


Introduction
Chinese privet (Ligustrum sinense Lour.) is an invasive shrub infesting roadsides, rights-of-way, and forested areas throughout the southeastern United States (Maddox et al. Reference Meloche and Murphy2010). It was introduced into the United States in 1852 and has since been widely used as a hedgerow and ornamental (Dirr Reference Dirr1990). Ligustrum sinense is a prolific fruit producer, and birds are an important mechanism of dispersal (Panetta Reference Siso and Burzycki2000; Swarbrick et al. Reference Young, Bell, Gross, Morrison and Haack-Gaynor1999). Once established, L. sinense also spreads asexually by lateral root sprouts and may form dense thickets. If left unchecked, L. sinense rapidly dominates forest understories by inhibiting woody native plant regeneration (Brantley Reference Brantley2008; Loewenstein and Loewenstein Reference Maddox, Byrd and Serviss2005) and shading out many herbaceous species (Merriam and Feil Reference Miller, Manning and Enloe2002). Ligustrum sinense also alters carbon and nitrogen dynamics (Mitchell et al. Reference Panetta2011). While the long-term dynamics of L. sinense invasion are still unclear, the loss of native woody and herbaceous species from heavily infested areas could result in a long-term shift from tree to shrub dominance (Hart and Holmes Reference Hart and Holmes2013).
Management Implications
Chinese privet (Ligustrum sinense Lour.) is one of the most invasive shrubs in the southeastern United States. Despite its abundance, there are few published studies evaluating the effectiveness of any control methods. We tested the cut stump treatment method with glyphosate and triclopyr at both spring and fall timings. We found both herbicides were extremely effective in controlling L. sinense at both timings at a lower concentration than generally used for cut stump treatment. However, we did observe a slight increase in L. sinense recovery from lateral root sprouts with the April timing compared with the November timing for triclopyr. Root collar diameter influenced treatment efficacy, with larger stems being more difficult to kill. These results provide land managers with multiple herbicides and application timings to achieve excellent L. sinense control.
To control L. sinense, many land managers have used physical methods, including mulching, shredding, and cutting, which provide immediate removal of the invaded shrub layer. However, these physical methods can be costly, labor-intensive (Smith et al. Reference Swarbrick, Timmins and Bullen1997), and ineffective for long-term control due to rapid stump and lateral root sprouting. Sprouting in woody plants, usually from suppressed buds immediately below the point of damage on the stem or roots, is typically a response to injury and results in the production of secondary trunks (Del Tredici Reference Del Tredici2001). Sprouting has been observed on many woody invasive plants, such as tree-of-heaven [Ailanthus altissima (Mill.) Swingle], for which cutting alone increased stand density, resulting in more than 12,355 stems ha−1 (Burch and Zedaker Reference Burch and Zedaker2003). Additionally, basal diameter can influence the number and vigor of the sprouts produced (Kays and Canham Reference Kays and Canham1991).
For species that can sprout, both the stems and roots must be killed for successful control (Burch and Zedaker Reference Burch and Zedaker2003). A year after cutting L. sinense without including an herbicide application, sprouts were large enough to be treated by a foliar spray (Harrington and Miller Reference Harrington and Miller2005). Cutting followed by an immediate application of herbicide to the stump (also known as cut stump treatment or a cut and treat application), can greatly reduce or prevent future sprouting in many woody and invasive species, including A. altissima, Chinese tallowtree [Triadica sebifera (L.) Small], black locust (Robinia pseudoacacia L.), hardy orange [Poncirus trifoliata (L.) Raf.], honey locust (Gleditsia triacanthos L.), shoebutton ardisia (Ardisia elliptica Thunb.), and many others (Burch and Zedaker Reference Burch and Zedaker2003; DiTomaso and Kyser Reference DiTomaso and Kyser2007; Enloe et al. Reference Enloe, O’Sullivan, Loewenstein, Brantley and Lauer2016; Harmoney Reference Harmoney2016; Hartman and McCarthy Reference Hartman and McCarthy2004; Meloche and Murphy Reference Merriam and Feil2006; Miller et al. Reference Miller, Manning and Enloe2013; Siso and Burzycki Reference Smith, Larson, Kelty and Ashton2004; Young et al. Reference Zedaker, Lewis, Smith and Kreh2017).
