Reference Waters and Stafford2007). Further confirmation of this age estimate comes from two soil organic matter dates on the S-1 paleosol dating the soil formation to a few hundred years after the mammoth's death (Table 1). The discovery of a nearly complete fluted projectile point in the same stratigraphic level as the mammoth approximately 20 m to the southeast of the bone bed confirms the cultural affiliation as Clovis.
Testing Cultural Association
There are a few patterns we expect to see at the La Prele Mammoth site if the cultural activities and mammoth remains are associated. First, there should be a mechanism to account for the vertical movement of materials on-site that would explain the elevation differences between the 1987 lithic artifacts and the bone bed. Second, there should be little to no evidence of horizontal disturbance, as this would make the redeposition of artifacts from a secondary source unlikely. Third, if the cultural materials and mammoth bone are associated, they must have been deposited at the same time and would have been subjected to the same postdepositional processes. Finally, there should be some direct evidence of interaction between the cultural materials and the mammoth remains. If these four lines of evidence are present, we can reasonably conclude that there is an association between the cultural component and the mammoth bone at the site. In order to test these four lines of evidence we look at three components of the site: (1) geologic context, (2) provenience of bone and artifacts, and (3) evidence of direct association.
Geologic Context
The single strongest piece of evidence for questioning a cultural association with the mammoth remains is the elevation difference between the four in situ artifacts and the mammoth bone (Byers Reference Byers2002). Byers noted that this discrepancy could be accounted for by “several geomorphological processes whose signatures are not readily identifiable . . . including bioturbation, frost heaving, and soil compaction” (Reference Byers2002:437). During the last four field seasons we have gained a better understanding of the geologic context and setting of the La Prele Mammoth site through excavations and geologic trenching. The exposure of a larger stratigraphic profile has allowed for a more detailed reconstruction of the Pleistocene surface. This greater understanding of the geologic context of the artifact-bearing deposits is useful for evaluating the first two lines of evidence: the presence or means for vertical and horizontal disturbances on-site.
Bioturbation
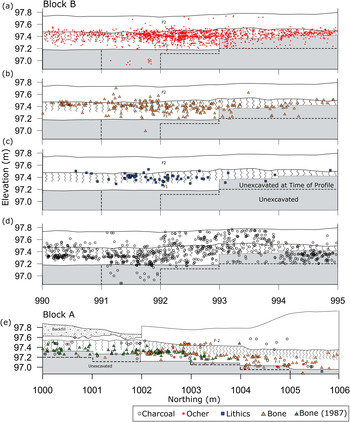
Back plots of in situ artifacts from Block A and Block B show that artifacts are stratigraphically associated with the mammoth remains (Figure 4). In all portions of the site the archaeological material is only associated with upper stratum F-1 and the S-1 paleosol. Although bioturbation had not been previously noted on-site, it is extensive, with visible mottling of the F-1 sediment and S-1 paleosol across the site (Figure 5). Artifact back plots from several site areas show the effects of bioturbation. In Block B, small numbers of artifacts appear as deep as 55 cm beneath the postulated occupation surface, but most materials are spread throughout a 35 cm zone. Based on the relatively small size of the majority of krotovinas (<3 cm), most of the bioturbation appears to be the result of insect and small rodent activity (Figure 5). Given this extreme bioturbation, it is unsurprising that a small number of the artifacts from 1987 were found at slightly higher elevations than the bone bed.
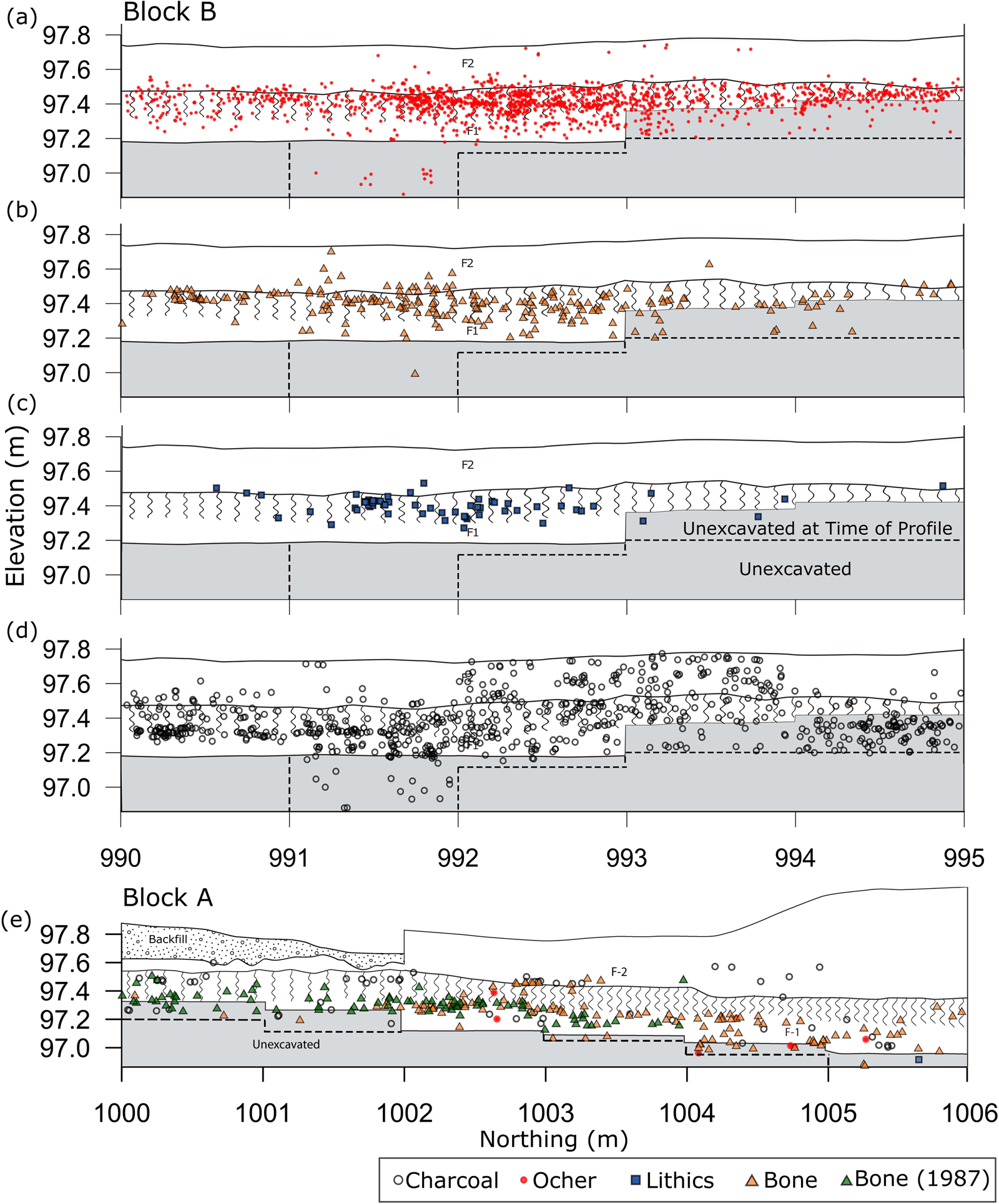
Figure 4. West wall profiles of excavation Blocks A and B with artifact back plots overlain. Deepest extent of excavation within the excavation block east of each profile is indicated by the dashed line. (Color online)

Figure 5. Typical extent of bioturbation with intermixing of the F-1 and S-1 strata. (Color online)
Reconstruction of the Pleistocene Surface
