



Introduction
Animals often face critical decisions throughout their lives, such as choosing food, mates and habitats, which can profoundly impact their survival. Poor decisions in these scenarios can have negative or fatal consequences, driving the evolution of decision-making mechanisms (MacArthur and Pianka, Reference MacArthur and Pianka1966; Stephens, Reference Stephens2008; Bell, Reference Bell2012; Ryan et al., Reference Ryan, Page, Hunter and Taylor2019). In terms of food selection, animals may weigh the energy gained from hunting against the energy expended (MacArthur and Pianka, Reference MacArthur and Pianka1966; Alcaraz and García-Cabello, Reference Alcaraz and García-Cabello2017; Norberg, Reference Norberg2021). The optimal foraging theory (MacArthur and Pianka, Reference MacArthur and Pianka1966; Pyke and Starr, Reference Pyke and Starr2021) elucidates predator feeding behaviour by considering factors such as nutrient concentration and food availability. This theory acknowledges the diversity of food environments, where decisions are influenced by interactions between multiple food components and the forager. Additionally, past experiences and biochemical contexts also shape food choices, affecting present preferences and intake (Villalba et al., Reference Villalba, Provenza and Han2004; Villalba and Provenza, Reference Villalba and Provenza2009; Ventura and Worobey, Reference Ventura and Worobey2013; Chen and Antonelli, Reference Chen and Antonelli2020). Studies on holometabolous insects highlight the impact of larval diet on adult host plant selection (Janz et al., Reference Janz, Söderlind and Nylin2009; Müller and Müller, Reference Müller and Müller2016; Nestel et al., Reference Nestel, Papadopoulos, Pascacio-Villafán, Righini, Altuzar-Molina and Aluja2016). Recent evidence suggests that larval experiences can influence host selection behaviour in adult insects across species (Gandolfi et al., Reference Gandolfi, Mattiacci and Dorn2003; Hora et al., Reference Hora, Roessingh and Menken2005; Olsson et al., Reference Olsson, Anderbrant and Löfstedt2006; Moreau et al., Reference Moreau, Rahme, Benrey and Thiéry2008; Salgado and Saastamoinen, Reference Salgado and Saastamoinen2019).
Coccinellids are generally known to exhibit three main feeding habits, i.e. predation (zoophagy), plant feeding (phytophagy) and fungus feeding (mycophagy) (Giorgi et al., Reference Giorgi, Vandenberg, McHugh, Forrester, Ślipiński, Miller, Shapiro and Whiting2009). Many coccinellid species are known to utilise alternative food sources in addition to their preferred ones, leading to the distinction between ‘essential’ food sources, which are sufficient for larval development and adult oviposition (Hodek et al., Reference Hodek, Honěk and Hodek1996, Reference Hodek, Honek and Van Emden2012), and merely facultative food sources. The use of honeydew, pollen, sap, nectar and various fungi as alternative food sources is widespread among predaceous groups (Sarwar, Reference Sarwar2016; Kundoo and Khan, Reference Kundoo and Khan2017), as is the exploitation of secondary or less favourable prey species (Hodek et al., Reference Hodek, Honěk and Hodek1996; Majerus, Reference Majerus2016). Their prey preference depends on various factors, including prey availability, specificity, capture and handling times, nutritive value, palatability and their contribution to fitness and reduced predation costs (Hodek et al., Reference Hodek, Honek and Van Emden2012; Guroo et al., Reference Guroo, Pervez, Srivastava and Gupta2017; Pervez and Chandra, Reference Pervez and Chandra2018; Canovai et al., Reference Canovai, Benelli, Ceragioli, Lucchi and Canale2019). Previous studies on prey preferences in predaceous ladybirds (Coleoptera: Coccinellidae) typically focus on single prey species and their effects on consumption and fitness (Pervez and Omkar, Reference Pervez and Omkar2004, Reference Pervez and Omkar2006; Omkar and Mishra, Reference Omkar and Mishra2005; Pervez and Chandra, Reference Pervez and Chandra2018; Kumari et al., Reference Kumari, Saifi, Arya and Perves2022; Kumar and Omkar, Reference Kumar, Omkar and Omkar2023). However, preference tests are more appropriate when multiple food sources are available in the same microhabitat, resembling a cafeteria setup (Nedved and Salvucci, Reference Nedved and Salvucci2008; Šenkeříková and Nedvěd, Reference Šenkeříková and Nedvěd2013). Cafeteria experiments, where various food types are equally offered to animals for a set duration, are the most effective method for conducting true food preference studies involving behavioural selection among multiple food types. Hence, it would be interesting to investigate how ladybirds respond when presented with both toxic and nutritious prey simultaneously. However, in the presence of more food choices, the beetle may end up confused and the probability of choosing a poor food may increase (Nedved and Salvucci, Reference Nedved and Salvucci2008).