Two herbicides widely used for cut stump treatment are glyphosate and triclopyr amine. Both are generally applied to the cut surface of target woody plants at a 50% to 100% concentration in water. However, anecdotal evidence suggests lower concentrations may be useful for certain species, but this has not been tested for L. sinense (Miller et al. Reference Miller, Manning and Enloe2013).
The effectiveness of these two herbicides may also vary among treated species. For L. sinense, Harrington and Miller (Reference Harrington and Miller2005) found that glyphosate was more effective than triclopyr when applied as a foliar spray. This may indicate differential sensitivity for cut stump applications as well. Additionally, the seasonal timing of cut stump treatments can strongly influence subsequent sprouting (Kays and Canham Reference Kays and Canham1991). Delanoy and Archibold (Reference Delanoy and Archibold2007) stated that the best time to treat European buckthorn (Rhamnus cathartica L.) was in the fall, while Evans et al. (Reference Evans, Moorhead, Bargeron and Douce2006) recommended fall application and avoiding cut stump treatments with triclopyr in the spring. Given these questions, the objective of this study was to compare the efficacy of triclopyr and glyphosate cut stump treatments at spring and fall timings. Additionally, we wanted to determine whether L. sinense stem size influences herbicide efficacy for glyphosate and triclopyr cut stump treatments. Answering these questions would greatly assist land managers in improving L. sinense control with cut stump herbicide treatments.
Materials and Methods
A study was conducted from 2008 to 2010 at two forested sites on the Auburn University campus in Auburn, AL. Auburn is within USDA plant hardiness zone 8a and is characterized by mild winters and hot summers with a mean annual precipitation of 1,360 mm. When compared with the historic average, mean monthly cumulative precipitation was generally lower for the 2008 to 2010 study period, especially over the summer months of June and July (Figure 1A). Monthly minimum and mean temperatures over the 3-yr study period were slightly higher than the long-term averages, while monthly mean maximum temperatures were slightly lower in the fall and winter (Figure 1B). This is generally reflective of slightly warmer and drier conditions than the historic averages.
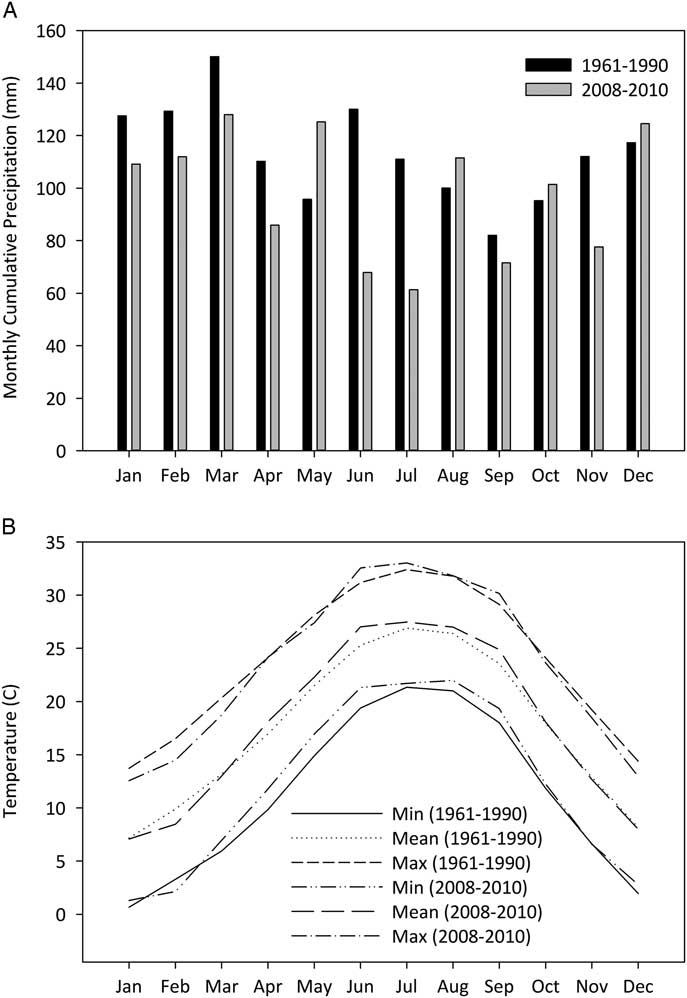
Figure 1 Mean monthly cumulative precipitation (A) and temperature trends (B) for the study period (2008–2010) compared with the long-term (1961–1990) trends.