Byers (Reference Byers2002:427) suggested that the artifacts could have been deposited by slope-related processes. This hypothesis was based on the modern ground surface west of the site, which has a gradual slope extending from the site to a nearby strath terrace of the North Platte (Figure 1). However, our larger exposures of the paleosurface show that the Pleistocene surface had relatively little slope. The F-1/F-2 contact drops only approximately 15 cm over 16 m from south to north (N990 to N1006; Figure 4), and from west to east the site drops 20 cm over 21 m (Supplemental Text 1; Supplemental Figure 1). Thus, slopes average just under a centimeter of elevation change per meter. Redeposition of artifacts by slope-related processes is not possible under these circumstances (Fanning and Holdaway Reference Fanning and Holdaway2001; Poesen et al. Reference Poesen, van Wesemael, Bunte and Benet1998).
Based on taphonomic analysis of bone weathering stages, Byers (Reference Byers2002) concluded that the mammoth was likely buried by multiple small flood events. While the S-1 paleosol in the excavation blocks is welded into a single cumulic A-horizon, the S-1 soil separates into three thin A-horizons (Soils 1a to 1c) north of Block A (Figure 3; Supplemental Text 1). Byers (Reference Byers2002) attributes the artifacts’ position higher than the bone as possible evidence for their deposition following one of these small flood events. We propose that the artifacts were shifted through bioturbation, as the lack of slope on the Pleistocene surfaces makes artifact redeposition over long distances unlikely. While we agree that the mammoth was buried in three small flood events, we do not think that those events were responsible for the association between the cultural materials and the mammoth remains.
Assemblage Composition and Spatial Patterns
The presence of bioturbation offers a plausible mechanism for vertical separation between artifacts and the bone bed, but it does not indicate contemporaneity of the human occupation and mammoth remains. The radiocarbon ages from the occupation show that both cultural materials (e.g., calcined bone) and mammoth remains have overlapping ages (Table 1). However, due to an unidentified source of contamination that is younger than the sample, the consistency between dates is by no means conclusive. Instead, we look for indications of contemporaneity using artifact distributions and assemblage attributes. We use three different tests: (1) vertical movement of different materials, (2) identification of artifact clusters, and (3) comparison of lithic attributes between blocks.
Vertical Distribution Analysis
The first test for contemporaneity between cultural and faunal remains examines the vertical dispersal of materials by bioturbation. If the cultural materials and mammoth are associated, they would have been deposited at the same time and been subjected to the same postdepositional processes. Therefore, we should expect similar bone and artifact vertical distributions by bioturbation. For this analysis, point-plotted cultural artifacts (lithics and ocher) are compared with point-plotted bone. Unfortunately, the exact elevation of the mammoth bone bed excavated in 1987 could not be reconstructed with sufficient confidence (within 3 cm) to be included in this analysis.Footnote 2 Only those bones recovered from recent excavations are included, including several larger pieces of mammoth bone (partial rib, tarsal, unidentified large fragments) and many small bone fragments. Although the majority of the new bone assemblage is too small for species identification, many pieces are likely mammoth, so it is used here to represent the mammoth provenience. The slope on the La Prele site is gradual enough that is it unlikely to redeposit artifacts, but it is large enough that raw artifact elevations cannot be directly compared within or between excavation blocks. Instead, all mapped bone and artifact elevations can be standardized to the F-2/F-1 contact, since it is a consistent lithostratigraphic contact that mirrors the topography of the paleosurface. We digitized the F-2/F-1 contact elevation on 17 stratigraphic profiles across the site and subtracted each artifact's elevation from the closest mapped F-2/F-1 contact elevation. The mapped stratigraphic contacts lie within approximately 5 m of each point-plotted artifact/bone/ocher (Figure 6).
