


The Ca-sensing receptor (CaSR) is a class C G-protein-coupled receptor that was originally identified as the molecular ion sensor for free ionised extracellular Ca (
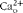 $$Ca _{o}^{2 + } $$
) homeostasis(
Reference Brown, Gamba and Riccardi
1
). Although CaSR's role in divalent cation metabolism has been well defined (reviewed by Brown & MacLeod(
Reference Brown and MacLeod
2
) and Hofer & Brown(
Reference Hofer and Brown
3
)), the CaSR is expressed in a number of tissues and cell types not typically associated with
$$Ca _{o}^{2 + } $$
homeostasis. Over the past few years, much work has been undertaken to elucidate the functional significance of CaSR expression in a wide number of other tissues including the brain and central nervous system(
Reference Bandyopadhyay, Tfelt-Hansen and Chattopadhyay
4
), the vasculature(
Reference Weston, Geraghty and Egner
5
) and the gastrointestinal (GI) tract.
$$Ca _{o}^{2 + } $$
) homeostasis(
Reference Brown, Gamba and Riccardi
1
). Although CaSR's role in divalent cation metabolism has been well defined (reviewed by Brown & MacLeod(
Reference Brown and MacLeod
2
) and Hofer & Brown(
Reference Hofer and Brown
3
)), the CaSR is expressed in a number of tissues and cell types not typically associated with
$$Ca _{o}^{2 + } $$
homeostasis. Over the past few years, much work has been undertaken to elucidate the functional significance of CaSR expression in a wide number of other tissues including the brain and central nervous system(
Reference Bandyopadhyay, Tfelt-Hansen and Chattopadhyay
4
), the vasculature(
Reference Weston, Geraghty and Egner
5
) and the gastrointestinal (GI) tract.
Although the main physiological agonist of the CaSR is Ca2+, this receptor can be activated by a diverse array of other multivalent cations including alkaline metals (Mg2+ and Sr2+), polycations (spermine and spermidine)(
Reference Quinn, Ye and Diaz
6
), aminoglycoside antibiotics (neomycin and gentamicin)(
Reference Brown, Butters and Katz
7
–
Reference Ward, McLarnon and Riccardi
9
) and cationic polypeptides (poly-l-arginine)(
Reference Brown, Katz and Butters
10
) (reviewed by Brown & MacLeod(
Reference Brown and MacLeod
2
)). Changes in ionic strength(
Reference Quinn, Kifor and Trivedi
11
) and pH(
Reference Quinn, Bai and Brown
12
) also affect CaSR's activity, as low ionic strength and high pH enhance CaSR's sensitivity to
$$Ca _{o}^{2 + } $$
.
Furthermore, as a class C G-protein-coupled receptor, the CaSR belongs to a family of extracellular amino acid sensors including the metabotropic glutamate receptors. The CaSR, along with the heterodimeric taste receptors (T1R1 and T1R3), and the goldfish 5.24 receptor and its mammalian orthologue GPRC6A form a distinct subgroup of broad-spectrum amino acid-sensing receptors, which have distinct yet overlapping sensitivities to different amino acids (Fig. 1).

Fig. 1 Amino acid profiles of the calcium-sensing receptor (CaSR), goldfish 5.24/ GPRC6A receptors, metabotropic glutamate receptor 5 (mGluR) and type 1 taste receptor (T1R)1/T1R3 heterodimer. Effective amino acid agonists of the CaSR, goldfish 5.24/GPRC6a receptors, mGluR and T1R1/T1R3 heterodimer are shown. Data for T1R1/T1R3 efficacy were obtained from the percentage of responding cells in the HEK-293 cell population expressing the mouse T1R1/T1R3 heterodimer in the absence of 2·5 mm-inosine monophosphate and normalised to 50 mm-l-Cys response(
Reference Nelson, Chandrashekar and Hoon
53
). For the CaSR, amino acid efficacy was determined from the percentage of maximum reduction in half maximal effective concentration (EC50) for extracellular
$$Ca _{o}^{2 + } $$
based on the method of Conigrave et al.