Ecological and nutritional plasticity, high intrinsic rates of increase, short life cycle, presence of sexual dimorphism and most abundance in agroecosystem are the most probable reasons for the selection of Propylea dissecta (Mulsant) (Coleoptera: Coccinellidae) as an experimental model for this study. It is predaceous in nature and preferably consume aphids followed by whiteflies (Pervez and Omkar, Reference Pervez and Omkar2004; Omkar and Mishra, Reference Omkar and Mishra2005; Hodek et al., Reference Hodek, Honek and Van Emden2012). Although P. dissecta have a diverse diet, their prey range is not as broad as that of other beetles (Pervez and Omkar, Reference Pervez and Omkar2004). Despite its potential as a predator, the smaller size of P. dissecta prevents it from effectively preying on a vast range of organisms. To test, if more food choices lead to bad results, we hypothesised that on being provided with a variety of food choices, P. dissecta will show poor food selection, which in turn will negatively affect their mating and reproductive parameters. This study was focused on gaining a deeper understanding of how ladybirds make decisions. It explores the strategies they employ when faced with various food choices, aiming to uncover patterns or preferences in their decision-making process.
Material and methods
Stock culture
Adults of Cheilomenes sexmaculata Fabricius (Coleoptera: Coccinellidae) and P. dissecta (n = 100 of each) were collected from the agricultural fields surrounding Lucknow, India (26°50′N 80°54′E). Adults were mated and placed in Petri dishes (hereafter, 9.0 × 2.0 cm unless mentioned) in temperature-controlled chambers maintained at 25 ± 2°C, 65 ± 5% RH, 14L:10D photoperiod. They were provided with ad libitum daily replenished supply of Aphis craccivora Koch (Hemiptera: Aphididae), infested on Vigna unguiculata Linnaeus reared in a polyhouse (25 ± 2°C, 65 ± 5% RH). The eggs laid were collected every 24 h and incubated under the above abiotic conditions until hatching. After hatching, the first instar larvae were removed using a fine camel hair paint brush and assigned individually to clean experimental Petri dishes (9.0 × 2.0 cm) and provided with ad libitum supply of aphids. To acclimatise to laboratory conditions, rearing was done for one more generation. Thereafter, the requisite stages were used for experimentation.
Food collection for dietary choice
For the food choice experiment, five different foods, heterospecific eggs, conspecific eggs, mixed pollen grains, Aphis nerii Boyer de Fonsclombe and A. craccivora were provided. For the collection of conspecific and heterospecific eggs, a pair of 10-day-old male and female (n = 50 pairs) of both species, i.e. P. dissecta and C. sexmaculata, were taken from stock and allowed to mate in plastic Petri dishes under abiotic conditions mentioned above and provided with ad libitum prey. The females were isolated post-mating in Petri dishes (biotic and abiotic factors as above) and fresh eggs laid were collected daily. Only fresh eggs were given as food choices in the experiment. Aphids, A. craccivora and A. nerii, were collected from their host plants V. unguiculata and Calotropis procera (Aiton), respectively. Calotropis procera infested with A. nerii were collected from local areas of Lucknow, India and kept in the temperature-controlled chambers maintained at 25 ± 2°C, 65 ± 5% RH, 14L:10D photoperiod. Both the aphids were provided on the leaves of their host plants during food choice treatments to restrict their mobility. Bee-collected mixed pollen grains used in the experiment were purchased from Royal Honey and Bee Farming Products (Nitinsbees), Chinhat, Lucknow, Uttar Pradesh, India.