The first study site was a 1-ha upland hardwood forest (32.5837°N, 85.5035°W) predominantly composed of L. sinense shrubs approximately 6-m tall. Native species scattered throughout the site included sugarberry (Celtis laevigata Willd.), mockernut hickory [Carya tomentosa (Lam.) Nutt], and blackjack oak (Quercus marilandica Münchh.). Other nonnative species occurring in the study area included sacred-bamboo (Nandina domestica Thunb.), chinaberry (Melia azedarach L.), and T. sebifera. Soils were a mixture of Pacolet sandy loam and Marvyn loamy sand. The second study site was a 1-ha bottomland forest (32.5961°N, 85.4956°W) with understory and midstory completely dominated by L. sinense approximately 6 m in height. The overstory was composed of water oak (Quercus nigra L.), loblolly pine (Pinus taeda L.), sweetgum (Liquidambar styraciflua L.), and eastern redcedar (Juniperus virginiana L.). Silktree (Albizia julibrissin Durazz.), M. azedarach, kudzu [Pueraria montana (Lour.) Merr. var. lobata (Willd.) Maesen & S.M. Almeida ex Sanjappa & Predeep], N. domestica, and English ivy (Hedera helix L.) were scattered throughout the site. The soil was a Kinston loam.
Individual L. sinense shrubs or small trees across a range of sizes from approximately 2 to 30 cm (measured by root collar diameter) served as experimental units in a completely randomized experimental design. This size range was slightly larger than reported for operational cut stump treatment of L. sinense from Arkansas (Young et al. Reference Zedaker, Lewis, Smith and Kreh2017) and consistent with other forests sampled in Alabama (Hart and Holmes Reference Hart and Holmes2013). Three treatments were applied at two seasonal timings at each of the two sites, and each treatment was replicated on 50 individual shrubs for a total of 600 experimental units. Spring treatments were applied between April 10 and 16, 2008, at both sites. Fall treatments were applied between November 18 and 21, 2008. Before the initiation of treatments, the root collar diameter of each shrub was measured with wooden calipers and recorded. Treatments consisted of glyphosate (Accord® Concentrate, 480 gL−1, Dow AgroSciences, Indianapolis, IN) applied as a 25% v/v solution (120 gL−1), triclopyr amine (Garlon® 3A, 360 gL−1, Dow AgroSciences) applied as a 25% v/v solution (90 g L−1), and an untreated control. Both herbicides were mixed with water and a nonionic surfactant (Activator 90, Loveland Products, Loveland, CO) at 0.5% v/v.
At each application timing, L. sinense shrubs were cut with a chainsaw (Husqvarna 123HD60, Husqvarna, Charlotte, NC 28269), leaving a flat, 2.5-cm stump. Any accumulated sawdust or other debris was quickly removed, and herbicide treatments were applied within 30 s of cutting. Treatments were applied with an Echo MS-4 backpack sprayer (Echo Incorporated, Lake Zurich, IL) with a single adjustable cone nozzle for the April timing and a 1.5-L pressurized Garden Plus hand sprayer (LG Sourcing, North Wilkesboro, NC) with a single adjustable cone nozzle for the November timing. To ensure adequate coverage, the entire cut surface of each stump was sprayed to wet but not to the point of runoff. The amount of herbicide applied varied with the size of each stump.
