



Management Implications
Management efforts are logistically and financially time-consuming. Treatments that allow weed managers to “work smarter, not harder” are clearly advantageous. In general, it is well worth being very clear about the management objective; specifically, are management efforts to be assessed at shorter- or longer-term timescales, and are density, growth or spread outcomes of most concern. For example, management efforts required to reduce abundance or growth of a broadly distributed weed species (such as mowing or herbicide application) may be very different than the interventions required to reduce spread and infilling (such as control of seed dispersal or transport) by a new incursion of the same species. Failing to be specific about objectives may thus lead to confusion or conflict if managers work to achieve different implicit aims. In the case of the invasive thistle Carduus nutans (musk thistle), the stated objective does indeed affect the chosen management intervention. We assessed three potential management objectives (reducing existing population density; decreasing long-term population growth; and limiting population spread) using 14 possible mowing treatments (in terms of intensity, frequency, and timing, and thus logistical effort) in a two-cohort field study of C. nutans. The most effective intervention depends on the desired outcome. However, importantly, for all objectives assessed, there is a consistent conclusion that fewer, well-timed mowing efforts are more effective than standard intervention recommendations to mow as often as possible. Thus, managers may be able to improve management results both in terms of effort and in terms of the directly measured outcome of management.
Introduction
Successful weed management depends on a clear statement of management goals (Shea et al. Reference Shea, Jongejans, Skarpaas, Kelly and Sheppard2010). Many managers discuss “weed control” or “weed management,” but these broad terms necessarily encompass a wide array of more precise objectives. Possible specific management goals with regard to invasive weeds include: to prevent arrival in the first place, to eliminate an existing population, to eradicate all populations in a geographic area, to lower plant population density to tolerable levels (i.e., below some economic or environmental damage threshold), to prevent or reduce weed reproduction, to contain the weed within a specific area, to reduce weed emergence after crop planting, to minimize crop damage, to minimize economic losses, to reduce impacts, to shift weed community composition to easier-to-manage species, or to reduce population growth or population spread of the targeted weed (Buhler Reference Buhler2002; Liebman Reference Liebman, Liebman, Mohler and Staver2001; Mangold et al. Reference Mangold, Fuller, Davis and Rinella2018; Mortensen et al. Reference Mortensen, Rauschert, Nord and Jones2009; Shea and Kelly Reference Shea and Kelly1998; Shea et al. Reference Shea, Jongejans, Skarpaas, Kelly and Sheppard2010; Skurski et al. Reference Skurski, Rew and Maxwell2014; Wilkerson et al. Reference Wilkerson, Wiles and Bennett2002). These management goals can differ in both their time dimensions and spatial scales. For example, some focus on short-term control outcomes such as immediate reductions in the density of existing populations, while others are aimed at long-term effects and are usually based on population projections such as growth rate. Although management should ideally focus on long-term outcomes rather than a single season or year (Jones and Medd Reference Jones and Medd2000), management decisions are often constrained by the fact that managers typically face economic challenges and act on short time horizons (Hyder et al. Reference Hyder, Leung and Miao2008). Furthermore, management goals can also be distinguished by the spatial scales on which they are defined. For instance, some may focus on localized population spread, which is dominated by habitat-infilling processes due to local dispersal. Others may focus more on the regional spread of the species, which includes establishment of new populations as a result of human-mediated long-distance dispersal.
Identifying the management goal is critical (Milner-Gulland and Shea Reference Milner-Gulland and Shea2017; Probert et al. Reference Probert, Shea, Fonnesbeck, Runge, Carpenter, Dürr, Garner, Harvey, Stevenson, Webb, Werkman, Tildesley and Ferrari2016; Shea et al. Reference Shea, Jongejans, Skarpaas, Kelly and Sheppard2010). Because specific objectives are evaluated based on different life-history traits or their resulting population outcomes, the best strategy to achieve an objective may vary. For example, herbicide application may be an effective approach to achieve short-term management goals but may ultimately lead to evolution of resistance (Shaner Reference Shaner2000) and off-target effects (Crone et al. Reference Crone, Marler and Pearson2009). In contrast, release of biocontrol agents (available for only a small proportion of invasive species) often involves time lags but is more effective in the long run (Hyder et al. Reference Hyder, Leung and Miao2008). In this case, having a clearly stated time horizon is a prerequisite for making management decisions. Outcomes may vary dramatically based on exactly how management treatments are applied (Rinella and Hileman Reference Rinella and Hileman2009).
