


Implications
Ruminants are important suppliers of high-quality protein because they utilize feed and by-products of little or no value for human food. Improved environmental and economic sustainability will derive from greater productivity per animal because this dilutes out fixed nutrient costs of maintenance and similar functions and reduces emission intensity of greenhouse gases (GHGs) per unit product. Applying strategies such as ration balancing and nutritional models, appropriate nutrient supplementation and using livestock that better fit the ‘market’ (e.g. Jerseys and Vechur v. Holsteins) will make ruminant production more sustainable and ruminants will retain their niche for providing valuable human food.
Introduction
Humans have depended for millennia on ruminant animals, both wild and domestic, for high-quality protein. This is of course still the case, with meat and dairy products from ruminants representing major dietary protein sources in developed countries. It is anticipated that people in emerging nations will substantially increase their intake of animal protein in the future. There has, however, been a steady trend in the last few decades toward increasing consumption of poultry meat as a replacement for beef plus pork. In the United States, pork has held at between 20 and 25 kg/person per year, but beef consumption has fallen from 43 kg in 1975 to 24 kg per year in 2015; poultry meat has displayed a mirror-image response over the same time period, going from 21 to 48 kg/year, an increase of nearly 130% over this 40-year period (Figure 1; United States Department of Agriculture Economic Research Service (USDA ERS), 2017a). Similar trends are apparent in the EU From 1995 to 2009, consumption of poultry meat increased by 25%, whereas beef consumption declined 9% (Food and Agriculture Organization of the United Nations (FAO), 2015). These changes are driven by consumer economics: for example, the relative cost of poultry meat during the last 6 months of 2016 was 4.2 USD/kg, whereas beef averaged 12.5 USD/kg during the same period (USDA ERS, 2017a). Although beef production is lower, energetic efficiency of poultry, swine and milk is similar; however, gross protein efficiency of poultry egg and meat production exceeds that of dairy and beef cattle and swine in converting dietary protein into food protein (Table 1; Council for Agricultural Science and Technology (CAST), 1999; Wilkinson, Reference Wilkinson2011). A more appropriate way to look at this conversion is to account for the proportion of feedstuffs for each production system deriving from potential human food. By this metric, beef and particularly dairy cattle are much more efficient in net production of human food protein. This is because both swine and poultry can consume dietary ingredients that can be eaten directly by humans. Of particular note are the data on Argentinian beef production: gross efficiency of converting dietary protein to meat protein is only 0.02, but conversion of human-edible protein is 6.1 (Table 1); this high value derives from the fact that beef cattle consume only pasture and non-edible by-products in the Argentinian production system. Ruminants are major recyclers of by-products from food systems throughout the World. Oil-seed meals such as soybean meal and canola meal, which are produced during extraction of cooking oil, are major protein supplements fed to ruminants. One may argue about the wisdom of producing fuel ethanol from grains, but ruminants are the principal consumers of distillers grains co-products; the same is true for fermentation by-products deriving from production of alcohol consumed by humans. By-products from human food production such as crop residues, almond hulls, vegetable wastes, ‘cookie dough’ and a wide variety of materials represent major dietary ingredients wherever ruminants are fed.

Figure 1 Per capita meat consumption in the United States (USDA ERS, 2017a).
Table 1 Animal systems: gross efficiencies of converting energy and protein into product and returns of human-edible inputs in productFootnote 1

1 Gross efficiencies estimated as outputs of human-edible energy and protein divided by total energy and protein inputs. Human-edible returns calculated as human-edible outputs divided by human-edible inputs.
2 Data summarized from CAST (1999).
3 Data summarized from Wilkinson (Reference Wilkinson2011). Values for ‘Beef’ are means of three production systems.
Value of consuming high-quality ruminant proteins
Human per capita protein consumption averages 77 g/day but the range is very wide – from 55 g/day in Sub-Saharan Africa to 103 g/day in the developed world (FAO, 2010). Even 55 g/day would be adequate if the protein had a good to excellent pattern of essential amino acids (EAA). The EAA are also referred to as indispensable amino acids – those nine protein amino acids that cannot be synthesized in the tissues but must be absorbed from the intestinal tract of all animals. Much of the food protein consumed in developing nations is of plant origin with less than optimal EAA composition. This point is illustrated by the data in Table 2, which compares protein quality of milk protein with that almond milk, a plant ‘milk’ the sales for which are growing rapidly in North America. As outlined by Professor Ertl at the Bristol meeting in January 2016, a comparison was made using the Digestible Indispensable Amino Acid Score (DIAAS) for evaluating the relative quality of food proteins (Ertl et al., Reference Ertl, Knaus and Zollitsch2016). This score relates a food’s pattern of truly digested EAA to one of three standard FAO (2013a) patterns of required EAA. Data are available on true digestibility of individual EAA in milk but not almond milk; as recommended by FAO (2013b), the overall true digestibility for almond protein was applied to all EAA. The FAO EAA pattern for children aged 6 months to 3 years was used as the standard. Aside from the fact that almond milk provides only 1 g protein per serving v. 8 g for cow’s milk, the DIAAS value of its protein is only 0.41 of the standard FAO pattern because of low lysine concentration. The DIAAS value of cow’s milk is 1.12 based on its S-amino acid (methionine plus cystine) content. Hence, the protein in cow’s milk has a relative DIAAS value=100×1.12/0.41=270% of the protein in almond milk. One must wonder at the growing popularity of almond milk.
Table 2 Comparison of the protein quality of almond milk and cow’s milk using the Digestible Indispensable Amino Acid Score (DIAAS) method (Ertl et al., Reference Ertl, Knaus and Zollitsch2016)

FAO=Food and Agriculture Organization of the United Nations.
