


INTRODUCTION
Diarrhoeal infections remain a major concern for travellers, especially for those bound for destinations where relatively substandard hygienic conditions exist. A Dutch study showed that in a sample of 1202 individuals travelling to developing countries, 50% experienced one or more diarrhoeal episodes [Reference Belderok1].
Campylobacteriosis is a leading cause of traveller's diarrhoea, particularly in travellers returning from Southern and South East Asia [Reference Ekdahl and Andersson2, Reference Hakanen3]. In The Netherlands (16·5 million population), an estimated 90 000 cases of campylobacteriosis occur annually, with ∼12% estimated as international travel-related [Reference Havelaar4]. Moreover, fluoroquinolone-resistant Campylobacter infections occur significantly more often in travel-related cases [Reference Hakanen3, Reference Evans5].
Travellers are particularly prone to experiencing (symptomatic) Campylobacter infections [Reference Ekdahl and Andersson2, Reference Doorduyn6, Reference Tam7], and susceptibility to disease is associated with duration of foreign stay. For instance, duration of residence of expatriates in Nepal was linearly correlated with protection from diarrhoeal infection [Reference Hoge8], and travellers experiencing multiple diarrhoeal episodes had a shorter duration of symptoms after the first episode [Reference Belderok1]. Similarly, in expatriates in Thailand, campylobacteriosis occurred significantly more often in those living there for <1 year [Reference Gaudio9]. A documented case of acquired immunity in developed countries concerned people professionally exposed. Newly employed poultry abattoir workers in Sweden have been shown to suffer more often from campylobacteriosis than their longer employed colleagues [Reference Cawthraw10]. If partial immunity to (severe) disease is acquired over time with repeated exposure, then such protection should correlate with age. Indeed, campylobacteriosis incidence peaks in infancy worldwide and older age groups are significantly less prone to infection with common Campylobacter strains compared to the young [Reference Miller11]. Moreover, Swedish travellers to countries such as Germany, France, Belgium, The Netherlands, Austria, Luxembourg or Switzerland, all of which are developed countries with high hygienic standards, still have a 4·4–21 times higher risk of acquiring campylobacteriosis compared to those travelling to neighbouring Norway [Reference Havelaar12]. A recent study comparing Campylobacter multilocus sequence typing (MLST) datasets from different countries has further highlighted the importance of geographical distance in strain dissimilarity [Reference Smid13]. Moreover, serological studies of patients and volunteers infected with campylobacters (reviewed in [Reference Havelaar14]) have revealed an array of immunogenic components and elicited antibodies displayed little cross-reactivity, indicating considerable antigenic variation. Although core-genome (as described by MLST) is not necessarily related to antigens, and cross-protection is expected to develop for strains sharing similar antigenic properties, the higher risk of campylobacteriosis in travellers does not appear to be limited just to higher levels of exposure in developing countries, but also to the possible presence of ‘new’ Campylobacter strains, endemic in the different regions to which travellers have rarely been exposed before [Reference Havelaar14]. It follows that, probabilistically, these new strains are more likely to be associated with regionally untested antigens than widespread strains, and acquired protection may be ineffective when exposed to uncommon strains, as evidenced by a Canadian study [Reference Arsenault15].
It is conceivable that the infected, but not necessarily symptomatic [Reference Havelaar14], returning travellers may introduce into the domestic population several ‘exotic’ strains with a higher probability of possessing antigens that are underrepresented in the local reservoirs, i.e. food-producing animals, pets and wildlife. These exotic strains may therefore constitute a distinctive, primarily human-restricted Campylobacter population that may have at least the potential to spread domestically via the person-to-person route. Although Campylobacter person-to-person transmission is believed to be uncommon and up to 66% and 21% of laboratory-confirmed cases in The Netherlands are attributable to chicken and cattle, respectively [Reference Mughini16], it has been shown that campylobacteriosis household outbreaks are more common than believed [Reference Ethelberg17] and up to 18% of putatively household outbreak-related cases are suggestive of secondary spread [Reference Rotariu18]. This raises the question to what extent exotically introduced strains may spill over into the domestic population and at first spread anthroponotically.
In this study, we investigated the MLST profiles of Campylobacter strains isolated from travellers returning to The Netherlands compared to those from domestically acquired cases. We also investigated putative determinants of travel-related campylobacteriosis by comparing the exposures of the returned travellers with those of travellers in the control population. Furthermore, we used a population genetics model for source attribution to estimate the probability that the domestically acquired infections were caused by exotic strains, putatively carried by returned travellers. Finally, factors associated with exotically introduced domestic campylobacteriosis were investigated and the percentage of domestically acquired cases due to person-to-person transmission of exotic strains was estimated.