Optimisation of food quantity in each dietary regime
Standardisation studies were conducted to evaluate the food quantity to be provided in each dietary regime and these food regimes were grouped as: (i) preferred aphid, A. craccivora, (ii) non-preferred aphid, A. nerii, (iii) freshly laid heterospecific eggs, (iv) freshly laid conspecific eggs and (v) pollen grains. In both the aphid dietary regimes, the first, second and third instars of P. dissecta were provided with 6–12 second and third instars of aphids per day and for the fourth instar, 10–20 gravid aphids were provided (Singh et al., Reference Singh, Mishra and Omkar2016). In the egg dietary regime, the number of eggs provided differed with instars: first instar 20 eggs, second instar 40 eggs, third instar 60 and fourth instar with 100 eggs (Yadav et al., Reference Yadav, Omkar and Mishra2019). In the pollen grains dietary regime, crushed pollen grains were initially provided to the instars, but they were found to be insufficient for larval development. Subsequently, we provided the pollen grains along with a cotton ball soaked in honey syrup. However, we also observed the highest mortality of larvae in this regime. Out of 378 larvae, only 49 survived on this diet. From these 49, we randomly selected 30 larvae for further experimentation in the pollen dietary regime. Throughout the study, we referred to the diet consisting of pollen along with honey syrup as the pollen grain dietary regime.
Experimental design
Freshly laid eggs of P. dissecta were collected from stock culture and incubated under the above-mentioned conditions until hatching.
Food choice trials
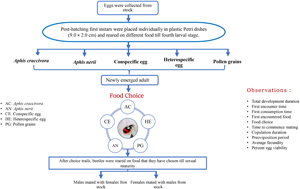
To test, if more food choices lead to poor food selection, immediately after hatching first instars were placed individually in Petri dishes and divided into following dietary groups for further rearing: (i) Aphid, A. craccivora, (ii) Aphid, A. nerii, (iii) freshly laid heterospecific eggs, (iv) freshly laid conspecific eggs and (v) pollen grains. Optimum food in each dietary group was provided and replenished daily (involves carefully transferring fresh food into a clean Petri dish) until pupation. Soon after emergence, adults were individually placed in large plastic Petri dish (14.5 cm diameter × 2.0 cm height) containing the food options, i.e. (i) A. craccivora, (ii) A. nerii, (iii) conspecific eggs, (iv) heterospecific eggs and (v) pollen grains, placed equidistantly (fig. 1). For placing food at equidistance, Petri dish was divided into five sections at 72° angles, had its radius measured at 7.25 cm, and food items were placed 3.6 cm from the centre along each marked line to ensure equidistance from both the centre and each other.

Figure 1. Diagrammatic representation of food choice trial setup.
Total developmental duration (development duration from egg to adult emergence), first food encountered (food which was encountered first), time to encounter first food (time from initially placing beetles into the experimental Petri dish until they first encounter food), first food choice (food which was consumed first) and time taken to consume first food (time taken elapsed from the initial encounter with the food to the first consumption of the food) were recorded. All the treatments were replicated 30 times.
Effect of preferred diet quality on mating and reproductive parameters
After food choice experiment, adults (both males and females) were maintained on the same diet they had chosen as their first preference for the next 10 days. Upon reaching sexual maturity, they were paired with opposite-gender adults of the same age from the stock culture under the above-mentioned abiotic conditions. The females in the choice treatment group were paired with males of the same age from the stock culture and the males from the choice experiment were paired with females of the same age from the stock culture. Mating parameters, i.e. time to commence mating (time taken by beetles to start mating behaviours after being introduced into a Petri dish) and copulation duration (time during which mating occurs between beetles), and reproductive parameters, i.e. fecundity (number of eggs laid by beetles) and per cent egg viability (percentage of eggs that are capable of developing into viable offspring), were recorded in all the replicates.