1 Standard pattern of require indispensable amino acids, expressed in mg amino acid/g protein consumed, for lysine (Lys), S-amino acids (SAA), threonine (Thr) and tryptophan (Trp) (FAO, 2013a).
2 SAA=methionine plus cystine.
3 True digestibility of almond protein, mean of three cultivars from Ahrens et al. (Reference Ahrens, Venkatachalam, Mistry, Lapsley and Sathe2005).
4 Digestible indispensable amino acids (DIAA) (true digestibility×amino acid concentration in protein).
Plant protein quality can be improved substantially by supplementation with ruminant proteins. The second comparison in Table 2 illustrates how complementary EAA patterns of animal and plant protein can be used to improve the value of the latter for human food. Peanuts (groundnuts) are relatively high in total protein and are widely consumed in both the developing and developed world. As is shown in Table 3, peanut protein is limiting in lysine, with 0.57 of the FAO standard for young growing children. However, a blend of milk and peanut proteins (two ‘cups’ of milk plus 150 g of peanuts) yields 53 g of protein that has 0.83 of the lysine concentration of the FAO standard. Thus, foods of ruminant origin provide proteins of excellent EAA composition with complementary EAA patterns.
Table 3 Comparison of the protein quality of peanut (groundnut) protein alone and when supplemented with milk protein using the Digestible Indispensable Amino Acid Score (DIAAS) method (Ertl et al., Reference Ertl, Knaus and Zollitsch2016)
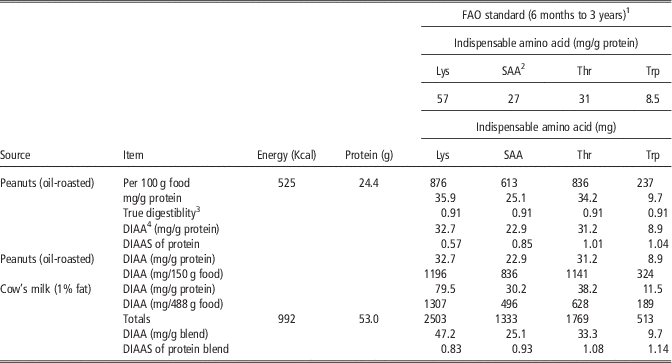
FAO=Food and Agriculture Organization of the United Nations.
1 Standard pattern of require indispensable amino acids, expressed in mg amino acid/g protein consumed, for lysine (Lys), S-amino acids (SAA), threonine (Thr) and tryptophan (Trp) (FAO, 2013a).
2 SAA=methionine plus cystine.
3 True digestibility of peanut protein from Rutherfurd et al. (Reference Rutherfurd, Fanning, Miller and Moughan2015).
4 Digestible indispensable amino acids (DIAA) (true digestibility×amino acid concentration in protein).
Improving productivity reduces environmental impact
In recent years, livestock production systems have seen considerable improvement in output of product per unit feed input. Dijkstra et al. (Reference Dijkstra, France, Ellis, Strathe, Kebreab and Bannink2013) computed that, using the UK metabolizable energy system, energetic cost per kilogram of fat-corrected milk (FCM) and protein-corrected milk decreased 19% when milk yield increased from 6000 to 10 000 kg/cow per year. Wilkinson and Garnsworthy (Reference Wilkinson and Garnsworthy2017) showed that replacing dietary grazed grass or grass silage with corn silage at similar milk yield increased N efficiency and reduced the C footprint. Similar reduction is also observed for metabolizable protein expenditures per kilogram of FCM and protein-corrected milk. These improvements result from dilution of nutrient costs for maintenance and other functions, such as those required for rearing the calf to first lactation and supporting the dry period. Dijkstra et al. (Reference Dijkstra, France, Ellis, Strathe, Kebreab and Bannink2013) also assessed human food energy and protein output per unit input of potential human food with differing number of lactations/cow. Efficiency increased substantially with increasing numbers of lactations/cow, maximizing at 3.5 lactations/cow; the US mean is 2.5 lactations/cow. Improved reproductive efficiency of lactating cows reduces the proportion of heifers that must be retained in the herd, as well as increases lactations/cow, thus improving efficiency of conversion of feed nutrients to milk (Garnsworthy, Reference Garnsworthy2004). Greater animal productivity, more lactations per cow and improved reproductive efficiency are clearly effective means for diluting out the nutrient costs of ruminant food systems.
Methane production in the rumen represents an important loss of feed energy. Moreover, methane is a major GHG and domestic ruminants contribute an estimated 25% of total anthropogenic methane production (United States Environmental Protection Agency (US EPA), 2016). It can be argued that this is not a new phenomenon, because wild ruminants were major methane contributors in the past. For example, one estimate indicates that methane production from the wild ruminants (mostly bison) in the pre-settlement contiguous United States was 86% of the current magnitude from farmed ruminants in the same area (Hristov, Reference Hristov2012). Just as happens with gross energy and protein efficiencies, methane emission per unit product is also reduced with increasing productivity. It is estimated that the intensity of methane production declined from 31 in 1924 to 14 g/kg milk in 2014 (US EPA, 2016). However, total GHG emissions (of which methane is a major part) plateau at about 2 kg CO2-equivalents/kg fat and protein-corrected milk when yield reaches 4000 kg/year of fat and protein-corrected milk, changing little at higher production (Gerber et al., Reference Gerber, Vellinga, Opio and Steinfeld2011).