METHODS
Data
An earlier case-control study on risk factors for campylobacteriosis conducted in The Netherlands between April 2002 and March 2003, the ‘CaSa study’ [Reference Doorduyn6], formed the basis of this study. Isolates of 3115 Campylobacter jejuni/coli cases identified by the Dutch Regional Public Health Laboratories (RPHL) through passive surveillance were sent to the Dutch Central Veterinary Institute (CVI) for molecular speciation [Reference Fermér and Engvall19, Reference Marshall20]. Controls were selected from RPHL population registries by frequency matching (aiming at two per case) according to age (0–4, 5–17, 18–29, 30–44, 45–59, ⩾60 years), sex, urbanization degree (urban: >2500 addresses/km2; urbanized: 500–2500 addresses/km2; rural: <500 addresses/km2) and season (April–June 2002, July–September 2002, October–December 2002, January–April 2003) [Reference Doorduyn6, Reference Mughini16, Reference Doorduyn21]. Cases and controls were asked to complete a questionnaire regarding foreign travel, food consumption, kitchen hygiene, contact with animals, contact with gastroenteritis cases, occupation, recreational activities, medication use and chronic disease history. Questions covered the 7 days prior to symptom onset (cases) or questionnaire completion (controls). Missing values were handled using multiple imputation [Reference Doorduyn6, Reference Doorduyn21].
Cases/controls not returning the questionnaire and/or living abroad were excluded, leaving 1428 cases and 3363 controls enrolled in the study. Of these, 328 cases and 244 controls had travelled abroad with ⩾1 overnight stay in the destination country. A total of 66 countries were visited, with 36 cases and 27 controls visiting >1 country during the same travel episode. For three cases and four controls the travel destination was unknown. Destination countries were grouped into travel regions by adapting the United Nations geoscheme (http://unstats.un.org/unsd/methods/m49/m49regin.htm#europe).
Isolates from 737 non-travellers (domestically acquired cases) and 232 travellers were typed using MLST [Reference Dingle22]. Association of the travellers' five most frequent sequence types (STs) and clonal complexes (CCs) with travel regions was tested using χ 2 or Fisher's exact tests. Proportional Similarity Index (PSI) [Reference Smid13] was used to measure the (dis)similarity between ST frequency distributions of travellers and non-travellers. The PSI ranges between 0 (no common ST) and 1 (identical distributions). Simpson's index of diversity was calculated to define the ST diversity of travellers and non-travellers as the probability that two randomly selected individuals were infected with different STs [Reference Hunter and Gaston23].
Source attribution
The Asymmetric Island (AI) model, a Bayesian population genetics algorithm for modelling Campylobacter evolution and transmission [Reference Wilson24], was used to estimate the probability (Pr) of the 737 non-travellers being infected with exotically introduced STs or with STs originating from four putative animal reservoirs (chicken, cattle, sheep, pig) or from the environment (water, sand, wild birds), a proxy for other unidentified reservoirs putatively of wildlife origin [Reference Mughini16]. This study was restricted to campylobacteriosis of probable exotic origin. Results regarding the other animal and environmental sources have been reported elsewhere [Reference Smid13, Reference Mughini16].
To run the AI model, C. jejuni/coli MLST data from the above-mentioned animal and environmental sources were supplied by the CVI and supplemented with other data [Reference Dingle22, Reference Strachan25, Reference Korczak26] to provide a representative dataset for each source (Table 1). Supplementary data were selected from other published datasets (reported in [Reference Smid13]) using Smid's methodology, which allows for the selection of non-local (and/or non-recent) MLST datasets for Campylobacter source attribution while minimizing potential biases [Reference Smid13, Reference Mughini16]. Differences in Pr for exotic origin (Pr e) were tested for the variables age, sex, and season using Kruskal–Wallis or Mann–Whitney tests.
Table 1. Campylobacter jejuni/coli strains typed with MLST used to feed the Asymmetric Island model for source attribution
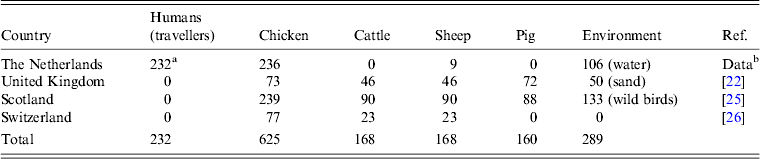
a From Campybobacter jejuni/coli infected travellers of the CaSa study [Reference Doorduyn6].
b Provided by the Central Veterinary Institute (CVI) in Lelystad, The Netherlands.