Data were collected at 6, 12, and 18 mo after treatment (MAT) for each treatment and application timing. Total number of stump sprouts originating from the stump and all sprouts initiating from lateral roots within a 30-cm radius of each stump were recorded. Additionally, the height of all stump and lateral root sprouts was measured at each 6-mo time point to assess regrowth vigor. Sprouts were separated by origination point (i.e., from the stump or lateral roots) to better quantify the impact of the treatments. Lateral root sprouts were only quantified within a 30-cm radius of each stump due to difficulty in ascertaining the origin of sprouts beyond that distance. The origin of sprouts from lateral roots within the 30-cm radius was easy to discern, as laterals were generally exposed or just below the soil surface for that distance. Additionally, L. sinense seedlings were hand pulled within the 30-cm radius around each stump at each sample date to prevent confusion with lateral root sprouts. Ligustrum sinense seedlings were very easy to distinguish from root sprouts due to the presence of cotyledons and a distinct taproot.
Statistical Methods
Analyses of rootstock mortality, number of either total sprouts (stump sprouts+lateral sprouts) or lateral sprouts per rootstock, and the sum of corresponding sprout heights per rootstock were performed as generalized linear models using PROC GLIMMIX in SAS (Littell et al. Reference Littell, Milliken, Stroup, Wolfinger and Schabenberger2006). The analysis of rootstock mortality treated mortality as a binary outcome using a logit-link function to estimate the probability that a rootstock was killed. The number of sprouts and the sum of sprout heights were considered Poisson-distributed random variables using the natural log of the rootstock sample size (the total of live and dead rootstocks) as an offset. Site, treatment, and timing were considered fixed effects. The site by treatment by timing interaction was considered a random effect in this design. Evaluation date (i.e., 6, 12, or 18 MAT) was considered a nested (subplot) effect within site by treatment by timing combinations for the initial analysis of mortality. The main effects (site, treatment, and treatment timing) error term was the pooled variation of these main effect interactions. This pooling was done after separate runs of the model found that the site by treatment (p=0.135) and site by timing (P=0.489) interactions were not significant.
Following the initial analysis of mortality, subsequent analyses focused on long-term control at 18 MAT and did not include MAT as a factor. Analysis of covariance was used to relate mortality rate at 18 MAT to rootstock diameter (measured before treatment). Mean comparisons were performed using the Tukey-Kramer adjustment for multiplicity and a significance level of P=0.05.
Results and Discussion
The initial analysis of rootstock mortality over time found no influence of evaluation date (6, 12, or 18 MAT) on the outcome (P=0.41). Additionally, for the initial analysis, there were no treatment, timing, or site interactions with evaluation date (Table 1). The lack of significant interactions with evaluation date indicated that most mortality likely occurred by the time of the first evaluation at 6 MAT. This is noteworthy, because woody plant mortality following herbicide treatment often occurs one to two full growing seasons after treatment (Zedaker et al. Reference Zedaker, Lewis, Smith and Kreh1987). However, for L. sinense, this is more likely an indication that any sprouting following cutting or cut stump treatment will generally occur within 6 mo, regardless of spring or fall cutting.
Table 1 ANOVA for mortality assessed at 6, 12, and 18 mo after treatment (MAT) and long-term control at 18 MAT for several variables. Significant values (P<0.05) are bolded for clarity.Footnote a
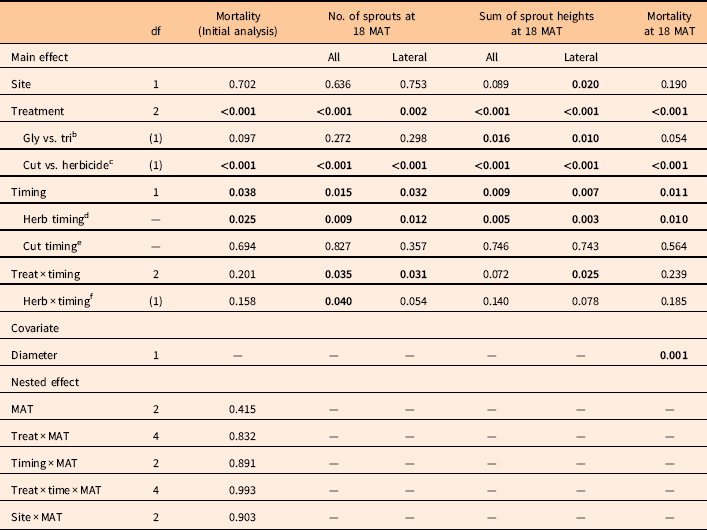
a Analysis of long-term control included mortality, average number of sprouts (total and lateral) per rootstock, and average sum of sprout heights (total and lateral) per rootstock. Analysis of mortality at 18 MAT included rootstock diameter as a covariate.
b Comparison of glyphosate to triclopyr treatments.
c Comparison of cut stump treatments to averaged herbicide treatments.
d Comparison of April vs. November timing for cut + herbicide treatments.
e Comparison of April vs. November timing for cut-only treatments.
f Interaction of herbicide treatments and timing (excluding cut-only treatments).