Reducing methane production in the rumen has been a much sought after goal for decades in ruminant nutrition. A number of compounds, for example, the ionophores monensin and lasalosid, have been found to reduce methanogenesis in short-term studies. The improvement in energetic efficiency obtained with feeding ionophores is widely believed to occur largely by suppressing methane formation. However, Guan et al. (Reference Guan, Wittenberg, Ominski and Krause2006) found that the depression of methanogenesis obtained with feeding cattle monensin alone or monensin plus lasalosid disappeared in 7 to 8 weeks. That these effects often do not persist is related to the thermodynamic advantage to methanogenic archaea of reducing carbon dioxide and other 1-carbon units with hydrogen; this results in rapid microbial adaptation such that methane production soon returns to pre-treatment levels (Patra et al., Reference Patra, Park, Kim and Yu2017); however, there is hope. Recent reports indicate that the inhibitor 3-nitrooxypropanol reduced methane production by more than 30%, an effect that persisted for at least 12 weeks (Hristov et al., Reference Hristov, Oh, Giallongo, Frederick, Harper, Weeks, Branco, Moate, Deighton, Williams, Kindermann and Duval2015). Moreover, certain dietary lipids such as medium-chain fatty acids and coconut oil also reduce methane production (Machmüller et al., Reference Machmüller, Dohme, Soliva, Wanner and Kreuzer2001). These lipids act partly by suppressing rumen protozoa, thus also suppressing the methanogenic archaea that exist in close association with protozoa (Patra et al., Reference Patra, Park, Kim and Yu2017). One wonders whether the effects of lipids and 3-nitrooxypropanol would be additive, or even complementary, such that combining these materials might yield even greater reduction in rumen methanogenesis. Abecia et al. (Reference Abecia, Martín-García, Martínez, Newbold and Yáñez-Ruiz2013) reported that reduced methane production persisted for at least 3 months after kids were treated with bromochloromethane, another methanogen inhibitor. Yanez-Ruiz et al. (Reference Yanez-Ruiz, Abecia and Newbold2015) summarized literature suggesting that altering the rumen microbiome in early-life may reduce methane emissions in later life.
For a long time, livestock farmers believed that high-protein diets were more beneficial, increasing animal productivity. Dairy scientists have recently conducted several trials with the objective of assessing ‘requirements’ for dietary CP and found that producers were often over-feeding CP. Our own work showed that, with step-wise increases from 15.1% to 16.7% and 18.4% CP (achieved by adding solvent-extracted soybean meal to the diet at the expense of grain), milk and protein yield increased with the initial increment, but the second CP increment increased feed intake but with no effect on milk or protein yield (Broderick, Reference Broderick2003). The only result of increasing CP from 16.7% to 18.4% was increased excretion of environmentally unstable urinary N; urinary N accounted for nearly all the additional CP equivalent. These three protein levels were each tested in diets containing three different concentrations of NDF. Although lower NDF (higher net energy) diets gave rise to greater milk yield, the relative response to dietary CP was the same across all three energy levels. A number of other trials were also conducted around this same time addressing dietary CP concentration (added as soybean meal). Wattiaux and Karg (Reference Wattiaux and Karg2004) found that increasing dietary CP from 16.5% to 17.9% did not improve yield of milk or milk components when diets contained 50% of dry matter (DM) as either alfalfa or corn silage. Olmos Colmenero and Broderick (Reference Olmos Colmenero and Broderick2006) made step-wise increases of 1.5 percentage units, from 13.5% to 19.4% CP and observed quadratic responses indicating that milk and protein yields maximized at 16.7% and 17.1% CP, respectively. In all three trials, rising milk urea N and urinary urea, and declining milk N/N intake, reflected the linear decline in N utilization with increasing CP. That this information has been applied in practice is illustrated in Table 4. Keuning et al. (Reference Keuning, Gunderson and Shaver1999) surveyed the six Wisconsin dairy producers with the greatest rolling herd averages (milk/305-day lactation) and found that their lactating cows diets averaged 19.4% CP; indeed, one producer was feeding 21.5% dietary CP. A more recent survey (Shaver, Reference Shaver2010) showed an interesting trend: lactating cows in the five Wisconsin herds with the greatest milk yield/lactation were being fed diets averaging 16.9% CP, with the lowest herd receiving 16.3% dietary CP. Per lactation rolling herd averages actually increased 1350 kg milk, 65 kg fat and 45 kg true protein (Table 4). Indeed, Shaver (Reference Shaver2010) reported that, in the period from 2004 to 2010, CP content of dairy rations declined by a mean 1.1 percentage units while milk and protein secretion rose, respectively, 1700 and 51 kg/lactation. Thus, not over-feeding dietary CP is an excellent means of improving N efficiency and reducing excretion of urinary N, the most environmentally labile form of manure N.
Table 4 Dietary CP contents and milk, fat and protein yields determined from surveying the Wisconsin dairy herds with highest rolling herd averages

DM=Dry matter.