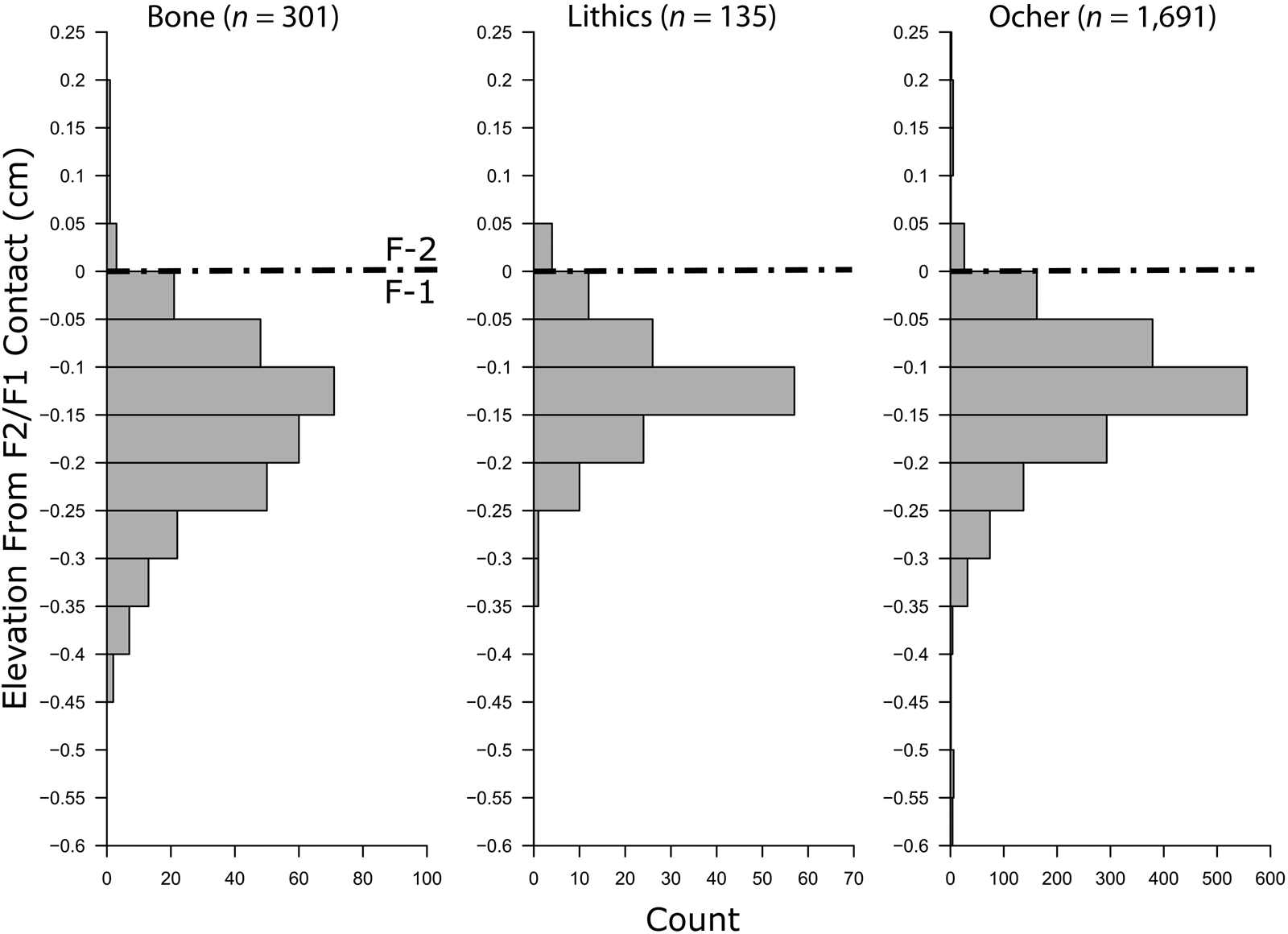
Figure 6. Dispersal of artifacts relative to the F-2/F-1 contact. The dashed line indicates the strata contact at elevation zero.
The distributions of all three material classes are remarkably similar, with modes at 10–15 cm beneath the F-2/F-1 stratigraphic contact. This marks the likely position for the occupation and is consistent with the highest artifact concentrations occurring in or near the lower part of the paleosol (S-1; Figure 4). Both the lithics and ocher are distributed across approximately 35 cm vertically, while most of the bone is spread over 50 cm (Figure 6). We suspect that the bone has a larger distribution because not all the fragments present are associated with the archaeological occupation, although all obvious microfauna remains were excluded from this analysis. In either case, all three distributions are similarly dispersed, suggesting that the artifacts and the majority of bone were deposited at the same time and have been subjected to the same postdepositional processes.