The initial analysis of mortality indicated treatment and application timing were significant. For treatment, these were driven by the difference between cutting alone versus cutting plus herbicide (P<0.001) (Table 1). For application timing, these differences were driven by herbicide application timing (P=0.025), as cutting alone by timing was not significant (P=0.694). This initial analysis of mortality indicated that significant effects were strongly herbicide driven, which is what would be expected.
In the subsequent analysis, the relationship between rootstock diameter and 18 MAT mortality rate was examined in a sequential fashion. The mortality rate is the predicted probability of mortality from the analysis of covariance given the treatment used and initial rootstock diameter. Mortality “rate” is not directly observable, because a rootstock is either live or dead, but the probability of mortality can be related to rootstock diameter using an analysis of covariance. A complication here was that reasonably good control from herbicide treatments limited the sample size of live rootstocks and limited the ability to obtain good estimates of the relationship between mortality rate and diameter. With this in mind, the first tests addressed whether rootstock diameter was a significant covariate (P<0.001) and whether slopes differed by treatment and timing (P=0.482). A rootstock size classification was then created for rootstocks less than 5.0 cm in diameter and those greater than 5.0 cm (53% of observations were in the smaller class), and this effect was incorporated into the analysis. Mortality was significantly different between the two diameter classes (P=0.021), but there were no significant interactions between diameter class and treatment by timing combinations (P=0.6). This process validated the analysis of covariance results, in that survivors (even when sparse) were mostly in the larger-diameter class regardless of treatment and timing. Finally, the covariance model using a common slope parameter for diameter was adopted to describe the relationship between mortality rate and rootstock diameter. Local area nonparametric regression plots of mortality with initial rootstock diameter (PROC SGPLOT with the LOESS statement) confirmed that the concave shape for the untreated cut stump treatments and the convex shape for herbicide treatments were consistent with the data.
The analysis of covariance for 18 MAT mortality (Table 1) tested for treatment differences at average rootstock diameter. There were significant differences due to treatment and timing, but these differences were between herbicide and untreated control treatments as well as due to application timing across herbicide treatments (Table 1). These findings are very similar to the initial mortality analysis that encompassed all sample dates. Differences in mortality at 18 MAT between triclopyr and glyphosate treatments were not significant at the 5% level (P=0.054), as they resulted in 89% and 94% mortality, respectively (Table 2). Mortality for the cutting treatment with no herbicide resulted in 14% mortality. The reason for this level of mortality is unclear but has been observed with other prolific sprouters, including A. altissima (Burch and Zedaker Reference Burch and Zedaker2003). Future research to elucidate why some shrubs died following cutting could be extremely beneficial to land managers, as a 15% reduction in application costs could be quite valuable.
Table 2 Probability of mortality at 18 months after treatment estimated using rootstock diameter as a covariate.
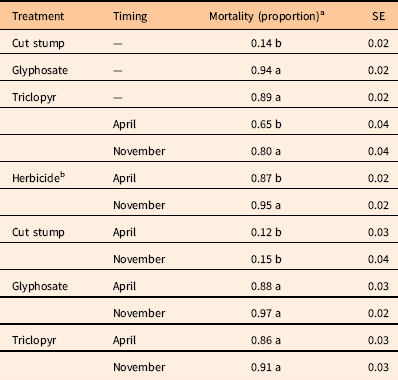
a Means within main effects (treatment or timing) or between interactions (treatment by timing) followed by the same letter are not different (P=0.05).
b April and November timing effects for herbicide applications (excluding the cut treatment).