(
Reference Conigrave, Quinn and Brown
13
) and normalised to the 10 mm-l-His response. Normalised data were obtained from Conigrave & Hampson(
Reference Conigrave and Hampson
14
). We considered an effective amino acid agonist for the T1R1/T1R3 and CaSR to be one with ≥ 50 % of their respective normalised responses. GPRC6A data are based on data obtained from mouse GPRC6A in the presence of 1 mm-Ca2+ and 1 mm-Mg2(
Reference Christiansen, Hansen and Wellendorph
54
,
Reference Wellendorph, Johansen and Brauner-Osborne
55
). Darker colours are used to indicate the top two to four amino acid activators for each receptor (where applicable) based on published results(
Reference Conigrave, Quinn and Brown
13
,
Reference Conigrave and Hampson
14
,
Reference Nelson, Chandrashekar and Hoon
53
–
Reference Wellendorph, Johansen and Brauner-Osborne
55
). The receptor response to each amino acid is as follows: outer circle/red – CaSR; 2nd circle/yellow – T1R1/T1R3 heterodimer; 3rd circle/green – GPRC6A; and inner circle/blue – mGluR. (A colour version of this figure can be found online at http://www.journals.cambridge.org/bjn)
The CaSR is allosterically activated by l-amino acids, being able to respond to aromatic, aliphatic and polar amino acids, but not to branched or positively charged amino acids( Reference Conigrave, Quinn and Brown 13 ). In contrast, taste receptors can be activated by aliphatic, polar, branched-chain and, to a lesser extent, charged amino acids, but not by aromatic amino acids. Lastly, the goldfish 5.24/GPRC6A receptors respond to basic, aliphatic and polar amino acids( Reference Conigrave and Hampson 14 ). The CaSR has also been shown to respond to small peptides, including glutathione and other γ-glutamyl peptides( Reference Wang, Yao and Kuang 15 , Reference Broadhead, Mun and Avlani 16 ).
This variety in ligands enables the CaSR to act physiologically as a multi-modal sensor for several key nutrients throughout the body, including the GI tract. Within the GI tract, the CaSR is widely expressed in a number of specialised cells including the oesophagus, stomach, small intestine and colon( Reference Chattopadhyay, Cheng and Rogers 17 – Reference Conigrave and Brown 19 ) and has roles in gastrin secretion, colonic fluid transport and intestinal epithelial cell growth, all of which have been reviewed in depth previously (see Conigrave & Brown( Reference Conigrave and Brown 19 ) and Hebert( Reference Hebert, Cheng and Geibel 20 )) and are listed in Table 1. In this review article, we examine the emerging physiological functions of the CaSR in sensing dietary nutrients in two separate roles: (1) as a taste receptor for both protein and oral Ca2+ and (2) as an amino acid sensor for the release of dietary hormones within the intestine.
Table 1 Known functions of the calcium-sensing receptor in the gastrointestinal tract

CCK, cholecystokinin; GIP, gluco-indulinotropic peptide; GLP-1, glucagon-like peptide 1; PYY, peptide tyrosine tyrosine.
Calcium-sensing receptor as a taste receptor
There is now emerging evidence suggesting that the CaSR may play a role in the regulation of appetite for nutrients by modulating taste perception. The first demonstration of the possible involvement of the CaSR in taste perception was given in bullfrogs, where a positive allosteric modulator of the CaSR, the ‘calcimimetic’ NPS R-467, stimulated taste cells with accompanying neuronal responses( Reference Okada, Imendra and Miyazaki 21 ). Expression of the CaSR in rat and mouse taste cells, namely in the circumvallate, foliate and, to a lesser extent, the fungiform papillae, has recently been reported( Reference San Gabriel, Uneyama and Maekawa 22 , Reference Bystrova, Romanov and Rogachevskaja 23 ).
Taste buds are generally composed of approximately 50–100 elongated taste cells, which belong to three different classes( Reference Finger 24 , Reference Chaudhari and Roper 25 ): type I (glial-like) cells, which seem to be involved in the clearance of neurotransmitters through absorption/degradation; type II (receptor) cells, which express G-protein-coupled receptors (including the taste receptors T1R and T2R), which bind to bitter, sweet and umami compounds, the downstream signalling components (e.g. phospholipase C β2 (PLCβ2)) that transduce these taste qualities and the G-protein gustducin( Reference Yang, Tabata and Crowley 26 ); type III (presynaptic) cells, which form synaptic contacts with nerve terminals and are known to receive and integrate signals from type II cells.