Horizontal Spatial Distributions
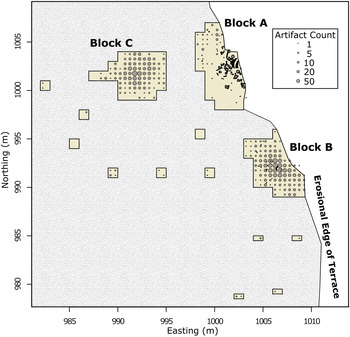
Spatial distributions can also help determine whether artifacts were redeposited on the site by establishing the horizontal spatial integrity of the La Prele deposits. If materials were disturbed or redeposited onto the site, we would expect to see a homogeneous distribution of artifacts across the site from their source (Gregg et al. Reference Gregg, Kintigh, Whallon, Kroll and Price1991). Based on the artifact frequency distributions, artifacts are not distributed homogeneously across the site; instead, each block encompasses distinct clusters with areas of low artifact densities separating clusters (Figures 7–8). This heterogeneous artifact distribution suggests that the horizontal spatial distributions on-site are relatively intact and that redeposition is not responsible for the artifacts found in the bone bed.
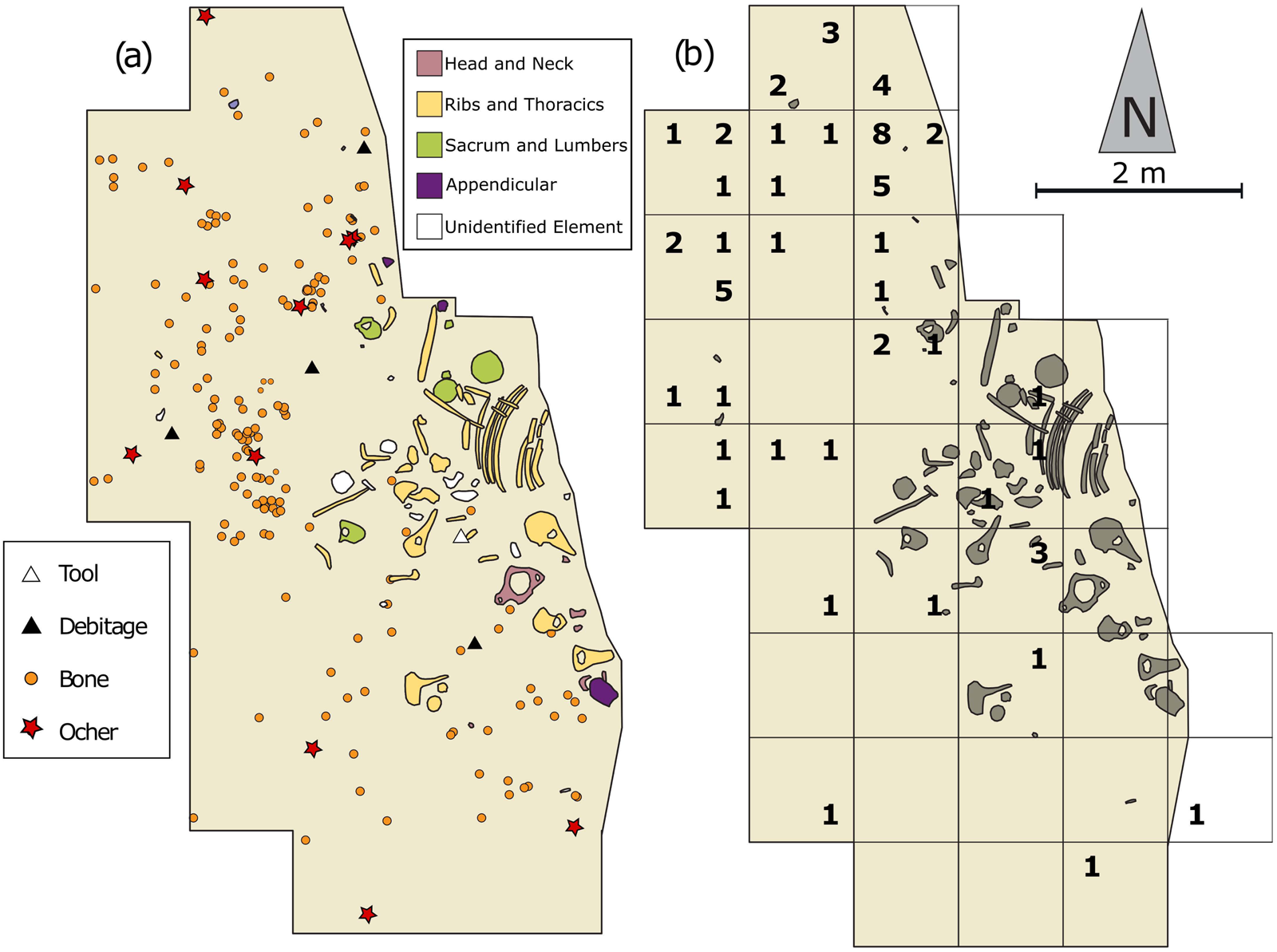
Figure 7. Map of Block A including 1987 and recent excavations: (a) artifact point plots and mammoth elements coded by portion; (b) lithic totals as of 2019 by 50 × 50 cm quadrant. (Color online)
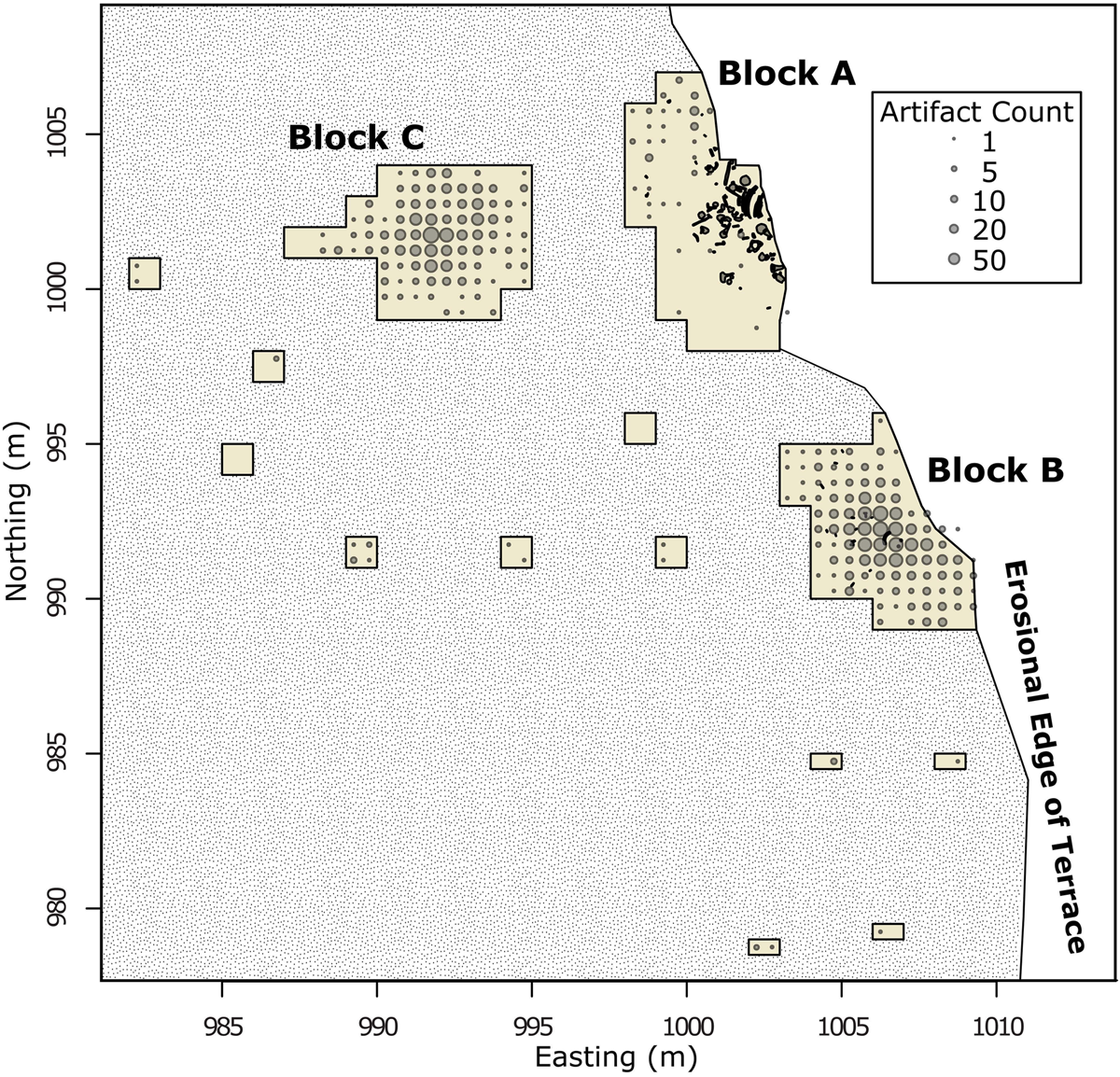
Figure 8. Frequency of lithics across the excavation blocks at the La Prele Mammoth site. Large piece-plotted bone is indicated including the 1987 bone bed in Block A. Block C screen processing is ongoing, so counts should be treated as preliminary. (Color online)
We can also test whether the artifacts are spatially associated with the mammoth remains in Block A (Figure 7). A simple test is to calculate the average distance of lithic artifacts from the closest large faunal element and compare that with the average distance seen in a random (null) artifact distribution (Giusti et al. Reference Giusti, Tourloukis, Konidaris, Thompson, Karkanas, Panagopoulou and Harvati2018). If artifacts are associated with mammoth remains, we would expect the distance between the observed artifact distribution and the closest bone elements to be smaller than the distance from the randomly generated artifact locations to mammoth elements. To run the analysis, we took the number of artifacts recovered as of 2019 