1 Rolling herd averages reported to farmers by AgSource Dairy Herd Improvement Cooperative.
2 Survey data on six herds reported by Keuning et al. (Reference Keuning, Gunderson and Shaver1999).
3 Yield and mean milk concentration of milk total protein.
4 Mean milk fat concentration.
5 Range of dietary CP concentrations.
6 Survey data on five herds reported by Shaver (Reference Shaver2010).
7 Yield and mean milk content of milk true protein.
Kalscheur et al. (Reference Kalscheur, Vandersall, Erdman, Kohn and Russek-Cohen1999) found that reducing dietary CP from 17.4% to a mean 15.2% during weeks 4 to 14 of lactation reduced yields of milk and milk components. However, decreases from 15.3% to a mean 13.4% CP (weeks 19 to 29 of lactation) and from 14.2% to a mean 12.6% CP (weeks 24 to 34 of lactation) did not significantly reduce production of milk and milk components. The principal effect observed in the latter two phases of lactation with lower CP were improvements of 2 to 3 percentage units in apparent N efficiency. Wu and Satter (Reference Wu and Satter2000) divided the lactation curve into early lactation (weeks 1 to 16 after calving) and later lactation (weeks 17 to 44 after calving), and applied 4 different protein supplementation regimes over the whole lactation (Table 5). The dietary CP regime supporting optimum yield of 3.5% FCM with minimal manure N excretion over the whole lactation involved feeding 17.4% CP for the first 16-weeks after calving, followed by 16.0% CP for the remaining 28 weeks. Increasing dietary CP to as high as 19.3% during the first phase, or to 17.9% CP during the second phase, did not improve FCM yield and only increased manure N output. Reducing dietary CP intake in lactating cows substantially reduced volatile N losses from the stored manure (Külling et al., Reference Külling, Menzi, Kröber, Nefte, Sutter, Lischer and Kreuzer2001). Partly as a convenience, dairy farmers often feed only a single ration to their lactating cows; because these diets are mainly designed for the animals with highest nutrient requirements, cows will be overfed CP for most of the lactation. In addition to reducing N excretion, group feeding cows to better match their nutrient requirements lowers feed costs and improves economic sustainability of dairy production. In their meta-analysis of North American and Northern European data on effects of varying dietary CP and energy content and intake on milk production and N utilization, Huhtanen and Hristov (Reference Huhtanen and Hristov2009) concluded that limiting CP intake was the most effective way to improve N efficiency. These workers also found that energy and CP intakes yielded the most reliable predictions of milk protein yield and N efficiency; adding rumen CP degradability (as predicted by National Research Council (NRC), 2001) reduced model precision. Sinclair et al. (Reference Sinclair, Garnsworthy, Mann and Sinclair2014) recently reviewed effects of dietary CP and energy intake in lactating cows and concluded that protein yield could be maintained at high N efficiencies.
Table 5 Effect on yield of 3.5% fat-corrected milk (FCM) and excretion of manure N of feeding dairy cows fed four different CP regimes during the first 16 weeks and last 28 weeks of 44-week lactations (Wu and Satter, Reference Wu and Satter2000)

a,b,cMeans in columns without common superscripts are different (P<0.05).
Protein quality is important to productive ruminants
Until the mid-20th century, students of ruminant nutrition were taught that dietary protein quality was of no importance because microbial protein synthesis in the rumen met the animal’s requirements for EAA. Indeed, there was clear evidence that rumen microbes utilized non-protein N sources such as urea to provide metabolizable EAA to the ruminant (e.g. Virtanen, Reference Virtanen1966). However, a paradigm shift started in the early 1960s with nutritionists recognizing the importance of EAA composition of ruminant diets. Large responses in wool growth were observed with abomasally infused protein and gram quantities of methionine and cystine (e.g. Reis and Schinckel, Reference Reis and Schinckel1964). Substantial responses were later obtained with abomasal infusions of casein into lactating cows (e.g. Broderick et al., Reference Broderick, Kowalczyk and Satter1970), and abomasal infusions of casein and EAA mixtures into growing lambs (e.g. Schelling and Hatfield, Reference Schelling and Hatfield1968) and beef steers (Titgemeyer and Merchen, Reference Titgemeyer and Merchen1990). Since that era, a large body of literature has developed indicating that, particularly in very productive ruminants, microbial protein synthesis is not adequate to meet all of the animal’s needs; these ruminants also required additional amounts of the EAA provided by dietary protein escaping the rumen (rumen-undegraded protein (RUP)). Supplementing RUP represents one way to reduce total CP intake, reducing the environmental footprint, without losing production. Typical results from dairy nutrition studies are presented in Table 6 (Broderick et al., Reference Broderick, Ricker and Driver1990). Replacing conventional solvent-extracted soybean meal with expeller soybean meal, which is subjected to substantial heating during processing and has greater RUP content, increased production and reduced milk urea concentrations, an indicator of urinary N excretion (Nousiainen et al., Reference Nousiainen, Shingfield and Huhtanen2004). Supplementation of about 400 g CP from expeller soybean meal actually provided more RUP, and supported numerically superior production, than supplementation with about 700 g CP from solvent-extracted soybean meal (Table 6). Brito and Broderick (Reference Brito and Broderick2007) observed substantially greater milk and milk protein secretion when supplementing 16.5% CP diets with true proteins (soybean meal, cottonseed meal and canola meal) v. urea; moreover, yield of milk components was greatest among the true proteins when diets contained canola meal, least with cottonseed meal and intermediate with soybean meal. These differential responses occurred, despite the fact that the cottonseed meal diet supplied the greatest amount of RUP (Brito et al., Reference Brito, Broderick and Reynal2007). The results were explained by the EAA composition of the RUP: although soybean meal is low in methionine, cottonseed meal is very low in lysine, and canola meal has a more balanced content of both methionine and lysine (NRC, 2001). Both a greater proportion RUP in its CP (Broderick et al., Reference Broderick, Columbini, Costa, Karsli and Faciola2016), plus greater methionine in that RUP (NRC, 2001), explain the consistently greater milk protein yields observed when canola meal replaces soybean meal in the diet of lactating dairy cows (Huhtanen et al., Reference Huhtanen, Hetta and Swensson2011; Martineau et al., Reference Martineau, Ouellet and Lapierre2013).
Table 6 Response of lactating cows to supplementation of rumen-undegraded protein (RUP) from solvent-extracted soybean meal (SSBM) or expeller soybean meal (ESBM) fed in supplement of alfalfa silage-based diets (Broderick et al., Reference Broderick, Ricker and Driver1990)Footnote 1

DM=Dry matter; MUN=milk urea N.
