


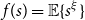

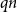
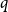
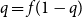

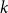
Crossref Citations
This article has been cited by the following publications. This list is generated based on data provided by Crossref.
Heuberger, Clemens
Selkirk, Sarah J.
and
Wagner, Stephan
2024.
The distribution of the maximum protection number in simply generated trees.
Combinatorics, Probability and Computing,
Vol. 33,
Issue. 4,
p.
518.