from Block A (n = 63) and randomized their location within their respective 50 × 50 cm quadrants (Figure 7). The same number of artifacts were randomly plotted across the entire excavation block to create random distributions. The average distance of the observed and random artifact distributions to mammoth elements was recorded, and the analysis was repeated. Twenty thousand iterations confirm that the observed assemblage (x̅ = 49.6 cm) was significantly closer (p = .0454) than the randomly distributed artifacts (x̅ = 60.9 cm) to faunal remains. This supports the hypothesis that artifacts were deposited in association with the faunal remains and indicates that horizontal distributions are relatively intact on-site.
The close spatial association of the mammoth remains and artifacts is also significant as the La Prele Mammoth site sits at an unremarkable location. La Prele Creek flows continuously for more than 40 km from its head in the Laramie Range to its confluence with the North Platte 1.6 km downstream of the site, and there are no obvious point attractors adjacent to the site for human or mammoth occupation, such as a spring. While we do not have a strong grasp of the age on the uppermost terrace deposits, we do know that the terrace continued to aggrade at least through the early Holocene and ceased before a cut and fill located above the mammoth was formed in the middle Holocene (Supplemental Text 1). No archaeological materials, other than Clovis, have been identified anywhere else in this sequence. It appears that the only human occupation of this locality occurred in the same stratigraphic position and in close spatial proximity to the mammoth. The chances of this spot on the landscape being the location of a natural mammoth mortality followed shortly by a Clovis occupation to never be used again seem exceedingly unlikely (Mackie Reference Mackie2019:52–87), especially at a time when human populations in Wyoming were very low (Kelly et al. Reference Kelly, Surovell, Shuman and Smith2013; Kornfeld et al. Reference Kornfeld, Frison and Larson2010). The most parsimonious explanation is that the mammoth remains and artifacts are associated.