1 Means from trials 1 and 2 (Broderick et al., Reference Broderick, Ricker and Driver1990).
Schwab et al. (Reference Schwab, Satter and Clay1976) found that milk protein secretion increased 100 g/day with abomasal infusion of a mixture of nine EAA plus arginine in dairy cows fed a low CP basal diet; milk protein response to abomasal infusion of lysine plus methionine alone was only 40 g/day. Consistent observation of methionine and lysine as limiting EAA in abomasal infusion studies led eventually to development of rumen-protected methionine (RPM) and rumen-protected lysine (RPL). There have been consistent responses to RPM in dairy cattle on practical diets; the meta-analyses of Patton (Reference Patton2010) and Zanton et al. (Reference Zanton, Bowman, Vázquez-Añón and Rode2014) summarize most of this research. Somewhat less consistent responses to RPL have been obtained in dairy cattle (Vyas and Erdman, Reference Vyas and Erdman2009). Rumen microbial protein has adequate levels of both lysine and methionine (NRC, 2001). However, RUP from soybean meal, the most common protein supplement, is high in lysine but relatively low in methionine (NRC, 2001), which probably explains these differential effects. Substantial milk protein responses to RPL are obtained when it supplements diets in which most or all of the RUP derives from grain sources such as corn distillers grains and corn gluten meal (e.g. Lobos et al., Reference Lobos, Broderick and Wattiaux2014). Rumen microbial protein is low in histidine relative to the EAA pattern of milk; Finnish workers detected histidine as first-limiting EAA when lactation diets were based on grass silage that contributed little RUP (Huhtanen et al., Reference Huhtanen, Vanhatalo and Varvikko2002). Lee et al. (Reference Lee, Hristov, Cassidy, Heyler, Lapierre, Varga, de Veth, Patton and Parys2012) fed RPM, RPL plus their own preparation of rumen-protected histidine in low protein (13.5% CP) diets and compared production to that on a 15.7% CP control diet (Table 7). These workers observed that milk and protein yield was only restored when all three rumen-protected EAA were supplemented. Addition of RPM and RPL increased blood methionine and lysine concentrations, but blood histidine remained very low until the rumen-protected histidine preparation was fed. Our excitement at the possibility of maintaining production while lowering urinary N excretion 32% on a 13.5% dietary CP is somewhat tempered by the apparent depression in NDF digestion in the rumen. Dietary CP not only supplies EAA via RUP, it also provides degraded CP that stimulates microbial fiber digestion in the rumen. A lack of degraded CP appears to have depressed fiber digestion.
Table 7 Effect of supplementing a low CP diet with rumen-protected methionine, lysine and histidine on production and metabolism of lactating dairy cows (Lee et al., Reference Lee, Hristov, Cassidy, Heyler, Lapierre, Varga, de Veth, Patton and Parys2012)

DM=dry matter; NRC=National Research Council.
a,b,cMeans in rows without common superscripts are different (P<0.05).
1 Supplemented with 18 and 24 g/day of rumen-protected methionine and lysine (ML).
2 Supplemented with 18, 24 and 12 g/day of rumen-protected methionine, lysine and histidine (MLH).
3 Probability of a significant effect of diet.
4 Metabolizable protein supply, computed using the NRC (2001) model.
5 PBV=net supply of total N to the rumen, estimated using the NorFor (2011) model. Negative values indicate inadequate supply of rumen-degraded protein.
Research applicable to developing countries
Much of the large body of literature on improving the environmental and economic sustainability of ruminant production derives from research conducted in developed countries, often under conditions of very high productivity. The amino acid composition of milk and tissue proteins is similar across mammalian species and ruminants depend on microbial protein to meet more than half of their requirements; thus, ruminants may be expected to respond similarly to improved supply of metabolizable protein and metabolizable amino acid at lower levels of production. This is illustrated by research results obtained in El Salvador by Corea et al. (Reference Corea, Aguilar, Alas, Alas, Flores and Broderick2017). These workers tested feeding cowpea hay (Vigna sinensis), a legume that is relatively productive when grown under tropical conditions, as a replacement for one-fourth of the commonly fed sorghum silage in the diet of crossbred dairy cows. As cowpea hay is substantially higher in CP than sorghum, less soybean meal was required in the diet, lowering the ration cost. Replacing sorghum silage with cowpea hay resulted in increased milk/DM intake and milk N/N intake, lowered milk urea and urinary and fecal N excretion, and improved income over feed cost (Table 8). Moreover, the authors reported that increasing dietary CP by 1.5 percentage units only increased N excretion and feed cost without improving production.
Table 8 Effect of dietary CP concentration and partial replacement of sorghum silage with cowpea (Vigna sinensis) hay on productivity and profitability of lactating dairy cows in El Salvador (Corea et al., Reference Corea, Aguilar, Alas, Alas, Flores and Broderick2017)
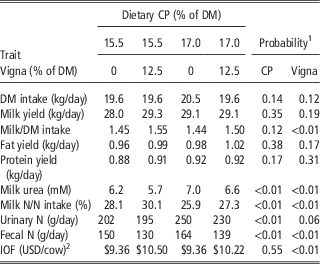
DM=dry matter.