Factors associated with travel-related campylobacteriosis
Logistic regression was used to investigate putative risk factors for travel-related campylobacteriosis. The 325 diseased travellers and the 238 healthy travellers with known travel destination were included as cases and controls, respectively. Analysis was performed in the same way as in previous studies [Reference Doorduyn6, Reference Mughini16, Reference Doorduyn21]. A total of 131 factors were tested for association with Campylobacter infection in the preliminary single-variable analysis and those factors showing P⩽0·10 were selected for inclusion in a multivariable model. A backward stepwise procedure was applied and variables with P < 0·05 were retained in the final model. Education level [Reference Mughini16], travel region, and length of stay (days) were always included as covariates to control for confounding in addition to the frequency-matched variables. As travel regions were almost mutually exclusive, Western Europe (which The Netherlands belongs to) was made the base category against which the other regions were assessed.
To explore if the risk factors of the reduced model differed according to age, sex, education level, season, and travel region, we also tested for their two-way interactions. The final multivariable model was then expanded to include significant interactions. Overall model significance and goodness-of-fit were verified by likelihood ratio χ 2 and Hosmer–Lemeshow tests, respectively. The best-fitting model was identified using Akaike's Information Criterion (AIC). Bias-corrected bootstrap confidence intervals were also calculated (1000 iterations) and compared to the standard ones. As these did not differ significantly, the standard values were reported. Statistical analysis was performed using Stata v. 11.2 (StataCorp., USA).
Factors associated with exotically introduced domestic campylobacteriosis
To investigate putative risk factors for domestically acquired campylobacteriosis caused by STs of probable exotic origin, MLST data of travellers were included as an additional source in the AI model. Similar to previous studies [Reference Mughini16, Reference Bessell27, Reference Mullner28], the Pr e distribution was assessed and a cut-off was determined to optimize the number of domestic cases assigned to be exotically introduced and the confidence as to their correct assignment derived by the highest possible Pr e. Logistic regression was then used to investigate risk factors for domestically acquired campylobacteriosis caused by STs with at least 77% probability (cut-off Pr e ⩾ 0·77) of originating from abroad. This cut-off Pr e resulted in the selection of 79 cases with a median Pr e of 0·89 (mean 0·88, range 0·77–0·99) belonging to 35 different STs. The 3119 non-travelling controls were included in this analysis.
The effect of the assignment cut-off Pr e on the risk factors was checked by sensitivity analysis, which was done by repeating the analysis for different cut-off Pr e values from 0·5 to 0·9. Low numbers of cases did not allow for the construction of models based on a cut-off of Pr e > 0·9. Finally, a case-case analysis comparing exposures of domestic infections with exotic vs. non-exotic STs was performed.
Estimation of domestic cases due to person-to-person transmission of exotic strains
The percentage of domestic cases due to person-to-person transmission of exotic strains, denoted by γ, was estimated as:
 $${ \gamma =} \displaystyle{{[1 - { \Pi} _{i = 1}^n (1 - {\rm PAR}_i )] \times N_{{\rm ED}}} \over {N_{\rm D}}} \times 100,$$
$${ \gamma =} \displaystyle{{[1 - { \Pi} _{i = 1}^n (1 - {\rm PAR}_i )] \times N_{{\rm ED}}} \over {N_{\rm D}}} \times 100,$$
where PAR is the population attributable risk (calculated based on multivariable odds ratios and prevalence of exposure in cases) for factor i out of n factors entailing person-to-person transmission in the final multivariable model. N ED is the estimated total number of domestic cases of probable exotic origin, given by Binomial(N D, Pr), where N D is the total number of domestic cases (non-travellers) enrolled in the study and Pr is the estimated probability for a domestic case to be infected with an exotic ST, assumed to be Beta(a, b)-distributed, with a being the 79 typed domestic cases with Pr e ⩾ 0·77 and b being the total number of typed domestic cases (n = 737) minus 79. Estimation was performed using @RISK (Palisade Corp., USA) by setting 10 000 iterations with the Latin Hypercube sampling technique and a seed of 1.
RESULTS
Genotypes
The 737 typed strains from non-travellers were assigned to 154 STs and 28 CCs, whereas those from the 232 travellers were assigned to 127 STs and 23 CCs. Twenty-eight STs from non-travellers and 23 STs from travellers were unassigned to previously identified CCs.