Statistical analysis
Data on total development duration, time to encounter the first food and time to consume the first food were first tested for normality (Kolmogorov–Smirnoff test). On being found non-normally distributed, Generalised Linear Model (GLM), using gamma log link function depending on the skewness of the data, was applied. To analyse the effect of the dietary regime on total development duration, the latter was used as a response factor and the former as a fixed factor in the GLM. To study the effect of multiple food choices on the time to encounter the first food, encounter time was taken as response factors and dietary regime and first food encountered were used as fixed factors in GLM. To reveal the effect of multiple food choices on the time to consume the first food, first consumption time was used as response factors and dietary regime and first food consumed were taken as fixed factors in GLM.
Chi-square (χ2) goodness-of-fit analysis was used to analyse the first encountered food and first consumed prey (food choice) by the newly emerged adult. To test the effect of food choice on mating and reproductive parameters of adults, data on time to commence mating, copulation duration, oviposition period, average fecundity and per cent egg viability were first tested for normality, and on getting non-normally distributed, GLM, using gamma log link function depending on the skewness of the data, was applied. Data on time to commence mating, copulation duration, oviposition period, average fecundity and per cent egg viability were taken as response factors and dietary regime and food choice were used as fixed factors in separate GLMs.
All the analyses were carried out by using SPSS 20 software, version 20.0 (IBM, Armonk, New York, USA).
Results
Effect of dietary regime on total development duration
The total developmental duration was significantly affected by the diet (F = 781.271, df = 4, P = 0.000). Shortest development duration was recorded on A. craccivora, whereas longest duration was found on mixed pollen grains (fig. 2)

Figure 2. Total development duration (in days) in each dietary regime. Values are mean ± SE. Different letters indicate comparison of means among different crowding regimes.
Food choice trials
Time to encounter first food was insignificantly affected by diet (F = 2.523, df = 4, P = 0.640); irrespective of the type of food first encountered (F = 4.150, df = 4, P = 0.386). The interaction between the dietary regime and food first encountered was also insignificant (dietary regime × first food encountered: F = 21.144, df = 16, P = 0.173) (fig. 3). First encountered food was insignificantly (χ2 = 21.446, df = 16, P = 0.162) affected by the dietary regime of the larvae. First encounters with food were random (fig. 4).

Figure 3. First encounter time (in seconds) in each dietary regime.
The box plots show the median. The whiskers extend to the minimum and maximum value. Circles and asterisk represent the outliers.

Figure 4. First food encountered in each dietary regime.
The time to consume the first food was insignificantly affected by both the dietary regime (F = 5.476, df = 4, P = 0.242) and the first food consumed (F = 1.341, df = 4, P = 0.854). Their interaction was also found to be insignificant (F = 14.179, df = 16, P = 0.585). Similar consumption time was recorded in each dietary regime (fig. 5). Food first consumed was also found insignificant across dietary regimes (χ2 = 15.671, df = 16, P = 0.476). In each dietary regime, the first consumed food was either A. craccivora or A. nerii (fig. 6)

Figure 5. First consumption time (in seconds) to consume first food in each dietary regime.
The box plots show the median. The whiskers extend to the minimum and maximum value. Circles and asterisk represent the outliers.

Figure 6. First consumed food in each dietary regime.
Effect of selected diet quality on mating and reproductive parameters
Here, we assessed the effect of selected food on beetles' mating and reproductive parameters. For male, the time to commence mating was significantly (F = 14.363, df = 4, P = 0.006) affected by the food choice they made on the day of their emergence but it was not influenced by their larval dietary regime (F = 2.521, df = 4, P = 0.641). The interaction of the first food choice and dietary regime was significant (F = 62.729, df = 14, P = 0.000). Copulation duration was significantly (F = 9.944, df = 4, P = 0.041) affected by the dietary regime but not by food choice (F = 5.128, df = 4, P = 0.274). However, the interaction between the two was significant (F = 27.065, df = 14, P = 0.019) (table 1).
Table 1. Effect of dietary regime and food choice on mating parameters of male Propylea dissecta

Values are mean ± SE. Letters indicate comparison of means among different dietary regimes, respectively. Means that do not share a letter are significantly different (P > 0.05).