Besides different time horizons, management objectives, and hence their corresponding optimal strategies, can also vary depending on population spatial structure (Thomas and Kunin Reference Thomas and Kunin1999). While many previous studies have focused on reducing local population abundance, only recently has much attention been given to population spread rates in invasion control modeling (Bogich et al. Reference Bogich, Liebhold and Shea2008; Dauer et al. Reference Dauer, Luschei and Mortensen2009; Epanchin-Niell and Wilen Reference Epanchin-Niell and Wilen2012; Marchetto et al. Reference Marchetto, Shea, Kelly, Groenteman, Sezen and Jongejans2014; Moody and Mack Reference Moody and Mack1988; Neubert and Parker Reference Neubert and Parker2004; Shea et al. Reference Shea, Jongejans, Skarpaas, Kelly and Sheppard2010), in part because of recently developed modeling methodologies for coupling structured demographic and dispersal processes (Jongejans et al. Reference Jongejans, Shea, Skarpaas, Kelly, Sheppard and Woodburn2008, Reference Jongejans, Shea, Skarpaas, Kelly and Ellner2011; Neubert and Caswell Reference Neubert and Caswell2000; Neubert and Parker Reference Neubert and Parker2004). Consequently, optimal management strategies for controlling local population density and for controlling population spread may not be the same. For example, in New Zealand, the biocontrol agent Rhinocyllus conicus (Froelich, Coleoptera: Curculionidae) (which attacks the thistle receptacle) had a larger effect on the rate of increase of local population size of musk thistle (Carduus nutans L., Asteraceae) than another agent, Trichosirocalus horridus (Panzer, Coleoptera: Curculionidae) (which attacks the plants’ roots), whereas the reverse was true for controlling the spatial spread of the population (Shea et al. Reference Shea, Jongejans, Skarpaas, Kelly and Sheppard2010).
In this study, we address three potential management objectives for the nonnative, invasive thistle Carduus nutans, a monocarpic perennial weed that is native to Eurasia but has become invasive in many other regions in the world (Allen and Shea Reference Allen and Shea2006). Infestations by this species cause significant loss to pastures and rangelands, as the invader prevents livestock from grazing nearby and reduces productivity (Desrochers et al. Reference Desrochers, Bain and Warwick1988). Carduus nutans flowering occurs from May to August (late spring to late summer) and is followed by a large production of wind-dispersed seeds (Rhoads and Block Reference Rhoads and Block2000). Mowing is a commonly applied management tool to control invasive weeds (DiTomaso Reference DiTomaso2000) and has been widely used to manage infestations caused by C. nutans (McCarty and Hatting Reference McCarty and Hatting1975; Popay and Medd Reference Popay and Medd1990; Tipping Reference Tipping2008). However, comparisons of management outcomes are often limited to only a few mowing regimes, and the evaluations of these regimes are mostly based on reductions in plant density or seed production. Therefore, an integrated evaluation is useful to assess a spectrum of treatments (differing in management timing, intensity, and frequency) in the context of these different possible management goals.
We investigated the effects of 14 different mowing regimes to control C. nutans. A previous two-cohort empirical study by Zhang and Shea (Reference Zhang and Shea2012) tested these regimes in terms of intensity (i.e., mow at 5 cm vs. 20 cm), frequency (i.e., mow once, twice, or three times within a growing season), and timing (i.e., mow before flowering, during early flowering, or at flowering peak). Zhang and Shea (Reference Zhang and Shea2012) found that, while many mowing studies focus on high-frequency and high-intensity mowing, once timing of key life-history or management events is also considered, frequency may be less relevant than it first appears; high frequency appears as statistically significant because it is more likely than low frequency to catch a key timing. The effect of these 14 mowing regimes on the survival, growth, and reproduction of C. nutans differed (Zhang and Shea Reference Zhang and Shea2012). We use these empirical data, combined with other demographic and dispersal data for this species, to develop models and compare the mowing regimes based on their effectiveness in terms of realizing each of the following management goals:
1. to reduce population density of the current cohort;
2. to reduce projected local population growth rate, λ; and
3. to reduce projected population spatial spread rate, c* (which integrates both population growth and propagule dispersal; Neubert and Caswell Reference Neubert and Caswell2000).