Artifact Assemblage Attribute Analysis
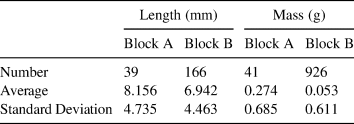
A second source for redeposition of artifacts is the possibility that one of the close artifact clusters is the source of the Block A artifact assemblage. Although slope analysis and the gap in artifact density between areas suggest that this is not the case, we can test this hypothesis using the size of a sample of debitage from Block A (n = 41) and Block B (n = 926; Table 2). Since matrix picking and lithic analysis of Block C is ongoing, its artifact assemblage was not included in this analysis. If the materials from Block B were redeposited to Block A, we would expect the Block A lithics to be smaller than the Block B lithics, as lighter lithics require less energy to be moved and would be preferentially transported (Byers et al. Reference Byers, Hargiss and Finley2015; Sheppard and Kleindienst Reference Sheppard and Kleindienst1996). In fact, the opposite pattern is apparent, as a sample of Block A flakes are (x̅ = 0.27 g, s = 0.685, n = 41), on average, significantly heavier (t = 2.066, df = 40, p = .0226, one sample) than Block B debitage (x̅ = 0.053, s = 0.611, n = 926).
Table 2. Summary of Block A and Block B Debitage Dimensions.
Direct Evidence of Association
The final test to establish an association between the cultural activities and the mammoth remains is direct association. The lack of traditional indications of butchery, such as cutmarks, does not necessarily support a natural mammoth death. Ethnographic/ethnoarchaeological studies have shown that cutmarks may not be produced during carcass processing or could be obscured by weathering (e.g., Crader Reference Crader, Clutton-Brock and Grigson1983; Frison Reference Frison1989; Haynes Reference Haynes, Willig, Aikens and Fagan1988, Reference Haynes1991:303; Haynes and Klimowicz Reference Haynes and Klimowicz2015). In fact, other accepted Clovis proboscidean kill/scavenge sites also lack clear butchery marks (Frison and Todd Reference Frison and Todd1986; Hofman Reference Hofman and West2001; Sanchez et al. Reference Sanchez, Holliday, Gaines, Arroyo-Cabrales, Martínez-Tagüeña, Kowler, Lange, Hodgins, Mentzer and Sanchez-Morales2014). Additionally, the La Prele mammoth lacks many elements, including most of its appendicular elements, and many of the present faunal remains are weathered, obscuring any possible surface cutmarks (Byers Reference Byers2002). It is unclear whether the absent elements are missing because they washed down La Prele Creek from the truncated portion of the site or whether they were moved to a yet unexcavated portion of the terrace. We assume that some portions of the mammoth were lost to site erosion based on the presence of rib and enamel plates on the terrace riser when the site was discovered in 1986. Instead we look to alternative methods to identify direct associations, specifically blood residue and lithic microwear analysis. Both analyses were run on the stone tools recovered from Blocks A and B, including the 1987 flake tool, a cobble tool, a Clovis point, and a set of flake tools (Figure 9). If the tools were used to butcher the mammoth, we would expect positive proboscidean blood residue results and microwear characteristics consistent with animal butchery.

Figure 9. La Prele Mammoth site tool assemblage from (a) a test unit, (b) Block A, and (c–l) Block B: (a) partial Clovis point from approximately 20 m south of Block A; (b) flake tool recovered from within bone bed in Block A in 1987 excavations (FS 16); (c) cobble chopper (FS 354); (d) flake tool with graver (FS 1326); (e) flake tool with graver (FS 1547); (f) spokeshave/flake tool (FS 2970); (g) conjoined flake tool fragment (FS 3840 and FS 4399); (h) partial flake tool (FS 3055); (i) flake tool fragment (FS 3013); (j) flake tool fragment (FS 1349); (k) conjoined flake tool (FS 1917 and FS 1922); (l) partial burned endscraper (FS 4624). (Color online)
Protein Residue Analysis
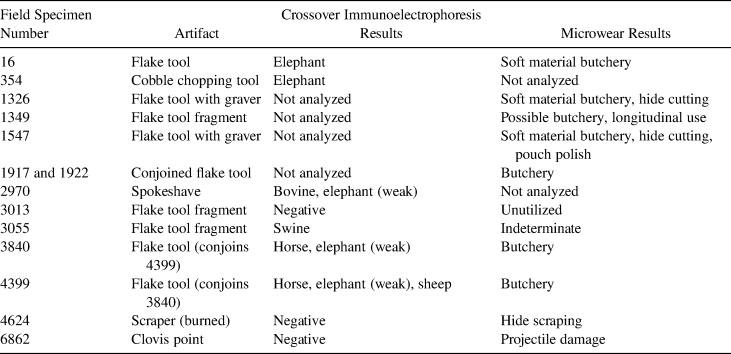
Protein blood residue analysis identifies plant and animal residues on flaked- and ground-stone lithic artifacts using immunological responses (Allen et al. Reference Allen, Newman, Riford and Archer1995; Gerlach et al. Reference Gerlach, Newman, Knell and Hall1996; Henrikson et al. Reference Henrikson, Yohe, Newman and Druss1998, Reference Henrikson, Byers, Yohe and De Carlo2017; Hyland et al. Reference Hyland, Tersak, Adovasio and Siegel1990; Kooyman et al. Reference Kooyman, Newman, Cluney, Lobb, Tolman, McNeil and Hills2001; Petraglia et al. Reference Petraglia, Knepper, Glumac, Newman and Sussman1996; Shanks et al. Reference Shanks, Kornfeld and Hawk1999; Yohe et al. Reference Yohe, Newman and Schneider1991). This method identifies the animal species artifacts may have contacted, although there are questions about its reliability (Eisele et al. Reference Eisele, Fowler, Haynes and Lewis1995; Fiedel Reference Fiedel1996). In this case, we used crossover immunoelectrophoresis (CIEP) to test archaeological residues using procedures developed by the Royal Canadian Mounted Police Serology Laboratory (1983). The samples were analyzed at the Laboratory of Archaeological Sciences located at California State University, Bakersfield. Reactions that produced particularly prominent or light