When pooled across all three treatments, mortality was greater with the November timing (80%) compared with the April timing (65%). This generally supports the concept that woody plant control is better with fall than spring treatments in the southeastern United States. This also was the case for the herbicide treatments, as mortality differed by timing when the untreated controls were excluded. The November herbicide timing resulted in 95% mortality, while the April herbicide timing resulted in 87% mortality (Table 2). However, specific comparisons of timing within treatments did not clarify any key drivers for this, as there were no differences between timings within each treatment (Table 2).
Predicted probability of mortality using the covariance model pooled across sites (Figure 2) shows the relationship between mortality and root collar diameter for treatment and timing combinations. Both herbicide treatments are included to show the distribution of rootstock size for each treatment, even though there were no significant differences between triclopyr and glyphosate (Table 1). The finding that mortality decreases with increasing rootstock diameter, particularly for the larger-sized rootstocks, was a somewhat unexpected result, as cut stump treatments are recommended across a wide range of tree sizes (Miller et al. Reference Miller, Manning and Enloe2013). The explanation of this outcome is not clear. However, it may possibly be due to the multistemmed nature of L. sinense, in which bark inclusions are likely common. This bark growth could possibly limit herbicide translocation if the height of stump cutting occurs where there are bark inclusions. We did not quantify this occurrence in our study, but regard it as possible, because many experimental units had multiple stems that originated below our cutting height.
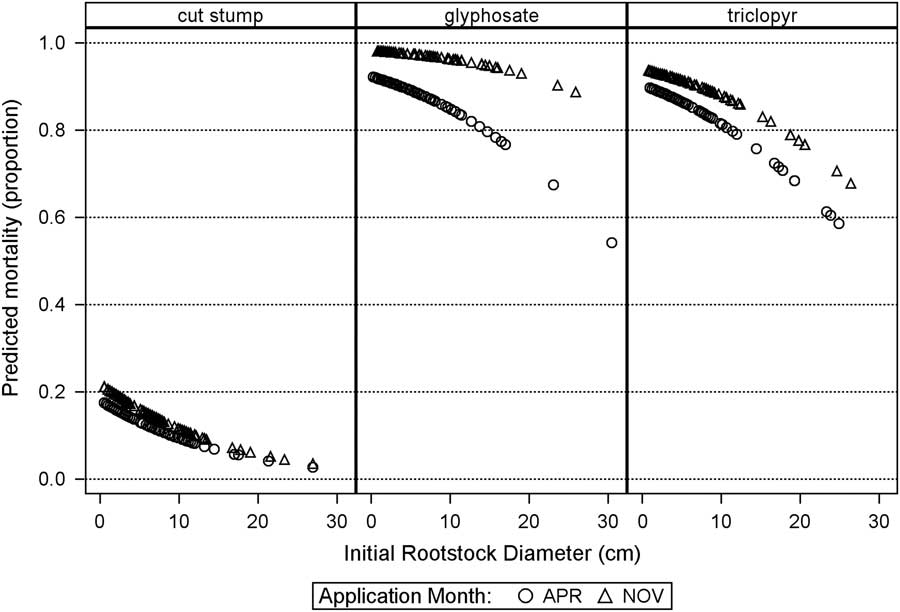
Figure 2 The relationship between predicted probability of mortality 18 mo after treatment and initial (baseline) rootstock diameter from the analysis of covariance model. Each symbol represents a rootstock observation.
The ANOVA results for number of sprouts at 18 MAT were similar to those for mortality (Table 1). The average numbers of total sprouts and lateral sprouts per treated rootstock were substantially reduced by both herbicide treatments compared with cutting alone (Table 3). However, there were no differences between the two herbicides in total sprouting or lateral sprouting. It is noteworthy that there were almost no stump sprouts found on stumps treated with either herbicide, and lateral sprouts were the dominant component of total sprouting. This has been commonly observed in other aggressive root-sprouting species such as A. altissima and T. sebifera. This was not the case for the control treatments, in which most sprouts originated from the stumps (Table 3). Sprout number was lower for November compared with April across all three treatments. However, there were no differences between herbicides within timings for either sprouting variable (Table 3).