1 Probability of orthogonal contrasts: CP=dietary CP concentration; Vigna=cowpea (Vigna sinensis) hay.
2 Income over feed cost in US dollars.
Small amounts of grain supplementation often result in substantial improvements in productivity of ruminants on pasture or other high-forage regimes. One example is the response reported by Wales et al. (Reference Wales, Kolver, Egan and Roche2009) to supplementing about 2.5 and 5.0 kg DM/day of a carbohydrate mix to lactating cows grazing ryegrass pasture (Figure 2). Both levels of carbohydrate supplementation reduced pasture forage consumption but the first increment improved milk yield by 2.2 kg/day and N efficiency by about 3 percentage units. The second increment also improved N efficiency but had no further effect on milk yield. Milk protein yield increased by about 100 g/day with the first increment of carbohydrate supplementation, and milk protein content increased about 0.1 percentage unit with each increment of supplementation. This often observed response occurs because fermentable energy stimulates microbial protein formation in the rumen and metabolizable protein supply to the animal – the carbohydrate actually serves as a protein supplement. Earlier reports of this type of response in dairy cows fed alfalfa silage in confinement are those of Dhiman and Satter (Reference Dhiman and Satter1993), where corn silage supplementation stimulated milk and protein yield, and Ekinci and Broderick (Reference Ekinci and Broderick1997), where grinding of high moisture corn reduced rumen ammonia and urinary N excretion and stimulated milk and protein yield. Sampath et al. (Reference Sampath, Chandrasekharaiah and Praveen2008) obtained very similar responses when replacing about 1.5 kg/day of wheat bran with about 1.5 kg/day of maize (corn) grain in native Indian cattle fed largely on locally grown grass hay. This substitution did not increase feed costs, increased yield of milk and milk components and increased income over feed cost by 13 rupees/cow/day. All of these approaches for increasing diet fermentability increased both economic and environmental sustainability of dairy production.
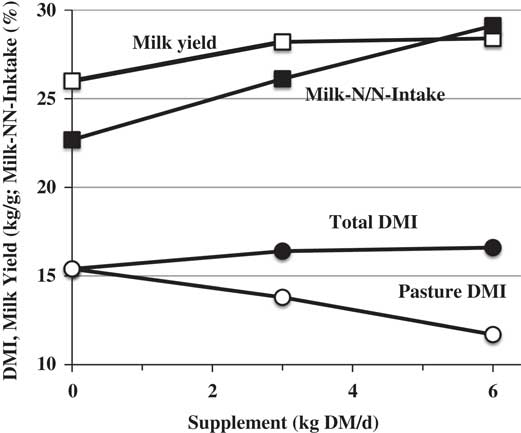
Figure 2 Effect on dry matter (DM) intake (DMI), milk yield and apparent N efficiency (milk N/N intake) of supplementing 2.5 and 5.0 kg DM/day from a carbohydrate mix to cows grazing perennial ryegrass in New Zealand (Wales et al., Reference Wales, Kolver, Egan and Roche2009). Supplement contained (DM basis) 65% barley, 30% steam-flaked maize, 5% molasses and 10% CP.
Impressive results on reducing GHG emission intensity were obtained by Garg et al. (Reference Garg, Sherasia, Phondba and Makkar2016) when instituting an online ration balancing system for smallholder operations in four dairy regions of India (Table 9). Versus conventional nutrition and husbandry, life-cycle analysis was used to assess the effects of balancing rations from a large array of locally available feeds on GHG emissions from about 164 000 each of dairy cows and buffalo. Based on these computations, ration balancing substantially improved yields of milk, fat and protein-corrected milk and reduced emission intensity (GHG/milk yield) by about one-third in both species. Note that methane and nitrous oxide production during lactation, and lifetime GHG equivalents from enteric methane, actually increased under the ration balancing regimes, but greater milk yields and much lower GHG contributions from unproductive animals improved emission intensity. Similar improvements in milk yields and in income with ration balancing were observed in smaller studies with indigenous dairy cows in Bangladesh (Kamal et al., Reference Kamal, Iqbal and Khaleduzzaman2009) and buffalo in a second Indian trial (Kumar et al., Reference Kumar, Nayak, Baghel and Khare2014). Clearly, strategic implementation of improved nutritional and management practices reduces the environmental footprint and improves the economic sustainability of ruminant production in smallholder dairy enterprises in developing countries.
Table 9 Effect of applying ration balancing to diets fed to 164 000 cows and 164 000 buffalo in four dairy production regions of India on estimated greenhouse gas (GHG) emissions and GHG emission intensity (Garg et al., Reference Garg, Sherasia, Phondba and Makkar2016)

MT=metric tons; CO2-eq=GHG expressed in CO2-equivalents; FPCM=fat- and protein-corrected milk.
*Probability of significant effect of ration balancing programs (P<0.05).
1 Before and after application ration balancing (RB) programs available to farmers online.
2 Unproductive animals are those that can no longer produce milk.
Using animals better adapted to local conditions
Milk production in the Holstein Friesian breed has increased to truly impressive levels, with average yield per animal exceeding 15 000 kg/lactation in elite herds (e.g. Shaver, Reference Shaver2010). Production per cow in the overall US dairy herd, which is more than 90% Holstein, exceeded 10 000 kg/year in 2014 (United States Department of Agriculture National Agricultural Statistics Service (USDA NASS), 2016). Although this level of production puts considerable stress on the animal, management has evolved such that these cows can be maintained in the resource rich environments available in North America and Europe. However, the market for dairy products has been shifting; much more milk equivalent is consumed as products such as cheese and much less as fluid milk. In the United States, per capita consumption of fluid milk has declined from 115 kg/year in 1975 to 90 kg/year in 2010 (USDA ERS, 2017b). In Wisconsin, about 90% of the 12.6 million metric tons of milk produced each year goes to cheese production and other non-fluid uses (USDA NASS, 2016). The 725 kg of lactose secreted in 15 000 kg of milk is a relatively low value product that puts considerable metabolic stress on the cow. The value of milk as human food is mainly related to its protein concentration even when consumed on the farm as fluid milk. The higher protein and fat concentrations in Jersey milk fit more closely with a cheese market. Prendiville et al. (Reference Prendiville, Pierce and Buckley2009) compared whole-lactation data from Holstein, Jerseys and Holstein×Jersey crosses under Irish grazing management with limited grain supplementation. As expected, Holsteins secreted more milk than purebred Jerseys and Holstein-Jersey crosses. However, yield of fat plus protein per unit total DM intake of Jerseys and Holstein-Jersey crosses was 11% greater than for Holsteins; protein yields for Holstein, Jerseys and Holstein×Jersey crosses were, respectively, 0.39, 0.37 and 0.41 kg/day in this study. Capper and Cady (Reference Capper and Cady2012) compared the environmental impact of the milk production from Jerseys v. Holsteins required to produce 500 000 metric tons of cheese. The authors computed that 4.94 billion kg of Holstein milk v. 3.99 billion kg of Jersey milk would be needed. Note that, because of lower protein : fat ratio, 500 000 metric tons of cheese made from Jersey milk would contain 148 000 tons of protein and 192 000 tons of fat v. 153 000 tons of protein and 188 000 tons of fat if the cheese were made from Holstein milk. Lower yield per cow meant a greater Jersey population would be necessary; however, factors such as lower maintenance due to smaller body size reduced overall requirements for nutrients, land area and water for the Jersey breed. The C footprint for producing the milk for 500 000 metric tons of cheese was reduced by 1.66 metric tons of CO2-equivalents for Jerseys v. Holsteins.