Expression of the CaSR has been observed in type III taste cells at the mRNA and protein levels( Reference San Gabriel, Uneyama and Maekawa 22 , Reference Bystrova, Romanov and Rogachevskaja 23 ), where the CaSR appears to be distributed throughout the plasma membrane; however, there have been conflicting reports on whether the CaSR is also expressed in type I and type II cells. Bystrova et al. ( Reference Bystrova, Romanov and Rogachevskaja 23 ) showed CaSR mRNA expression at the single-cell level in a number of type I cells, but not in type II cells, using serial multistandard-assisted reverse transcriptase - polymerase chain reaction (SMART-PCR); however, they were unable to demonstrate functional coupling to the PLC-dependent Ca2+ signalling pathways in type I cells.
The possibility remains that the CaSR signals in a PLC-independent manner in type I taste cells. Type I taste cells express the renal outer medullary K+ channel on their apical membrane, where this channel might play a role in the recycling of the K+ that accumulates in the restricted spaces between type II and type III cells( Reference Dvoryanchikov, Sinclair and Perea-Martinez 27 ). This scenario would be reminiscent of the thick ascending limb of the kidney, where the activation of the basolateral CaSR has been shown to inhibit apical renal outer medullary K+ through signalling pathways involving arachidonic acid and its metabolites( Reference Wang, Lu and Hebert 28 ), and the type I taste cell CaSR may signal in a similar manner (Fig. 2).

Fig. 2 Relationship between calcium-sensing receptor (CaSR) half maximal effective concentration (EC50) values and kokumi taste intensity. A total of six γ-glutamyl peptides were tested for kokumi taste intensity by a panel of assessors. The intensity of kokumi taste was quantified in reference to the glutathione (GSH) concentration required to achieve an equivalent intensity of taste sensation. EC50 values for these substances were determined by measuring agonist-evoked increase in intracellular Ca2+ concentrations in HEK-293 cells transiently expressing human CaSR. Substances with stronger kokumi taste intensity exhibited a higher potency for CaSR activation than substances with lower kokumi taste intensity. Data were obtained from Ohsu et al. ( Reference Ohsu, Amino and Nagasaki 31 ) and redrawn. (A colour version of this figure can be found online at http://www.journals.cambridge.org/bjn)
Conversely, San Gabriel et al. ( Reference San Gabriel, Uneyama and Maekawa 22 ) and Maruyama et al. ( Reference Maruyama, Yasuda and Kuroda 29 ) have demonstrated expression of the CaSR in a subset of taste cells that express either neural cell adhesion molecule (NCAM) (a marker of type III cells) or PLCβ2 (a marker of type II cells) using immunofluorescence. CaSR-positive, PLCβ2-expressing type II cells did not express one of the subunits required for the sweet/umami taste receptors, T1R3( Reference Maruyama, Yasuda and Kuroda 29 ); however, whether other receptors, such as the bitter T2R receptor, are co-expressed in CaSR-positive type II taste cells is currently unknown.
Interestingly, recent work(
Reference Rogachevskaja, Churbanov and Bystrova
30
) has demonstrated that in CaSR-expressing HEK-293 cells this receptor may be stimulated by the bitter compound denatonium in the millimolar range. Similar to other small peptides, such as glutathione and γ-glutamyl peptides(
Reference Wang, Yao and Kuang
15
,
Reference Broadhead, Mun and Avlani
16
), it seems to have a positive allosteric effect on
$$Ca _{o}^{2 + } $$
concentration–response curves, although whether denatonium stimulates the CaSR in taste cells is unknown(
Reference Rogachevskaja, Churbanov and Bystrova
30
).
The exposure of CaSR-expressing type III taste cells to l-amino acids (l-Phe and Arg), γ-glutamyl peptides (such as glutathione and γ-glutamyl-valine-glycine) and calcimimetics (cinacalcet) has been shown to evoke intracellular Ca2+(
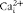 $$Ca _{i}^{2 + } $$
) transients, which are ablated by the non-specific PLC inhibitor U73122(
Reference Bystrova, Romanov and Rogachevskaja
23
,
Reference Maruyama, Yasuda and Kuroda
29
). High concentrations (3 μm) of the negative CaSR allosteric modulator, the ‘calcilytic’ NPS 2143, have also been shown to inhibit γ-glutamyl-valine-glycine-mediated
$$Ca _{i}^{2 + } $$
responses, suggesting that these responses might be mediated through the activation of the CaSR (Fig. 2)(
Reference Maruyama, Yasuda and Kuroda
29
).