Table 3 Mean number of sprouts per treated rootstock at 18 months after treatment.Footnote a

a Means within main effects (treatment or timing) or between interactions (treatment by timing) followed by the same letter are not different (P=0.05).
b April and November timing effects for herbicide applications (excluding the cut treatment).
These results carried over to the sum of sprout heights at 18 MAT, except that there were significant differences between glyphosate and triclopyr treatments (Table 1). Cumulative sprout heights are an indication of the vigor of the regrowth that occurred following treatment. Glyphosate reduced total and lateral sums of sprout heights more than triclopyr (Table 4). Most of this difference was from lateral sprouting, as both herbicides almost completely prevented stump sprouts. This differential inhibition of lateral root sprouts has been observed in silvicultural studies, which suggest glyphosate is more effective than triclopyr for controlling species prone to lateral root sprouting (Kochenderfer et al. Reference Kochenderfer, Kochenderfer and Miller2012). Application timing also was significant, as the November timing resulted in significantly lower sprout heights compared with the April timing, and this difference was almost exclusively from lateral root sprouts. Control of many woody plants is commonly suggested to be better when translocation patterns are reflective of a downward flow of photosynthates to the roots in the fall for winter storage.
Table 4 Sum of sprout heights per treated rootstock at 18 months after treatment.Footnote a
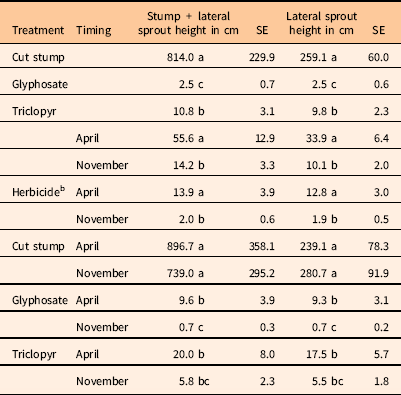
a Means within main effects (treatment or timing) or between interactions (treatment by timing) followed by the same letter are not different (P=0.05).
b April and November timing effects for herbicide applications (excluding the cut treatment).
These results indicate that cut stump treatments of both glyphosate and triclopyr amine are highly effective in controlling L. sinense. The fall treatment timing was significantly better than the spring timing which is in general agreement with other timing studies. Enloe et al. (Reference Enloe, O’Sullivan, Loewenstein, Brantley and Lauer2016) found basal bark treatment of L. sinense with triclopyr ester in the spring was not as effective as in the winter. In other studies, Delanoy and Archibold (Reference Delanoy and Archibold2007) found a fall cut stump application for R. cathartica was an effective control method, but Boudreau and Willson (Reference Boudreau and Willson1992) showed that both fall and summer timings were equally effective for R. cathartica, resulting in 100% mortality. Kochenderfer et al. (Reference Kochenderfer, Kochenderfer and Miller2006) suggested that the best time to apply glyphosate is when the plant is actively growing but that control of root sprouts can also achieved if herbicide is applied during a time of no growth. However, Kochenderfer et al. (Reference Kochenderfer, Kochenderfer and Miller2006) also noted that herbicide efficacy for controlling American beech (Fagus grandifolia Ehrh.) root sprouts is less consistent when herbicide is applied to stumps in the spring.
These results also show that lower than labeled concentrations of both herbicides are very effective for controlling L. sinense. Both herbicide labels for the products used in this study recommend a 50% to 100% v/v concentration. We found a 25% v/v concentration was very effective for both, which would reduce herbicide inputs into the environment by 50% to 75%. Reduced herbicide use is highly desirable goal of many land managers, and our work strongly supports this goal. These improved results are not always possible with all woody invasive plants. For example, Enloe et al. (Reference Enloe, Loewenstein, Streett and Lauer2015) found that triclopyr amine applied at a 25%v/v concentration was not at all effective for T. sebifera control. Future work on optimizing herbicide dose should focus on concentration and application rate determined by stem size and expanded testing of other active ingredients, with superior matches leading to even further reductions.
Acknowledgments
The authors would like to thank Brian Anderson for coordinating use of the one of the sites on the Auburn Swine Unit and the Alabama IPM Mini-grant Program for funding this research. No conflicts of interest have been declared.