There is renewed interest in smaller, native breeds such as Vechur cattle (Eisler et al., Reference Eisler, Lee, Tarlton, Martin, Beddington, Dungait, Greathead, Liu, Mathew, Miller, Misselbrook, Murray, Vinod, Van Saun and Winter2014). Such cattle produce much less milk and meat per animal but are better-adapted to the tropical and sub-tropical environments in the developing world. Many of the responses described above with improved nutrition and management were obtained using native breeds (Garg et al., Reference Garg, Sherasia, Phondba and Makkar2016) or crosses between European breeds and native cattle (Corea et al., Reference Corea, Aguilar, Alas, Alas, Flores and Broderick2017). Another relatively new direction is the identification of, and selection for, dairy cattle with lower than average residual feed intakes (RFI); RFI values reflect the feed that goes for other than productive functions, such as maintenance and GHG emissions. Two recent reviews dealing with RFI and factors influencing RFI have been recently published (Waghorn and Hegarty, Reference Waghorn and Hegarty2011; Vandehaar et al., Reference VandeHaar, Armentano, Weigel, Spurlock, Tempelman and Veerkamp2016). Genetic selection for lower RFI will likely be a tool used for future improvements in sustainability of ruminant livestock production.
Precision feeding of ruminant livestock
Precision livestock farming is a relatively new approach to the very old idea of matching as precisely as possible production with procurement of the resources required for a livestock farming operation (Halachmi and Guarino, Reference Halachm and Guarino2016). Interest is such that the journal Animal (no. 9, vol. 10, 2016) published a recent issue devoted solely to precision livestock farming. Precise feeding of a farm’s livestock is an important component of precision livestock farming. Moreover, nutritional models are now widely applied to improve nutrient efficiency as well as profitability of dairy production (Feed into Milk, 2004; NorFor, 2011; INRA, Sauvant et al., Reference Sauvant, Cantalapiedra-Hijar, Delaby, Daniel, Faverdin and Nozière2015; NRC, White et al., Reference White, Roman-Garcia, Firkins, VandeHaar, Armentano, Weiss, McGill, Garnett and Hanigan2017a and Reference White, Roman-Garcia, Firkins, Kononoff, VandeHaar, Tran, McGill, Garnett and Hanigan2017b). Close attention to individual animals is, of course, possible on smallholder operations; applying the newly available ration balancing tools has led to the impressive improvements in GHG emission intensity described earlier. Tools are also available in developed countries for much greater precision of nutritional management. Robotic milking systems, plus automated measurement of rumen pH, body core temperature and perhaps even feed intake, allow for collection of individual nutritional data. Heretofore, automated measurement of milk urea has not been available to farmers. The strong correlation of milk urea and N utilization efficiency would make timely milk urea data invaluable to dairy producers. For example, milking to milking changes in feed-group means for milk urea concentrations could be used to detect errors in ration formulation that require immediate correction, and would in this way improve nutrient efficiency.
Improved crops for livestock production
Certain plants contain compounds or enzyme systems that alter nutrient utilization, often improving productive efficiency and reducing environmental impact of ruminant production. In many areas of the world, forages are harvested as silages rather than hay because of greater speed of harvest and reduced risk of weather damage. However, during ensiling, usually more than half of the forage protein is broken down to amino acids and small peptides by enzymes released from cell rupture in the foliage. Red clover (Trifolium pratense) has a polyphenol oxidase (PPO) enzyme system that forms o-quinones from endogenous plant o-diphenols that react with foliage proteins to reduce their breakdown both in the silo (Lee et al., Reference Lee, Winters, Scollan, Dewhurst, Theodorou and Minchin2004; Fijałkowska et al., Reference Fijałkowska, Pysera, Lipiński and Strusińska2015) and the rumen (Broderick and Albrecht, Reference Broderick and Albrecht1997). As a result, rumen degradation of red clover protein is lower than is typical for most silages. For example, milk N/N intake was 2 to 4 percentage units higher in lactating cows fed red clover silage v. alfalfa silage in two trials (Broderick et al., Reference Broderick, Walgenbach and Maignan2001). Although DM intake was lower on red clover, milk yield/DM intake also was greater than on alfalfa silage. Marita et al. (Reference Marita, Hatfield, Brink and Mertens2012) recently reported results from feeding silage mixtures produced from co-cultures of two grass species, one of which was high in PPO (orchard grass or cocksfoot, Dactylis glomerata) and the other of which (tall fescue, Festuca arundinacea) was high in chlorogenic acid, a substrate for PPO. Versus feeding silages made from the individual grass species, feeding silages made from the grass co-cultures more than doubled mean N retention in growing wether lambs.