In non-travellers, the top five STs (ST53, ST50, ST21, ST48, ST45) accounted for >25% of cases and the top five CCs (CC21, CC45, CC206, CC257, CC48) for >50% of cases (Fig. 1). In travellers, the top five STs (ST572, ST21, ST50, ST53, ST353) accounted for ∼20% of cases and the top five CCs (CC21, CC353, CC828, CC206, CC52) for >50% of cases (Fig. 2). STs occurring once accounted for 46% and 70% of STs in non-travellers and travellers, respectively. PSI between travellers and non-travellers was 0·47 [95% confidence interval (CI) 0·34–0·59] while Simpson's index was 0·972 (95% CI 0·968–0·976) in non-travellers and 0·988 (95% CI 0·984–0·992) in travellers.

Fig. 1. Sequence types identified in Campylobacter jejuni/coli isolates from (a) 737 non-travellers (infections acquired in The Netherlands) and (b) 232 travellers returning to The Netherlands. Category ‘others’ includes sequence types that occurred less than five times (non-travellers) and less than twice (travellers).
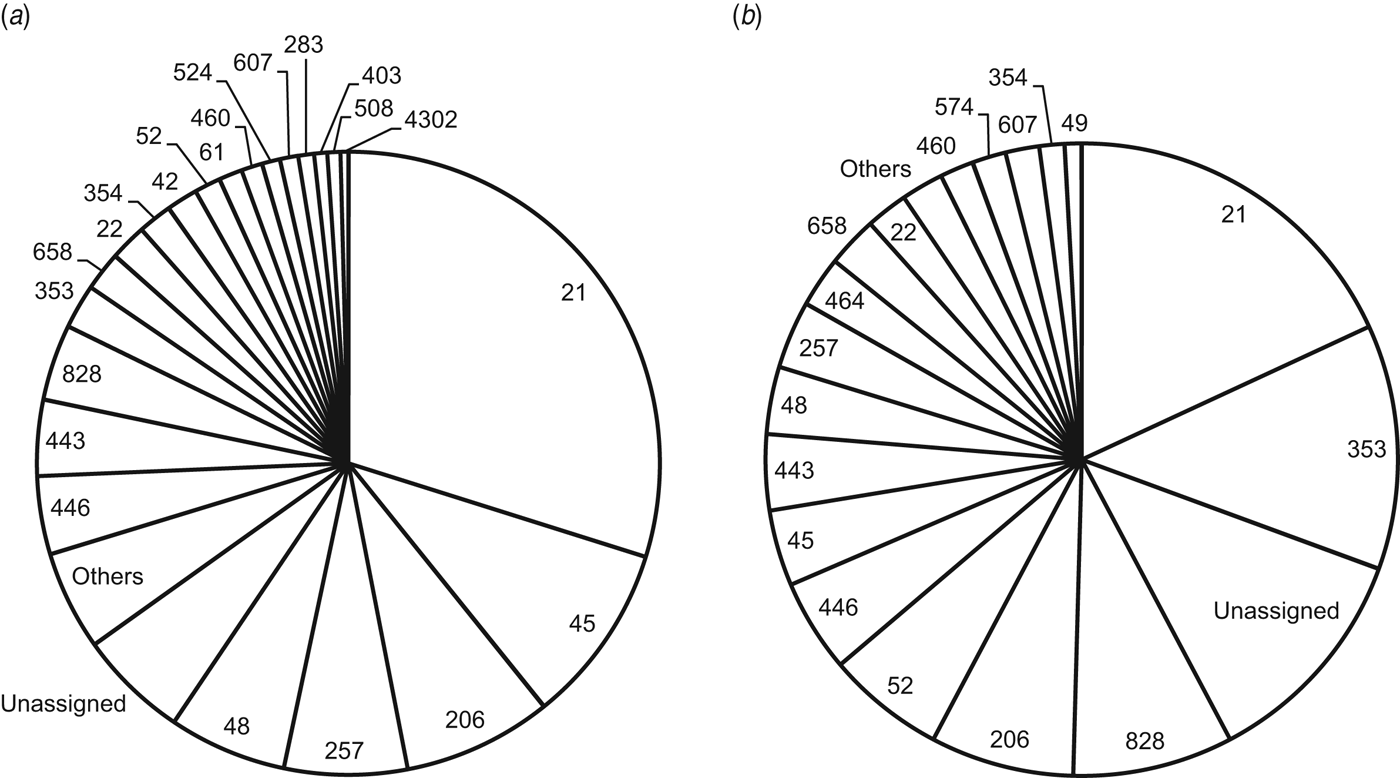
Fig. 2. Clonal complexes identified in Campylobacter jejuni/coli isolates from (a) 737 non-travellers (infections acquired in The Netherlands) and (b) 232 travellers returning to The Netherlands. Category ‘others’ includes clonal complexes that occurred less than twice (non-travellers) and less than once (travellers).