$$Ca _{i}^{2 + } $$
) transients, which are ablated by the non-specific PLC inhibitor U73122(
Reference Bystrova, Romanov and Rogachevskaja
23
,
Reference Maruyama, Yasuda and Kuroda
29
). High concentrations (3 μm) of the negative CaSR allosteric modulator, the ‘calcilytic’ NPS 2143, have also been shown to inhibit γ-glutamyl-valine-glycine-mediated
$$Ca _{i}^{2 + } $$
responses, suggesting that these responses might be mediated through the activation of the CaSR (Fig. 2)(
Reference Maruyama, Yasuda and Kuroda
29
).
Although the majority of this work has been completed in rodent taste cells, there is evidence that the CaSR plays a role in human taste transduction. Human sensory analysis has demonstrated that a number of CaSR activators, including glutathione and γ-glutamyl-valine-glycine, act as kokumi taste substances( Reference Ohsu, Amino and Nagasaki 31 ), enhancing sweet, salty and umami tastes without producing a taste of their own. There seems to be a positive correlation between kokumi taste intensity and CaSR agonist activity, as determined by the half maximal effective concentration (EC50) values in CaSR-expressing HEK-293 cells (Fig. 3, modified from that of Ohsu et al. ( Reference Ohsu, Amino and Nagasaki 31 )). In addition, high concentrations of the calcilytic NPS 2143 have been found to significantly suppress the kokumi taste effects of CaSR agonists( Reference Ohsu, Amino and Nagasaki 31 ).

Fig. 3 Proposed roles of the calcium-sensing receptor (CaSR) within taste cells. This diagram depicts the proposed roles and signalling pathways for the CaSR in type I, II and III taste cells. In this figure, the CaSR is positioned on the membrane that is most appropriate for its postulated role. (1) In type I (glial-like) cells, activation of the basolateral CaSR may be linked to the regulation of potassium recycling by the apical renal outer medullary potassium (ROMK) channel, similar to its role in the kidney. (2) The CaSR is most probably co-expressed in type II (receptor) cells with T2R receptors on the luminal membrane, where it may play a role in the transduction of both bitter taste (oral Ca2+/denatonium) and kokumi taste (l-amino acids (l-AA)/γ-glutamyl). Activation of CaSR homodimers, or possible CaSR/T2R heterodimers, leads to activation of the G-protein gustducin (αgus) and phospholipase C β2 (PLCβ2). PLCβ2 catalyses the formation of inositol 1,4,5-trisphosphate (IP3) and diacylglycerol (DAG) from phosphatidylinositol 4,5-bisphophate (PIP2), leading to the release of Ca2+ from the endoplasmic reticulum (ER) through IP3 binding to an IP3 receptor (IP3R). Increased intracellular Ca2+ concentrations lead to the depolarisation of the cell through the actions of the Na+ channel transient receptor potential cation channel, subfamily M, member 5 (Trpm5), delayed rectifying potassium channels (KDR channels) and voltage-gated calcium channels (VGCC). Furthermore, the cells release ATP through pannexin 1 (Panx1), exciting the ATP receptors P2Y and P2X on sensory never fibres( Reference Chaudhari and Roper 25 ). (3) In type III presynaptic cells, activation of the apical CaSR by γ-glutamyl peptides leads to an increase in intracellular Ca2+ concentrations by a PLC-dependent pathway. An increase in intracellular Ca2+ concentrations in type III cells is linked to the release of the serotonin 5-hydroxytryptamine (5-HT), which can inhibit type II receptor cells; however, whether this occurs in a CaSR-dependent manner is currently unknown. (A colour version of this figure can be found online at http://www.journals.cambridge.org/bjn)
There may also be an additional role for the CaSR in the oral detection of
$$Ca _{o}^{2 + } $$
, which has a distinctive taste quality, with aqueous Ca-containing solutions tasting quite bitter to humans(
Reference Tordoff
32
). Interestingly, Ca deprivation has been shown to increase the palatability of Ca in rodents(
Reference Tordoff, Shao and Alarcon
33
). The exact mechanism by which animals detect/‘taste’ Ca is still yet to be fully elucidated; however, previous work(
Reference Tordoff, Shao and Alarcon
33
) has implicated the involvement of the T1R3 receptor. It has been suggested that the T1R3 receptor may heterodimerise with the CaSR to form a functional Ca2+ sensor in type II taste cells(
Reference Tordoff, Shao and Alarcon
33
); however, there is no evidence to date that the expression of T1R3 and that of the CaSR overlap(
Reference Bystrova, Romanov and Rogachevskaja
23
,
Reference Maruyama, Yasuda and Kuroda
29
).