Given the differing effects of these management strategies on different vital rates (Zhang and Shea Reference Zhang and Shea2012), we hypothesize that the most effective management strategies for the three management goals should differ.
Materials and Methods
Fourteen mowing treatments (with 12 replicates each) and a control treatment (no mowing, with 24 replicates to provide a solid baseline) from each of two cohorts were included in this study, using data from the experimental study by Zhang and Shea (Reference Zhang and Shea2012). Mowing treatments refer to the combination of one, two, or three separate empirical mowing events (Table 1). Empirical survival of adult plants was used to evaluate management effectiveness in terms of reducing density of the existing population (Objective 1). Evaluation of reductions in projected population growth rate (Objective 2) was based on a population matrix model with modified demographic vital rates. Reductions in projected population spread (Objective 3) were assessed using a spatial matrix model that incorporates modifications on both demographic vital rates and an important dispersal-related trait, plant height (Skarpaas and Shea Reference Skarpaas and Shea2007). Details of the demographic, dispersal, and spread models are available in the Supplementary Material, but are summarized below.
Table 1. Summary of effects of different empirical mowing treatments on plant survival, lifetime capitulum production, and plant height from Zhang and Shea (Reference Zhang and Shea2012).a
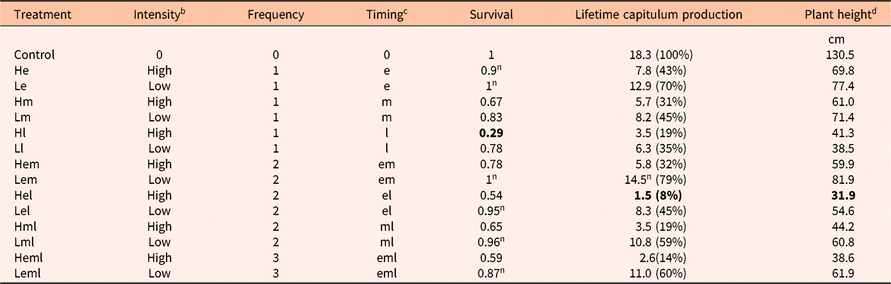
a Zhang and Shea (Reference Zhang and Shea2012) data were used to parameterize the models for each of the mowing treatments. Note that a small subset of these data were presented in Figure 1 in Zhang and Shea (Reference Zhang and Shea2012). Bold numbers denote the lowest values for the 14 mowing regimes. The superscript “n” denotes results that are not significantly different from the control (P > 0.05). For survival, treatments He, Le, Lem, Lel, Lml, and Leml were not different from the control. For lifetime capitulum production, only treatment Lem was not different from the control. Numbers in parentheses denote percentage modifications of reproduction used in the models.
b high, cutting at 5 cm aboveground; low, cutting at 20 cm aboveground.
c e, early cut (immediately after individual plants reached 40 cm in height); m, middle cut (2 wk after plants reached 40 cm); l, late cut (4 wk after plants reached 40 cm).
d Plant height in all mowing treatments is lower than in the control.
Plant survival was defined as the probability of surviving to produce viable seeds in the same season. Plant height was defined as height of flowering plants at the end of the growing season. Plant lifetime capitulum production comprises two parts of reproduction: (1) capitula produced before the initiation of the mowing treatments (i.e., pre-mow reproduction); and (2) capitula produced after the completion of the mowing treatments (i.e., post-mow reproduction). No mature capitula developed between any two mowing events in any of the mowing treatments.
Plant survival and capitulum production of surviving plants (Table 1) were incorporated into a 4 by 4 size-structured baseline demographic matrix model. This model has four stages: seedbank, small rosettes, medium rosettes, and large rosettes. Baseline model parameters were derived from a previous field experiment conducted at the same field site (the Pennsylvania experimental population in Jongejans et al. [Reference Jongejans, Shea, Skarpaas, Kelly, Sheppard and Woodburn2008]). We then modified reproduction based on significant percentage changes caused by the mowing treatments in the present study. In these models, we assumed that percentage decreases in reproduction caused by mowing did not depend on the size of the plants, and seed production per capitulum, as well as other vital rates, remained unchanged. Population growth rates λ were calculated as the dominant eigenvalues of the demographic matrices.