There were 68 STs (74 cases) found only in travellers and absent in any of the considered sources and in non-travellers (Table 2). Most (73%) cases infected with these traveller-only STs had travelled to Asia or Africa compared to 46% of all travel-related cases returning from these continents (z test, P < 0·001). Conversely, 23% of cases infected with traveller-only STs had travelled within Europe compared to 53% of all travel-related cases travelling within Europe (z test, P < 0·001).
Table 2. Sequence types found exclusively in travellers returning to The Netherlands but not in cases acquired in The Netherlands, nor in any of the sourced animal and environmental reservoirs

ST572 was significantly overrepresented (64%) in travellers from Western Europe (P = 0·001); ST50 in those from Western Asia (40%, P = 0·014); ST53 in those from Southern Europe (50%, P = 0·035); ST353, CC353 and CC828 in those from Northern Africa (38%, 27%, and 46%; P = 0·003, P < 0·001, and P < 0·001, respectively); CC52 in those from Eastern Europe (23%, P = 0·001).
STs with Pr e ⩾ 0·77 included in the risk factor analysis for exotically introduced domestic campylobacteriosis were ST1009, ST400, ST353, ST4307, ST5, ST2100, ST4308, ST2088, ST2151, ST1728, ST2844, ST508, ST403, ST443, ST350, ST3130, ST2156, ST878, ST657, ST4269, ST2882, ST2130, ST1080, ST4300, ST2187, ST586, ST696, ST587, ST2641, ST2553, ST3015, ST2123, ST4311, ST4278 and ST4284. There was a significant seasonal effect (P = 0·036) on Pr e, which peaked in October–December and decreased in April–June. No significant age and sex effects on Pr e were found.
Factors associated with travel-related campylobacteriosis
Compared to travelling within Western Europe, travelling to any region in Asia, Africa, Latin America and the Caribbean, and Southern Europe posed a higher risk of acquiring campylobacteriosis (Table 3), whereas the risk posed by Northern and Eastern Europe and Oceania, as well as the length of stay, were not significant (P > 0·05).
Table 3. Multivariable odds ratios (OR) and corresponding 95% confidence intervals (CI) of risk factors for Campylobacter jejuni/coli infection in travellers returning to The Netherlands

n.s., Not significant (P > 0·05).
a Adjusted for age, sex, degree of urbanization, season and level of education.
b Continuous variable, expressed as number of days stayed.
c Adapted from the United Nations scheme of the composition of macro-geographical (continental) regions, geographical sub-regions, and selected economic and other groupings. (http://unstats.un.org/unsd/methods/m49/m49regin.htm#europe).
d Includes travellers returning from Germany, France, Belgium, Austria, Luxemburg, and Switzerland.
e Includes travellers returning from the UK, Ireland, Denmark, Sweden, Norway, and Finland.
f Includes travellers returning from the Czech Republic, Hungary, Poland, Slovakia, and Romania.
g Includes travellers returning from Spain, Italy, Portugal, Greece, Croatia, and Malta.
h Includes travellers returning from Morocco, Egypt, and Tunisia.
i Includes travellers returning from Benin, Cameroon, Ethiopia, Ghana, Kenya, Mali, Nigeria, Rwanda, Tanzania, Botswana, Burkina Faso, South Africa and Namibia.
j Includes travellers returning from Turkey, Jordan and Iraq.
k Includes travellers returning from China, Indonesia, Malaysia, Singapore, Philippines, Thailand, and Vietnam.
l Includes travellers returning from India, Nepal, and Bangladesh.
m Includes travellers returning from Australia and Fiji Islands.
n Includes travellers returning from Bolivia, Ecuador, Peru, Venezuela, Chile, Costa Rica, Cuba, Dominican Republic, Haiti, Mexico, and Guatemala.
Factors positively associated with travel-related campylobacteriosis (Table 3) were: using proton-pump inhibitors, consuming vegetable salad when travelling outside Europe, handling and/or eating raw or undercooked pork, having chronic enteropathies, drinking bottled water when travelling to Southern Europe or non-European countries, and consuming chicken. Consuming yoghurt and being employed in healthcare were negatively associated with the outcome.