precipitation lines were identified, respectively, as “strong” or “weak.” Tests were repeated multiple times, and soil samples were tested to rule out any false positives. Eight family-level antisera (pronghorn antelope, bovine, Camelidae, Cervidae, Elephantidae, equine, Caprinae, and porcine) were tested on nine stone tools (Figure 9). Two tools and the Clovis point (Figure 9; Table 3) did not return a positive result with any antisera. Six other tools tested positive with at least one antiserum, including elephant, bovine, horse, swine, and sheep (Table 3). Multiple positive reactions for one flake tool could indicate repeated use of the same artifact or the species of materials used for hafting. Weak responses to elephant antiserum should be viewed with caution, as they could be the result of false positives, soil contamination, or protein degradation. While we do not have corollary evidence for all the species identified with CIEP, the strong reaction of two tools to proboscidean antiserum provides direct evidence of interaction between cultural materials and the mammoth remains (Table 3). Notably the two positive proboscidean reactions came from the flake tool recovered from within the bone bed in 1987 (Figure 9b) and the cobble chopping tool found in Block B (Figure 9c). The La Prele Mammoth cobble chopper is consistent with expedient cobble tools that have been found at other Clovis proboscidean kill sites (Frison and Todd Reference Frison and Todd1986; Haury et al. Reference Haury, Sayles and Wasley1959; Haynes and Huckell Reference Haynes and Huckell2007), presenting another line of evidence for human use of the site.
Table 3. Summaries of Crossover Immunoelectrophoresis and Microwear Lithic Analysis on Block A and Block B Stone Tools.
Lithic Microwear Analysis
Since tool morphology is not always consistent with function (Jeske and Sterner-Miller Reference Jeske and Sterner-Miller2015; Knutsson et al. Reference Knutsson, Knutsson, Taipale, Tallavaara and Darmark2015; Odell Reference Odell1981; Rockwell Reference Rockwell2017), lithic microwear studies aim to establish precise designations of tool functions using microscopic scarring patterns left on stone tools (Keeley Reference Keeley1974; Odell Reference Odell1975). Low-powered microscopic (10–120× magnification) use-wear analysis was conducted on 10 tools from the La Prele Mammoth assemblage using a Nikon SMZ800 microscope (Odell and Odell-Vereecken Reference Odell and Odell-Vereecken1980; Tringham et al. Reference Tringham, Cooper, Odell, Voytek and Whitman1974). Microwear patterns from the La Prele tools were compared with a reference collection of tools with known function to identify activities and contact materials consistent with observed scarring patterns (Supplemental Text 2). When possible, contact materials were further refined by the description of polish and striations.
Eight of the 10 flake tools analyzed showed characteristics consistent with butchery of soft materials or hide working (Table 3; Supplemental Text 2; Supplemental Table 3). Two of these tools had corners reworked into graver tips that had scarring patterns and polish on the lateral margins of the flakes; this is consistent with butchery and hide preparation (Figure 9d–e; Supplemental Figures 2–3). One flake tool has notable wear on body arrises consistent with pouch polish, indicating that the tool may have been carried or curated for an extended duration (Figure 9e). The Clovis point was broken on its proximal end likely within the haft as ground basal edges are present (Figure 9a). While microwear analysis cannot attribute a specific species, the presence of wear consistent with butchery is what would be expected for a mammoth butchery site and generally correlates with the CIEP results.
Conclusion
To recap, a true association between cultural materials and the mammoth remains would be supported by (1) a mechanism for vertical dispersal, (2) little evidence for horizontal movement, (3) bone and artifacts showing similar distributions, and (4) evidence of interaction between humans and the mammoth remains. There is support for all four hypotheses based on the geologic context of the site, observed spatial distributions, artifact assemblage attributes, and tests for direct associations between the cultural materials and the mammoth remains. The minimal slope of the paleosurface makes artifact redeposition unlikely, while the bioturbation provides a parsimonious explanation for the elevation discrepancies identified by Byers (Reference Byers2002). The overlapping dates (Table 1), similar vertical distribution of artifacts (Figures 4 and 6), and spatial distributions (Figure 8) support the hypothesis that Blocks A, B, and C were contemporaneously occupied. The presence of distinct artifact clusters with minimal “smearing” by postdepositional processes indicates intact horizontal artifact distributions. The CIEP and lithic microwear analysis are consistent with animal butchery and, in the case of the CIEP results, directly link some artifacts to the mammoth remains (Table 3). Finally, it seems highly unlikely that two closely timed events (a mammoth dying and a Clovis occupation) would occur in such a small time period without being associated, particularly since the locality was never occupied again.