As the activation of the CaSR-positive taste cells has already been implicated in the modulation of kokumi taste, it is perhaps not such a great leap to imagine that the CaSR could play a similar role in the detection of oral Ca. In support of this argument is the finding that a substantial proportion of variation in Ca taste preference among mice is due to CaSR polymorphisms( Reference Tordoff, Reed and Shao 34 ).
Calcium-sensing receptor in the intestine
The CaSR is present in the stomach, where its activation stimulates the secretion of gastrin (by G cells) and of H+ (by antral cells) (reviewed by Geibel & Hebert( Reference Geibel and Hebert 35 )). In gastrin-secreting G cells, the CaSR is expressed on both the apical and basolateral membranes( Reference Buchan, Squires and Ring 36 ), suggesting that they have the ability to respond to changes in both the luminal contents and blood( Reference Conigrave and Brown 19 ), whereas in parietal cells, the CaSR is expressed only on the basolateral membrane( Reference Cheng, Qureshi and Chattopadhyay 18 ), which could allow the stimulation of gastric acid secretion by intestinally absorbed l-amino acids( Reference Conigrave and Brown 19 ).
Seminal studies carried out by Geibel & Hebert( Reference Geibel and Hebert 35 ) have demonstrated that the CaSR, expressed on both the apical and basolateral membranes of colonic crypts, plays a fundamental role in the colon in NaCl and water transport and raises the possibility of using CaSR-based therapeutics to prevent toxin-induced secretory diarrhoea, one of the most debilitating conditions in underdeveloped countries. Luminal Ca2+ also promotes gut epithelial differentiation, while CaSR-mediated signalling suppresses gut cell proliferation while preserving epithelial integrity( Reference Liddle, Goldfine and Rosen 37 ). Dietary Ca2+ intake is associated with a reduced risk of colon cancer and CaSR expression is absent in colon cancer specimens while being highly abundant in normal tissue from the same patients (see Rogers et al. ( Reference Rogers, Hanly and Collins 38 ) for a review). While a definitive and direct link between loss of CaSR expression and malignant transformation in the gut remains to be elucidated, it has been hypothesised that the CaSR might be a drug target for the treatment of colon cancer.
Recent evidence has demonstrated that the CaSR also plays an important role in the regulation of hormone secretion in intestinal enteroendocrine cells in response to l-amino acids. Aromatic amino acids have been shown to mediate the secretion of the GI hormone cholecystokinin (CCK)( Reference Furuse, Chol and Yang 39 – Reference Konturek, Radecki and Thor 43 ). The secretion of CCK from enteroendocrine cells in the small intestine is a major regulator for the release of bile by the gall bladder, as well as the secretion of digestive enzymes from the pancreas( Reference Liddle, Goldfine and Rosen 37 , Reference Yang, Furuse and Sugishita 42 ). CCK also acts as a satiety hormone, reducing food intake in various species, including humans( Reference Ballinger and Clark 44 ).
Due to the difficulties in obtaining sufficient amounts of homogeneous I cells from intestinal tissue, initial experiments examining the cellular mechanism by which aromatic amino acids mediate CCK secretion had focused on the murine enteroendocrine cell line STC-1. Here, l-Phe was shown to stimulate CCK secretion in a
$$Ca _{o}^{2 + } $$
-dependent manner (Fig. 4(a) and (b)) (
Reference Mangel, Prpic and Wong
45
). l-Phe also increased
$$Ca _{i}^{2 + } $$
concentrations and Ca2+ channel activity, while the Ca2+ channel blocker diltiazem inhibited CCK secretion(
Reference Mangel, Prpic and Wong
45
). Phe-mediated secretion is also stereoselective for the natural l-isomer (Fig. 4(a)). High concentrations of the calcilytic NPS 2143 abolished l-Phe-stimulated CCK secretion, suggesting that the CaSR may play a role in l-Phe mediated CCK secretion in STC-1 cells. Consistent with this hypothesis, CaSR mRNA expression was detected in STC-1 cells using RT-PCR(
Reference Hira, Nakajima and Eto
46
). Recent work(
Reference Nakajima, Hira and Hara
47
) has also demonstrated that the STC-1 cells respond to protein hydrolysates (such as egg albumin, meat, potato, casein and soyabean) with an increase in CCK secretion, which is suppressed in the presence of NPS 2143, except for meat hydrolysate-induced CCK secretion. Together, these studies strongly support a role for the CaSR as an amino acid sensor in these cells.