Factors associated with exotically introduced domestic campylobacteriosis
Factors positively associated with exotically introduced domestic campylobacteriosis (Table 4) were: not washing hands after toilet visit in January–April 2003, using proton-pump inhibitors, being a school-attending child in April–June 2002 and October–December 2002, attending public swimming pools in October–December 2002, and contact with gastroenteritis cases outside the household. Consuming yoghurt was negatively associated with the outcome. Sensitivity analysis of the cut-off Pr e revealed that this had no effect on the main results. In case-case analysis, the same factors were identified apart from consuming yoghurt and antacids (no longer significant), and therefore no further results are presented.
Table 4. Multivariable odds ratios (OR) and corresponding 95% confidence intervals (CI) of risk factors for Campylobacter jejuni/coli infection acquired in The Netherlands caused by strains of most likely exotic origin
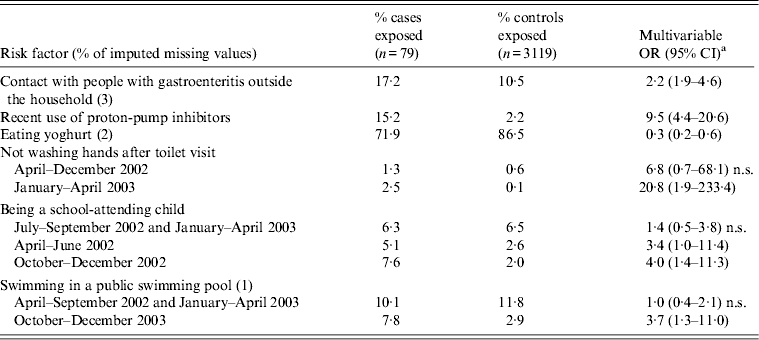
n.s., Not significant (P > 0·05).
a Also adjusted for age, sex, degree of urbanization, season and level of education.
It was estimated that, on average, 3·2% (95% CI 2·4–4·1) of all domestic cases are due to person-to-person transmission of exotically introduced STs.
DISCUSSION
The risk for Dutch travellers acquiring campylobacteriosis depended on the travel destination. Consistent with previous studies [Reference Ekdahl and Andersson2, Reference Hakanen3], Asia, Africa, Latin America and the Caribbean, and Southern Europe were regions associated with an increased risk for campylobacteriosis compared to Western Europe, which comprises the neighbouring countries of The Netherlands. Regional risk variations may be due to differences in local epidemiology and hygiene standards. However, a cautious interpretation of these figures is warranted, as physicians may be more likely to decide on laboratory investigation when a gastroenteritis patient has travelled to a high-risk destination [Reference Ekdahl and Andersson2]. Although it is also possible for regional risk variation to be substantial in domestic travel [Reference Strachan29], this may not apply to a relevant extent to The Netherlands, which is a relatively small country and is highly interconnected by high-speed routes and public transport, allowing for easy daily commute and travel, even from and to neighbouring countries such as Belgium and Germany, with very limited inbound overnight stay by the Dutch.
STs of travellers and non-travellers were relatively similar and travellers showed more ST diversity than non-travellers, a possible reflection of the numerous countries where they acquired infection. Moreover, some STs were more likely than others to infect travellers visiting specific regions, and travellers infected with STs that were undetected domestically had travelled predominantly to distant destinations in Asia and Africa, suggesting that differences in STs are related, to some extent, to the geographical distance of the travel region compared to The Netherlands, with STs from nearby European countries being generally more similar than those from farther destinations [Reference Smid13]. Another study [Reference Strachan29] showed nearly identical C. jejuni ST distributions in two English populations, but clear differences were found compared to Finland, Australia and New Zealand. Yet, differentiation was not proportional to distance, with the English sample being more similar to the samples from Australia and New Zealand than to the Finnish one, suggesting that similarities in STs between countries mainly reflect similarities of sources of infection rather than geographical proximity per se.