The demographic matrices were then coupled with the Wald analytical long-distance dispersal model (Katul et al. Reference Katul, Porporato, Nathan, Siqueira, Soons, Poggi, Horn and Levin2005) to project population spread rates, c*, using integrodifference equations (Jongejans et al. Reference Jongejans, Shea, Skarpaas, Kelly, Sheppard and Woodburn2008; Neubert and Caswell Reference Neubert and Caswell2000; Shea et al. Reference Shea, Jongejans, Skarpaas, Kelly and Sheppard2010; Skarpaas and Shea Reference Skarpaas and Shea2007; Zhang and Shea Reference Zhang and Shea2012; see Supplementary Material for model details). Dispersal parameters were assumed to be the same as for the Pennsylvania experimental population in Jongejans et al. (Reference Jongejans, Shea, Skarpaas, Kelly, Sheppard and Woodburn2008), except for plant height, as plant heights differed significantly from the control of the present experiment (Table 1). Surrounding vegetation height was assumed to be 5 cm for all mowing treatments to mimic heavily grazed pastures where this species is most successful. Mowing started the latest in treatments Hl and Ll (the high-and low-intensity, late-mow treatments, respectively), where pre-mow reproduction was significantly different from zero (1.9 ± 0.3 and 2.3 ± 0.4, respectively). Therefore, seeds from these capitula could have dispersed from a taller height (than plant height at the end of the growing season) before being mowed. However, we did not consider differences in dispersing height between pre-mow and post-mow capitula in the model, and thereby may underestimate c* for these two treatments in particular.
Results and Discussion
Mowing in our previous experiment generally decreased plant survival, reproduction, and height (Zhang and Shea Reference Zhang and Shea2012); all mowing treatments reduced plant performance relative to the control of no mowing. However, there was considerable variation in the impact achieved by the different mowing interventions. All empirical results are presented in Table 1, allowing comparisons of any treatments according to any chosen measure. Results addressing our objective-dependent management questions are presented in Figure 1. While Table 1 and Figure 1 allow for all possible comparisons to be made, we discuss key points here.
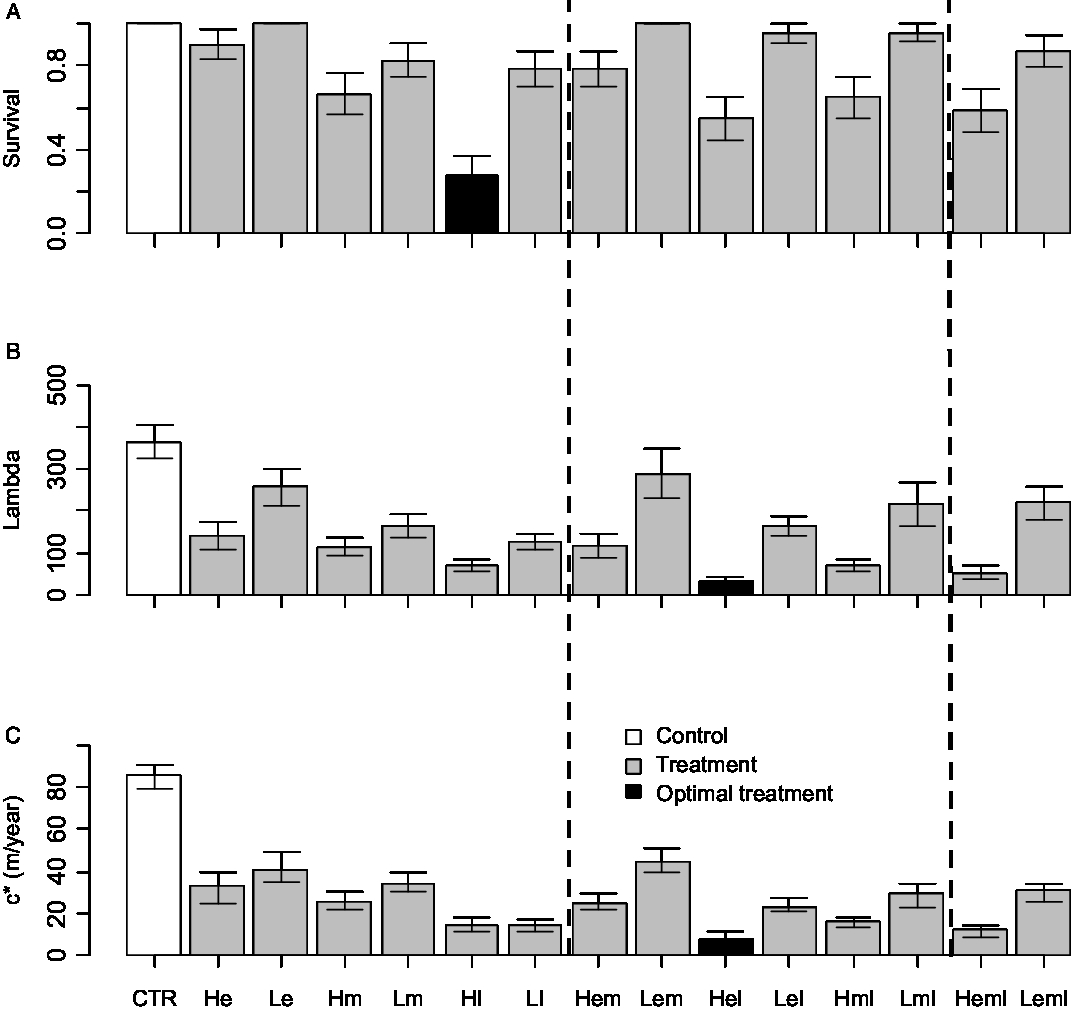
Figure 1. Evaluation of effectiveness of mowing treatments (percentage reductions compared with the control) based on three management goals of different time horizons: (A) to reduce density of adult plants in the present season (i.e., adult survival); (B) to reduce the long-term population growth rate λ; and (C) to reduce the long-term population spread rate c*. Error bars show standard errors (either from the raw data [for survival] or calculated from the model [for lambda and c*]). The white bar denotes the control (no