Fig. 4
l-Amino acid-induced cholecystokinin (CCK) secretion from STC-1 cells. (a) Stimulation of CCK secretion in response to l-Phe, l-Trp and d-Phe at 1·5 mm-
$$Ca _{o}^{2 + } $$
(■) or 3·0 mm-
$$Ca _{o}^{2 + } $$
(■) concentrations in STC-1 cells. Both the Ca-sensing receptor-active aromatic amino acids l-Phe and l-Trp stimulated increases in CCK secretion in a calcium-dependent manner. CCK secretion was stereoselective, as exposure to d-Phe had a minimal effect on CCK secretion. (b) Concentration-dependent CCK secretion in STC-1 cells. Increasing concentrations of both
$$Ca _{o}^{2 + } $$
and l-Phe induced increases in CCK secretion in a concentration-dependent manner in STC-1 cells. ■, 1·0 mm-Ca2+; □, 1·5 mm-Ca2+; ![]() , 2·0 mm-Ca2+;
, 2·0 mm-Ca2+; ![]() , 2·5 mm-Ca2+. For (a) and (b), STC-1 cells were exposed to various agonists at different
$$Ca _{o}^{2 + } $$
concentrations (1–3 mm) for 30 min at 37°C before CCK secretion was determined using a commercial CCK enzyme immunoassay kit (Phoenix Pharmaceuticals, Inc.). Values are % mean change from baseline, with their standard errors from three to four wells represented by vertical bars.
, 2·5 mm-Ca2+. For (a) and (b), STC-1 cells were exposed to various agonists at different
$$Ca _{o}^{2 + } $$
concentrations (1–3 mm) for 30 min at 37°C before CCK secretion was determined using a commercial CCK enzyme immunoassay kit (Phoenix Pharmaceuticals, Inc.). Values are % mean change from baseline, with their standard errors from three to four wells represented by vertical bars.
The involvement of the CaSR in the mediation of CCK secretion has been elucidated by two groups using bacterial artificial chromosome transgenic mice, which permitted the identification of specific cell types using enhanced green fluorescent protein (eGFP). This elegant approach has permitted the isolation of CCK-secreting cells from CCK–eGFP bacterial artificial chromosome transgenic mice by fluorescence-activated cell sorting( Reference Liou, Sei and Zhao 48 , Reference Wang, Chandra and Samsa 49 ).
The first such study, carried out by Liou et al.
(
Reference Liou, Sei and Zhao
48
), used fluorescence-activated cell sorting to isolate native CCK-secreting duodenal I cells from CCK–eGFP bacterial artificial chromosome mice. Using quantitative RT-PCR, these isolated CCK–eGFP cells were shown to express CaSR mRNA transcripts at a level approximately 900-fold higher than that in non-eGFP-expressing cells, and the presence of CaSR protein was confirmed with immunofluorescence. The exposure of isolated I cells to phenylalanine induced intracellular Ca2+ influx, which was
$$Ca _{o}^{2 + } $$
dependent and stereoselective for l-Phe. l-Phe-dependent CCK secretion in native I cells was enhanced in the presence of supraphysiological
$$Ca _{o}^{2 + } $$
concentrations, indicating a synergistic effect of
$$Ca _{o}^{2 + } $$
and this amino acid. Interestingly, supraphysiological
$$Ca _{o}^{2 + } $$
concentrations alone were unable to evoke an increase in CCK secretion. Based on these results, the authors concluded that the CaSR acts as an amino acid sensor in this physiological setting(
Reference Liou, Sei and Zhao
48
).
Deletion of the CaSR from CCK–eGFP I cells did not affect basal CCK secretion; however, l-Phe-mediated
$$Ca _{i}^{2 + } $$
influx was lost. Furthermore, l-Phe and supraphysiological
$$Ca _{o}^{2 + } $$
concentrations surprisingly suppressed CCK secretion by approximately 20–30 % in these cells, compared with basal levels, suggesting that not only is the CaSR required for l-Phe-mediated CCK secretion, but also the absence of a fully functional receptor may inhibit l-amino acid-induced CCK secretion(
Reference Liou, Sei and Zhao
48
).