It is well known that presence of condensed tannins in certain legume forages such as birdsfoot trefoil (Lotus corniculatus), sainfoin (Onobrychis viciifolia) and sulla (Hedysarum coronarium) result in reduced breakdown of their proteins in the silo (Dewhurst et al., Reference Dewhurst, Delaby, Moloney, Boland and Lewis2009) and reduced degradation in the rumen (Broderick and Albrecht, Reference Broderick and Albrecht1997). Waghorn (Reference Waghorn2008) provided evidence that birdsfoot trefoil produces a type of condensed tannins in its foliage that is particularly effective for improving ruminant performance. We observed greater milk yield and N efficiency, and reduced milk urea and urinary N excretion, when replacing alfalfa silage with birdsfoot trefoil silage in three lactation studies (Hymes-Fecht et al., Reference Hymes-Fecht, Broderick, Muck and Grabber2013; Broderick et al., Reference Broderick, Grabber, Muck and Hymes-Fecht2017). Misselbrook et al. (Reference Misselbrook, Powell, Broderick and Grabber2005) observed reduced losses of volatile N from the manure from the cows fed birdsfoot trefoil in the first of these trials. Similar effects have been observed with the condensed tannins in the foliage of leguminous trees grown in tropical agroforestry. Muinga et al. (Reference Muinga, Thorpe and Topps1992) observed greater milk yield when supplementing crossbred cows with Leucaena leucocephala forage under tropical conditions. Although L. leucocephala has been widely adopted, this species is susceptible to attack by the insect Lucaena psyllid; Huang et al. (Reference Huang, Liang, Tan, Yahya, Khamseekhiew and Ho2010) reported that adding tannins extracted from a Leucaena hydrid resistant to this insect was more effective than L. leucocephala tannins in reducing both rumen protein degradation and methanogenesis. Archiméde et al. (Reference Archimède, Rira, Barde, Labirin, Marie-Magdeleine, Calif, Periacarpin, Fleury, Rochette and Morgavi2016) showed that replacing 44% of dietary DM from Dichanthium grass with one of three forage species containing condensed tannins increased dry matter intake (DMI) and depressed methane formation per unit DMI, without reducing total tract DM digestibility; the most effective forage species were L. leucocephala and Manihot esculenta, which contained the highest tannin concentrations. Lüscher et al. (Reference Lüscher, Mueller-Harvey, Soussana, Rees and Peyraud2014) have recently reviewed many of the positive aspects, including reduced GHG footprint, of feeding forages containing condensed tannins to ruminants.
Production per unit area and persistency of red clover, birdsfoot trefoil and similar legume forages lag behind that of alfalfa. However, the agronomic characteristics of these forages might well be comparable with alfalfa had they received the same amount of plant breeding research. There have been recent, successful efforts at inserting the PPO enzyme system into alfalfa (Sullivan et al., Reference Sullivan, Hatfield and Samac2008), although efforts continue to enable that plant to produce an endogenous source of o-diphenol substrate (Sullivan and Zeller, Reference Sullivan and Zeller2013). Genetic engineering is underway to extend condensed tannin expression from the seed coat to the herbage in alfalfa (Li et al., Reference Li, Dong, Ge, He, Verdier, Li and Zhao2016). Although approaches of this type maybe unacceptable in other parts of the world, the Committee on Genetically Engineered Crops of the National Academy of Sciences of the United States (Gould et al., 2016) concluded that:
… research that has been conducted in studies with animals and on chemical composition of GE [genetically engineered] foods reveals no differences that would implicate a higher risk to human health from eating GE foods than from eating their non-GE counterparts. Long-term epidemiological studies have not directly addressed GE food consumption, but available time-series epidemiological data do not show any disease or chronic conditions in populations that correlate with consumption of GE foods. The committee could not find persuasive evidence of adverse health effects directly attributable to consumption of GE foods.
Summary and conclusions
Ruminant livestock have the ability to produce high-quality human food, particularly high-quality protein, from feedstuffs of little or no value for human food and, thus, do not compete directly with the human population. Strategies that increase productivity per animal are effective for improving the sustainability of ruminant production by diluting out maintenance and other nutrient costs. Recent research indicates there is widespread over-feeding of protein to dairy cattle; milk and component yields can be maintained, and sometimes even increased, with reduced protein intake. Feeding proteins resistant to rumen microbial degradation and, in some cases rumen-protected EAA, elevate metabolizable protein and EAA supplies, further increasing performance and reducing environmental footprint. Evidence of this type has stimulated interest in precision feeding: using nutritional models to more accurately meet animal requirements. Ration balancing or feeding small supplements of readily fermented carbohydrate reduces urinary excretion of environmentally labile urea N. Modest grain supplements to grazing ruminants and those fed hay-crop silages improve N efficiency by stimulating microbial protein formation in the rumen. These strategies, plus newly developed methane inhibitors, indicate that significant reduction of enteric GHG is possible. The value of milk is directly related to its nutrient content whether milk is marketed or consumed on the farm as fluid milk; hence, nutritional strategies should maximize component yield rather than milk volume. Thus, moving away from Holsteins toward smaller breeds such as Jerseys, Holstein-Jersey crosses or locally adapted breeds (e.g. Vechur) would improve environmental and economic efficiency. Forages containing condensed tannins or PPOs have reduced rumen protein degradation; ruminants capture this protein more efficiently for meat and milk. Although these forages generally have low yields and persistence, genetic modification would allow insertion of these traits into more widely cultivated forages such as alfalfa and forage grasses. Ruminants will retain their niches because of their ability to produce valuable human food on low value feedstuffs. Emerging strategies of this type will allow improved productive efficiency of ruminants in both developing and developed countries.
Acknowledgement
This paper is an invited contribution following the First Global Farm Platform conference (12–15 January 2016, Bristol, UK). The Global Farm Platform is an international initiative linking research farms around the globe to develop solutions for sustainable ruminant livestock production (www.globalfarmplatform.org).