Pre-oviposition period was insignificantly (F = 3.552, df = 4, P = 0.470) influenced by the food choice but significantly (F = 15.625, df = 4, P = 0.004) influenced by the dietary regime. The interaction of food choice and dietary regime (F = 42.327, df = 14, P = 0.000) was significant. Average fecundity was significantly affected by both dietary regimes (F = 46.628, df = 4, P = 0.000) and food choice (F = 15.882, df = 4, P = 0.003) and their interaction was also significant (F = 60.022, df = 14, P = 0.000). Per cent egg viability was also found to be significantly affected by both the dietary regime (F = 64.871, df = 4, P = 0.000) and the food choice (F = 16.966, df = 4, P = 0.002) and their interaction was also significant (F = 85.864, df = 14, P = 0.000) (table 2).
Table 2. Effect of dietary regime and food choice on reproductive parameters of male Propylea dissecta

Values are mean ± SE. Letters indicate comparison of means among different dietary regimes, respectively. Means that do not share a letter are significantly different (P > 0.05).
For the female, the time to commence mating was insignificantly (F = 2.129, df = 4, P = 0.712) affected by the food choice but was significantly (F = 9.697, df = 4, P = 0.046) affected by the dietary regime. The interaction between food choice and the dietary regime was insignificant (F = 8.600, df = 10, P = 0.570). Copulation duration was significantly influenced by both dietary regime (F = 38.298, df = 4, P = 0.000) and food choice (F = 10.962, df = 4, P = 0.027). Their interaction was also found to be significant (F = 50.980, df = 10, P = 0.000) (table 3).
Table 3. Effect of dietary regime and food choice on mating parameters of female Propylea dissecta

Values are mean ± SE. Letters indicate comparison of means among different dietary regimes, respectively. Means that do not share a letter are significantly different (P > 0.05).
Pre-oviposition period was insignificantly influenced by both dietary regime (F = 7.088, df = 4, P = 0.131) and food choice (F = 1.557, df = 4, P = 0.816). Their interaction was also insignificant (F = 8.827, df = 10, P = 0.549). Average fecundity was significantly influenced by dietary regime (F = 11.152, df = 4, P = 0.025) but it was not affected by the food choice (F = 3.008, df = 4, P = 0.556). Their interaction was also found to be insignificant (F = 11.983, df = 10, P = 0.286).
The per cent egg viability was also found to be insignificantly (F = 4.625, df = 4, P = 0.328) affected by the food choice but was significantly affected by the dietary regime (F = 14.554, df = 4, P = 0.006). Their interaction was also found to be insignificant (F = 12.664, df = 10, P = 0.243) (table 4).
Table 4. Effect of dietary regime and food choice on reproductive parameters of female Propylea dissecta

Values are mean ± SE. Letters indicate comparison of means among different dietary regimes, respectively. Means that do not share a letter are significantly different (P > 0.05).
Discussion
The present study was conducted to see the effect of multiple food choices on food selection ability of the ladybird beetle and consequently affecting their reproductive physiology. However, our hypothesis was not supported, as ladybird beetles showed a clear preference for natural prey, specifically aphids. The food choices made by male beetles upon emergence had an impact on the time to commence mating, average fecundity and the per cent egg viability. Additionally, factors such as copulation duration and pre-oviposition period were influenced by previous dietary regime. The females' previous dietary regime influenced all reproductive and mating parameters, while food choice made after emergence affected only copulation duration.