The second study, carried out by Wang and colleagues, examined CCK-secreting intestinal mucosal cells in CCK–eGFP bacterial artificial chromosome transgenic mice. Expression of the CaSR was confirmed with quantitative RT-PCR and immunofluorescence and was found to be localised in both the apical and basolateral regions of CCK–eGFP cells, similar to that shown in the study carried out by Liou et al
(
Reference Liou, Sei and Zhao
48
). Aromatic amino acids l-Phe and l-Trp, but not the non-aromatic amino acid l-Ala, caused transient increases in
$$Ca _{i}^{2 + } $$
concentrations and stimulated CCK secretion. Antagonisation of the CaSR with the calcilytic Calhex 231 blocked aromatic amino acid-mediated CCK secretion, without affecting the effect of hyperpolarising concentrations of KCl, again pointing to a role for the CaSR in the modulation of the effects of certain amino acids on CCK secretion(
Reference Wang, Chandra and Samsa
49
).
Recently, the CaSR has also been implicated in the regulation of K- and L-cell activity in response to l-amino acids( Reference Mace, Schindler and Patel 50 ). Isolated loops of the rat small intestine were used to quantify the secretion of three anti-diabetic gut peptides (gluco-indulinotropic peptide, glucagon-like peptide 1 and peptide tyrosine tyrosine) in response to a number of l-amino acids.
l-Phe, l-Trp, l-Arg, l-Asn and l-Gln induced the secretion of gluco-indulinotropic peptide, glucagon-like peptide 1 and peptide tyrosine tyrosine in the presence of physiological
$$Ca _{o}^{2 + } $$
concentrations (i.e. 1·25 mm). Characteristic of a CaSR-mediated response, l-amino acid-induced secretion responses were abolished in the absence of
$$Ca _{o}^{2 + } $$
for all the three peptides. High concentrations of the CaSR antagonist Calhex 231 suppressed l-amino acid secretion responses to various degrees, with the exception of l-Gln-stimulated glucagon-like peptide 1 secretion(
Reference Mace, Schindler and Patel
50
). Inhibition of the CaSR by Calhex 231 was most efficient at suppressing aromatic amino acid responses(
Reference Mace, Schindler and Patel
50
), perhaps unsurprisingly as it is the most potent CaSR ligand(
Reference Conigrave, Quinn and Brown
13
). Furthermore, the addition of a CaSR allosteric agonist, NPS R568, after the initial elevation of gluco-indulinotropic peptide, glucagon-like peptide 1 or peptide tyrosine tyrosine secretion by l-amino acids further enhanced secretion to a maximal level, while an increase in
$$Ca _{o}^{2 + } $$
concentrations increased the potency of l-Phe-induced L-/K-cell response.
Summary and conclusion
The role of the CaSR in nutrient sensing within the GI system continues to evolve with time. Previous studies( Reference Kerstetter, O'Brien and Insogna 51 , Reference Kerstetter, Caseria and Mitnick 52 ) have demonstrated that a protein-rich diet improves bone health and is associated with a reduced risk of fracture and an improved post-fracture recovery, underlying a link between dietary protein intake and Ca metabolism. New developments, presented in this review article, implicate the involvement of the CaSR in the modulation of appetite and control of satiety and anti-diabetic hormone secretion in response to amino acids/dietary Ca. Overall, these findings present the CaSR as a possible new therapeutic target in the ongoing fight against obesity and osteoporosis and their related disorders.
Acknowledgements
The authors are grateful to Professor Arthur Conigrave and Professor Steve Simpson from the University of Sydney for allowing the inclusion of previously unpublished CCK secretion data obtained by S. C. B. in their laboratory. They acknowledge the Marie Curie ITN ‘Multifaceted CaSR’ for providing financial support (grant 264663 to D. R.). T. S. D. is a recipient of a BBSRC-CASE studentship.
The authors' contributions are as follows: S. C. B. prepared the manuscript; T. S. D. and M. S. assisted in revising the manuscript and preparing the figures; D. R. was involved in the preparation and editing of the manuscript. All the authors read and approved the final manuscript.
The authors have no conflicts of interest to declare.