Consistent with [Reference McCarthy30], travel-related isolates had a significantly higher ST diversity than domestic isolates, with a different spectrum of MLST genotypes representative of the different countries. The larger ST diversity in travellers combined with the association of some STs with specific destinations is also consistent with the presence of heterogeneously distributed clones that are endemic in the different regions but not so prevalent elsewhere in the world. Regionally endemic STs have been identified, e.g. in Australia [Reference Mickan31], New Zealand [Reference McTavish32] and Curaçao [Reference Duim33], and may emerge because of clonal expansion, niche adaptation, geographical isolation and host immune selection [Reference Gupta and Maiden34]. Although so far there has been no evidence of ST-specific immune responses, it can be hypothesized that the chance of being exposed to a ST with uncommon antigens is somewhat higher for STs that are rarely, rather than commonly, encountered. STs that are associated with strong regional clustering would therefore pose a higher risk to travellers because of limited, prior (repeated) exposure in addition to issues related to sanitation failure. The risk posed by uncommon STs is also substantiated by their age distribution [Reference Mughini16, Reference Kärenlampi35]. For instance, the three most common STs in non-travellers were mainly found in the young relative to other STs, decreasing steadily with age (data not shown). Conversely, rare STs (<5 isolates) occurred independently of age. According to interpretation of similar findings [Reference Miller11], it is likely that antigenic properties associated with the common STs are frequently encountered throughout life; thus, the young would be more susceptible because they have encountered these STs less often. In contrast, rare STs, more probably related to uncommon antigens, would have rarely been encountered by all age groups.
The original CaSa study was designed to investigate factors associated with domestic campylobacteriosis and not purposely with travel-related Campylobacter infection [Reference Doorduyn6]. However, most risk exposures apply to some extent world-wide, and while the behaviour of travellers (and hence their exposure to campylobacters) may be different from the domestic population, it has been reported that most STs found in travellers returning to Scotland are still attributable to chicken in a similar proportion to that of domestic cases [Reference Strachan29]. Although this study did not have the power to conduct a stratified analysis by travel destination, it was possible to highlight some interactions between the exposures and the travel destinations.
Eight factors significantly associated with travel-related campylobacteriosis were identified. Consistent with evidence that poultry is the main reservoir for campylobacters, most studies concerning risk factors for campylobacteriosis have identified an association with eating chicken [Reference Doorduyn6, Reference Tam7, Reference Mughini16, Reference Evans36, Reference Kapperud37], suggesting that this factor is not exclusive for acquiring infection abroad. This also applies to consuming antacids and having chronic enteropathies [Reference Doorduyn6, Reference Tam7, Reference Mughini16].
In The Netherlands, <1% of domestically acquired Campylobacter infections have been attributed to pigs [Reference Mughini16]. Moreover, eating pork has been associated with a reduced risk for C. coli [Reference Doorduyn6] and chicken-borne C. jejuni/coli [Reference Mughini16] infections. Accordingly, Dutch retail pork has rarely been found contaminated with campylobacters [38]. The positive association with handling and/or eating raw or undercooked pork we found therefore suggests that pigs may be an important reservoir (and pork an important exposure) of campylobacteriosis outside The Netherlands.
In contrast to previous findings indicating that eating vegetable salad is negatively associated with the occurrence of (domestic) campylobacteriosis [Reference Doorduyn6, Reference Mughini16], we observed that this factor was associated with increased odds of acquiring campylobacteriosis when travelling outside Europe. In Europe, extensive sampling of raw vegetables, including ready-to-eat salads, has generally found no, or very few, campylobacters [38], suggesting that contamination of such items during irrigation, harvesting and processing is unlikely and that salads may occasionally become cross-contaminated during food preparation [Reference Evans36]. Conversely, exceptionally high Campylobacter isolation rates (∼68%) in raw vegetables were reported from countries such as Malaysia [Reference Chai39], indicating that major problems can arise by consuming vegetables if hygiene practices are absent or break down.
Drinking bottled water exclusively was associated with increased odds of acquiring campylobacteriosis when travelling to high-risk destinations. In the UK, drinking bottled water has been found to be positively associated with campylobacteriosis [Reference Evans36], particularly C. coli infection [Reference Gillespie40], and ciprofloxacin-resistant Campylobacter infection acquired abroad [Reference Evans5], suggesting that bottled water could, given the right circumstances, provide a vehicle for campylobacters [Reference Evans36]. In fact, bottled water, unlike tap water, is not usually treated and testing for Campylobacter is rarely undertaken [Reference Evans36, Reference Gillespie40]. Moreover, in the event of dual contamination of bottled water (campylobacters and organic matter), C. jejuni may survive for prolonged periods [Reference Tatchou-Nyamsi-König41]. However, our association with bottled water was only significant when travelling to high-risk destinations, supporting the hypothesis that drinking bottled water acts as a proxy for local circumstances where there is a generally high risk for campylobacteriosis. Travellers are indeed usually advised to drink bottled water where there is any doubt about the local water quality. The use of bottled water may help in preventing infection but there may be circumstances where the risk is higher than that which can be prevented by drinking bottled water. Moreover, our questionnaire did not distinguish between sparkling and still bottled water and did not ask whether it was consumed with or without ice. Therefore, further investigation is needed to assess if the advice of drinking bottled water merits any refinement.