We used total developmental duration as an indicator of food suitability for P. dissecta. A decrease in the developmental duration of P. dissecta indicates a decrease in food suitability, which in this case was A. craccivora>A. nerii>conspecific eggs>heterospecific eggs>pollen grains. A possible reason for the observed decrease in the developmental duration of P. dissecta could be the varying nutritional quality of different food sources provided. Specifically, A. craccivora appears to be the most suitable food, providing optimal nutrition and leading to shorter developmental duration. In contrast, A. nerii, conspecific eggs, heterospecific eggs and pollen grains offer progressively lower nutritional value or less efficient energy conversion, resulting in longer developmental durations. Earlier studies have indicated the least suitability of A. nerii for P. dissecta (Pervez and Omkar, Reference Pervez and Omkar2004; Omkar and Mishra, Reference Omkar and Mishra2005). Osawa (Reference Osawa2002) reported the faster development of immature stages of H. axyridis on the diet of conspecific eggs as compared to a non-cannibalistic diet. A similar result was reported in our finding as well where faster development of larvae on conspecific eggs over heterospecific eggs was recorded. Between the conspecific and heterospecific eggs, conspecific eggs were more nutritious in the larval stages than the heterospecific eggs (Kajita et al., Reference Kajita, Obrycki, Sloggett and Haynes2010; Singh et al., Reference Singh, Mishra and Omkar2020). Additionally, it has been suggested that P. dissecta slows down its development on heterospecific eggs, possibly because they contain toxic alkaloids (Pervez et al., Reference Pervez, Chandra and Kumar2021). Beetles reared on pollen grains exhibited delayed development suggesting that pollen grains may serve as an alternative food source (De Lima et al., Reference De Lima, Pontes and de Lucena Nóbrega2020). Following our findings, Farag et al. (Reference Farag, Abd El-Wahab and Abdel-Moniem2011) reported bee pollen as the least suitable diet for Coccinella undecimpunctata Linnaeus (Coleoptera: Coccinellidae) larvae, among seven different combinations of diets tested. Similar results were reported for the larva of Coccinella transversalis Fabricius (Coleoptera: Coccinellidae), which developed more efficiently when fed honey and mealy bugs compared to pollen or sugar syrups (Maurice et al., Reference Maurice, Kumar and Ramteke2012).
First encounter time was random with all types of food. Even the first food encountered was found to be random. Similar findings were also reported for H. axyridis where both adults and larvae searched randomly for prey-location (Canovai et al., Reference Canovai, Benelli, Ceragioli, Lucchi and Canale2019). This behaviour could be attributed to two factors: (1) the arrangement of food, and (2) the type of food. On placing the newly emerged adult in the middle of the arena surrounded by the variety of foods enabled the beetle to first do extensive and then intensive search for food. Moreover, due to the presence of hemipteran and non-hemipteran food, i.e. conspecific eggs, heterospecific eggs and pollen grains, cues from these food items and the presence of sensory abilities in predators helped them to do extensive and then intensive search for all the food items. Earlier similar findings were reported that on locating their prey, beetles tend to do intensive search (YunDing et al., Reference Yunding, Gaochao, Qinglei, Jiguang and Gongming1997; Hinkelman, Reference Hinkelman2012). Being a generalist predator, P. dissecta not only move towards hemipterans but also towards non-hemipteran food types. It was reported that in dense prey colony, beetles walk leisurely (Murdie, Reference Murdie1971) and Hodek et al. (Reference Hodek, Honek and Van Emden2012) pointed them as ‘blundering idiots’. Studies into prey preference have shown that ladybirds often exhibit a preference for specific types of prey (Guroo et al., Reference Guroo, Pervez, Srivastava and Gupta2017; Pervez and Kumar, Reference Pervez and Kumar2017). This preference is significantly influenced by the host plant, as aphids have been observed to sequester allelochemicals from their host plants, which they then utilise as a defence mechanism against ladybirds.
Time to consume first food and first food consumed both were found to be unaffected by the previous dietary regimes. In each dietary treatment, adults tend to prefer their natural food over non-hemipteran diets. This indicated the choosiness of newly emerged adult P. dissecta. Even on being provided with a variety of food choices, they were found to be rigid in their food selection. Aphids being general nursery prey for the aphidophagous ladybird beetle, P. dissecta, were preferred the most. On the other hand, pollen grains, conspecific and heterospecific eggs were least preferred in each dietary regime possibly due to the presence of less nutritive contents (Farag et al., Reference Farag, Abd El-Wahab and Abdel-Moniem2011), insufficient immature stage of nutrition and toxic allelochemicals (Pervez et al., Reference Pervez, Chandra and Kumar2021), respectively. Overall comparison in different dietary regimes indicated food choice for aphids only. Thus, it suggests that the availability of diverse food options did not lead to poor food choices in P. dissecta and irrespective of the choices offered, beetles preferred their natural prey.