Generally, archaeologists use three criteria to evaluate the evidence for human-killed/scavenged megafauna: (1) contemporaneity, (2) association, and (3) utilization (Grayson and Meltzer Reference Grayson and Meltzer2002, Reference Grayson and Meltzer2015; Haynes and Stanford Reference Haynes and Stanford1984). In the case of the La Prele Mammoth site, contemporaneity is supported by new radiocarbon dates and consistent stratigraphic association. The close spatial association of mammoth remains and artifacts provides a case for a strong association, while the new understanding of geologic setting offers a reasonable explanation for the previously raised doubts. Although this site does not have any traditional indicators of direct use (e.g., cutmarks), the identified tools have tested positive for proboscidean blood antigens and have lithic microwear consistent with animal butchery. We feel that the site rises to the “consonance of data” (Haynes and Stanford Reference Haynes and Stanford1984:220) required to fulfill these three criteria. We propose that the La Prele Mammoth site is at least the sixteenth case of culturally associated megafauna kill/scavenge sites (Grayson and Meltzer Reference Grayson and Meltzer2015).
La Prele Mammoth demonstrates the value of reinvestigation of previously doubted archaeological kill/scavenge sites. Without further excavation and analysis, the site would have continued to be another doubted case of a human-mammoth association. As it represents the addition of another site to debates where the number of sites is often presented as evidence against (e.g., Grayson Reference Grayson2001; Haynes and Stanford Reference Haynes and Stanford1984; Meltzer Reference Meltzer1986; Wroe et al. Reference Wroe, Field, Fullagar and Jermin2004) or for (Surovell and Grund Reference Surovell and Grund2012; Surovell and Waguespack Reference Surovell and Waguespack2008) Pleistocene overkill and Clovis large game specialization, La Prele serves as a reminder that there are likely other sites that are currently underinvestigated to be considered reliable enough to include in counts of terminal Pleistocene megafaunal kill/scavenge sites. The traditional three-part criteria (contemporaneity, direct association, and use) employed to evaluate Clovis kill/scavenge sites are reasonable but require a burden of proof that is not possible without significant investigation and careful excavation. Byers's (Reference Byers2002) doubt was reasonable given the limited artifact assemblage and minimal geologic context available. The results of new excavations at the La Prele Mammoth site should encourage the investigation of other proboscidean sites that are currently in limbo due to insufficient reporting or excavation.
Clarification of the number and type of proboscidean death/butchery sites can contribute to a variety of debates in Paleoindian archaeology, including overkill, Clovis subsistence, and the social and economic implications of procuring large animals. As at least the sixteenth identified extinct megafauna processing site, La Prele, and its expanded assemblage, is an important contribution to the Clovis literature. The presence of multiple artifact concentrations contemporaneously occupied during mammoth butchery is uncommon, with only one other case (or possibly two) reported (Haynes and Huckell Reference Haynes and Huckell2007; Sanchez et al. Reference Sanchez, Holliday, Gaines, Arroyo-Cabrales, Martínez-Tagüeña, Kowler, Lange, Hodgins, Mentzer and Sanchez-Morales2014). Further excavation and analysis of these activity areas is ongoing and will contribute to our understanding of the full suite of activities that took place during megafauna butchery in the terminal Pleistocene.
Acknowledgments
We thank Jack and Zach Amen for allowing access to the site. We are very grateful to James and Shirley Baker for running the backhoe, providing various site construction materials, and fixing all the things we have broken over the years. Without countless excavators including volunteers, undergraduate and graduate students, members of the Northern Arapahoe Tribe, Rich Adams, and four years of the University of Wyoming Archaeological Field School (2014–2017), this project would not have been possible. Laboratory analysis would have taken significantly longer without the help of volunteers including Paul Sanders, Sandra Zarzycka, Rebecca Mattson, and a team of additional students. Thank you goes to the University of Wyoming Archaeological Repository for allowing us access to the 1987 collection and to Sarah Allaun D'Lopez for translating the Spanish abstract. Support for this project was provided by the Frison Institute of Archaeology and Anthropology, the University of Wyoming Archaeological Field School, the Shlemon Center for Quaternary Studies, the National Geographic Society (#9896-19), the Wyoming Cultural Trust Fund, and the Quest Archaeological Research Program. We thank Thomas Jennings, Lynn Gamble, and three anonymous reviewers for their comments on an earlier draft of this manuscript.
Data Availability Statement
All data and collections associated with the La Prele Mammoth site are housed at the University of Wyoming Department of Anthropology in Laramie, Wyoming.
Supplemental Materials
For supplementary material accompanying this essay, visit https://doi.org/10.1017/aaq.2020.8.
Supplemental Text 1. Description of geologic context for the La Prele Mammoth site.
Supplemental Text 2. Lithic microwear methods, blind test results, and wear descriptions for La Prele Mammoth (LPM) tools.
Supplemental Table 1. Radiocarbon dates from excavations (Blocks A and B) and Trench 1 at the La Prele Mammoth site.
Supplemental Table 2. Optically stimulated luminescence dates from Trench 1.
Supplemental Table 3. Microwear characteristics for tools listed in Table 3.
Supplemental Figure 1. Profile of Trench 1 from 2014 including all optically stimulated luminescence (OSL) and radiocarbon dates. Note that OSL dates are presented in calendar years before present while radiocarbon dates are presented in radiocarbon years before present.
Supplemental Figure 2. Examples of lithic microwear patterns observed on the La Prele Mammoth (LPM) tool assemblage and the polar coordinate system for identifying use locations.
Supplemental Figure 3. Examples of unmodified and retouched tools from an experimental collection.