Proportionally more controls than cases reported consuming yoghurt. It is believed that probiotic bacteria in yoghurt may alter the intestinal microflora in a way that prevents infection [Reference Kapperud37], while people working in healthcare might be particularly aware of the health risks (and ways to avoid them) when travelling.
Putative determinants of exotically introduced domestic campylobacteriosis were suggestive of anthroponotic transmission, namely contact with gastroenteritis cases outside the household (thus less likely to share the same exposure); not washing hands after toilet visit; being a school-attending child (usually having high frequencies of contacts); and attending public swimming pools (as recreational water has been proposed as a vehicle for Campylobacter transmission [Reference Mughini16, Reference Kapperud37]). Except for the first factor, the others were unidentified in previous analyses where the same cases were not split according to their estimated exoticism [Reference Doorduyn6, Reference Mughini16]. Moreover, we found significant interactions with season, which is in accordance with the seasonal nature of travelling, as also shown by the finding that Pr e varies seasonally. Periods most at risk were mainly those around popular holidays in The Netherlands, notably the autumn break in October, Christmas/New Year in December–January, and Easter in April–May. Moreover, people most at risk were school-attending children for which additional peaks in domestic campylobacteriosis have already been noted shortly after the end of school breaks, suggesting that these additional peaks are due to exposure to less common strains from less common foods consumed during the festivities and to the mixing of people that have not been in contact for a long time following on from the previous holidays [Reference van Pelt42]. It can therefore be hypothesized that travellers infected with strains possessing uncommon antigens might still be shedding them after returning home, most likely asymptomatically [Reference Havelaar14]. As there is unlikely to be a high prevalence of acquired protection against these exotic strains domestically, there is at least the potential for them to spread even through limited person-to-person transmission, with an average of 3·2% domestic cases being estimated to be due to anthroponotic transmission of exotically introduced strains. In conclusion, this study sought to provide an outline of Campylobacter transboundary epidemiology that may apply to other countries as well. This ranges from the identification of C. jejuni/coli MLST genotypes causing infection in international travellers to the risk factors potentially responsible for acquiring infection with such strains upon travelling, as well as those potentially responsible for their secondary spread to domestic populations. Besides the identification of high-risk travel destinations and universal correlates of campylobacteriosis, such as eating chicken, using antacids and having chronic enteropathies, this study also identified eating vegetable salad outside Europe, drinking bottled water in high-risk destinations and handling and/or eating raw or undercooked pork as factors associated specifically with travel-related campylobacteriosis. Moreover, suggestive evidence was provided that international travellers are particularly prone to infection with Campylobacter MLST genotypes that are endemic in the different travel regions, and that these exotic strains may be carried by the returning travellers and be spread to domestic populations through limited person-to-person transmission. As travellers have dynamic interactions with people, places, and microbes during their journeys, it is conceivable that they can be victims, carriers, and eventually transmitters of such agents to new regions and populations. Our understanding of campylobacteriosis may therefore depend on increased insight into regional risk differences, high-risk exposures and Campylobacter behaviour in response to newly available susceptible populations and changing environments.
ACKNOWLEDGEMENTS
We are grateful to M. A. S. de Wit and Y. Doorduyn for their contribution in designing the study; to P. Mullner for help with access to data; and to the laboratory staff of the Central Veterinary Institute, especially E. Pothoven, for taking care of the strain collection. We also thank the participating Regional Public Health Laboratories for their contribution to the data collection, especially F. Vlaspolder, J. H. Sloos, J. Spaargaren, J. Peereboom, M. A. Schouten, R. W. Brimicombe, F. W. Sebens, H. Rothbarth, L. J. M. Sabbe, H. Mulder, D. Veenendaal, E. Ijzerman, J. H. T. Wagenvoort, J. H. van Zeijl, B. M. de Jongh, M. Tersmette, P. Voorn, A. M. Horrevorts, J. Buitenwerf, B. G. A. Hendrickx, M. Peeters and A. R. Jansz. Thanks are also extended to H. C. Boshuizen for help in multiple imputation method and to M. L. Callegari for help in data entry.
DECLARATION OF INTEREST
None.