In males, the time to commence mating was significantly influenced by their post-emergence food choice rather than their dietary regime. The prolonged time to commence mating in males with different food choices was indicative of them being not preferred by females reared on A. craccivora. This also suggested that females were able to identify the males based on their dietary history. The dietary history of male ladybird beetles significantly impacts their attractiveness to females through visual, chemical and behavioural cues. Females recognise these cues and prefer males that signal a better diet and higher fitness (Xie et al., Reference Xie, De Clercq, Zhang, Wu, Pan and Pang2015a) For instance, male crickets that have low nutritional concentrations or are provided with low-quality food are perceived as less attractive mates by females (Wilder and Rypstra, Reference Wilder and Rypstra2007). In Cryptolaemus montrouzieri Mulsant (Coleoptera: Coccinellidae) female beetles exhibited a preference for mating with males that were raised with an abundant nutritional diet, regardless of the nutritional conditions they encountered during their larval development (Xie et al., Reference Xie, De Clercq, Pan, Li, Zhang and Pang2015b).
In females, we recorded the opposite results. The time to commence mating was not impacted by the food choice but was affected by the dietary regime. Males reared on A. craccivora and females reared on A. nerii showed a delayed time to commence mating than females of other dietary regimes. It indicated that poor dietary history of females changed the females' attractiveness as mates. Similar to our findings in butterflies and other holometabolous insects, reproduction depends on the reserves accumulated during the larval dietary history (Bauerfeind and Fischer, Reference Bauerfeind and Fischer2005).
Post-emergence food choice by the adults did not affect males' copulation duration but it did in females as the females of food choice treatment were paired with the males of stock culture. This is indicative of the dependency of female copulation duration on food choice. In the recent past, it was observed that mating, being an energy consuming process, alters the food choices in females of P. dissecta (Verma et al., Reference Verma, Mishra and Omkar2023). Interestingly, the copulation duration in both sexes was significantly influenced by the dietary regime. The highest copulation duration was recorded for adults with the larval dietary regime of A. craccivora followed by A. nerii, and conspecific eggs, whereas it was shortest in adults with larval dietary regime of heterospecific eggs and pollen grains. The lower copulation durations of larvae reared on heterospecific eggs and pollen grains can be explained by the fact that they were earlier considered to be toxic (Omkar and Pervez, Reference Omkar and Pervez2011) or insufficient nutrients (Maurice et al., Reference Maurice, Kumar and Ramteke2012). The interaction of food choice and dietary regime significantly affected the copulation duration. Previous studies have reported that food availability significantly affects the individual's attractiveness as potential mates and results in varied copulation duration.
Pre-oviposition period, a crucial phase in the reproductive cycle, remained unaffected by the food choice of adult insects but was notably impacted by the dietary regime of the individuals. This finding suggests that the availability of food sources at the time of adult eclosion does not play a significant role in determining the pre-oviposition period. This finding was consistent with the previous study; it was found that the diet consumed at the moment of eclosion had no discernible effect on factors, such as the pre-oviposition period, daily egg oviposition rates or the viability of egg batches (Hatt and Osawa, Reference Hatt and Osawa2021). Findings underscore the importance of the dietary regime during the larval and developmental stages as a critical determinant of the pre-oviposition period with adult food choices being limited (Dmitriew and Rowe, Reference Dmitriew and Rowe2011). However, when males with different food choice were mated with females reared on A. craccivora, interaction of dietary regime and food choice significantly influenced pre-oviposition period but an insignificant result was observed when females reared on different food choices were mated with males reared on A. craccivora. This could be due to males transferring accessory proteins during mating, which are essential for the pre-oviposition period (Jin and Gong, Reference Jin and Gong2001). Nutritional content present in food affects the production of proteins in the male ejaculate that are important for induction of the post-mating response (Schultzhaus and Carney, Reference Schultzhaus and Carney2017). The longest pre-oviposition period occurred when males reared on A. craccivora in their larval stage switched their diet to heterospecific eggs as adults, while the shortest period was observed when males reared on heterospecific eggs in their larval stage switched their diet to A. nerii as adults. Switching from eggs to aphids provided males with a more balanced diet, reducing the pre-oviposition period, whereas switching from aphids to eggs led to nutritional imbalances, resulting in longer pre-oviposition periods. In contrast, the pre-oviposition period was similar in case of females that were reared on different diets which can be attributed to the fact that the males that mated with these females were reared on A. craccivora.
In our study, we found that both the dietary regime and specific food choices played a significant role in determining the average fecundity and the per cent egg viability in A. craccivora reared females when they were mated with males exhibiting different food choices. There might be two possible reasons for this result, (1) when females reared on a preferred diet and males with different food choices were mated, females enhanced their investment in the offspring. Similar to our findings, Callosobruchus maculatus Fabricius (Coleoptera: Chrysomelidae) females laid more eggs in better food conditions (Iglesias-Carrasco et al., Reference Iglesias-Carrasco, Bilgin, Jennions and Head2018). With different food choices or diets switching from larval to adult stage might lead to male beetles being poorly fed which resulted in enhanced investment of females in fecundity. In consistence with our results, swordtail females also enhanced their investment in offspring size and numbers when mated to poorly fed males (Kindsvater et al., Reference Kindsvater, Simpson, Rosenthal and Alonzo2013). (2) The other possible reason would be that male feeding treatment did not affect female fecundity (Boggs and Freeman, Reference Boggs and Freeman2005). Interestingly, it was found that the dietary history traits of females had a substantial impact on their average fecundity, while the food choices of females did not have a significant influence on their reproductive parameters. The possible reason for this finding could be that females having different food choices might be food deprived perhaps because of food switching from larval to adult stage. This trend was also mirrored in the percentage of egg viability. These observations may be attributed to changes in the dietary regimes of both males and females during their emergence into adulthood, indicating the intricate interplay between food choice, dietary history and the reproductive outcomes of these insects. In decorated crickets, Gryllodes sigillatus Saussure (Orthoptera: Gryllidae) food-limited females, but not males, have been shown to decrease and delay their investment in reproduction (Houslay et al., Reference Houslay, Hunt, Tinsley and Bussiere2015). The insignificant effect of females' food choice and the significant effect of males' food choice on per cent egg viability were recorded; it could be the nutritional needs of male P. dissecta, concerning reproductive viability, are different from the needs of female P. dissecta. Similar to our results, in Podisus maculiventris Say (Hemiptera: Pentatomidae) male nutritional demands are different from the females for the reproductive viability (Wittmeyer et al., Reference Wittmeyer, Coudron and Adams2001).
In conclusion, our results rejected the hypothesis that when offered a variety of food choices P. dissecta will show poor food choice. Contrary to this they consistently select their natural and preferred food. Larval dietary history of adult P. dissecta significantly influenced the development duration, A. craccivora being the best suitable food and pollen grains being the least suitable food for the development of P. dissecta. Dietary history and food choice significantly affected the mating and reproductive parameters, indicating that food availability impacts the individual's attractiveness as potential mates and affects the mating and reproductive behaviour. These findings imply that understanding P. dissecta's dietary preferences and histories is crucial for ecological management and biological control programmes. Future research should explore the mechanisms behind these dietary influences to optimise the use of P. dissecta in pest management.
Acknowledgement
L. V. gratefully acknowledges Jessica Guezen, Global Ecological Change and Sustainability Lab, School of Environmental Sciences, University of Guelph, for improving the language of the paper. L. V. also acknowledges the Department of Zoology, University of Lucknow, Lucknow, Uttar Pradesh, India for providing facilities for this research.
Author contributions
Lata Verma (L. V.), Omkar (O.) and Geetanjali Mishra (G. M.) conceived the idea of the study and L. V. conducted experiment. Data were interpreted by L. V., G. M. and O. Manuscript was written by L. V., G. M. and O. All authors have read and approved the final version of the manuscript.
Financial support
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Competing interests
On behalf of all authors, the corresponding author states that there is no conflict of interest.