mowing), and black bars denote the most effective mowing treatments, which resulted in the largest percentage reductions. Vertical dashed lines separate treatments that were mowed one, two, or three times. Note that in C, the spread rates for treatment HI and LI may be underestimated, as the release height of pre-mow capitula in these treatments was assumed to be the same as plant height at the end of the season. See Table 1 for definitions of treatment abbreviations.
Empirical mowing treatments with higher intensity and mowing treatments including a late cut reduced survival and reproduction more than those with lower intensity and those without a late cut (Table 1). In fact, several low-intensity mowing treatments had plant survival outcomes that were not significantly different from the control (P > 0.05). As a result, reductions in population growth rates λ were larger in treatments with higher intensity and treatments including a late cut (Figure 1B). Treatment Hl (with high intensity, late mowing) had the largest impact on survival of existing plants (a reduction of 71%; Table 1; Figure 1A). Treatment Hel (with high-intensity mowing both early and late in the season) caused the largest reduction in lifetime capitulum production (a reduction of 92%; Table 1) and hence on population growth rate (a reduction of 91%; Table 1, Figure 1B).
Reductions in plant height followed the same pattern as for survival and reproduction—treatments with higher intensity and treatments including a late cut caused a larger reduction than those with lower intensity and those without a late cut (Table 1). Treatment Hel caused the largest reduction in plant height (a reduction of 76%; Table 1), and therefore this treatment had the greatest impact on seed dispersal. Coupled with the largest reduction in population growth, treatment Hel therefore caused the largest reduction in population spread rate (a reduction of 91%; Table 1; Figure 1C).
Generally, mowing treatments with higher intensity and late timing had a larger impact on survival, reproduction, plant height, population growth rate, and population spread. However, the choice of the most effective mowing regime depends on the management objective under consideration. Treatment Hl is most effective for reducing population density of existing adult plants (i.e., lowest survival) and therefore for attaining immediate weed “control.” A high-intensity, late cut removed most biomass and caused the most damage, because it started late in the season when plants had already invested significantly in biomass. This objective may be the first choice for managers concerned with current-season minimization of the number of plants in the field to reduce grazing productivity loss.
In our study, we used reductions in population growth rate λ and population spread rate c* to evaluate the long-term effectiveness of the treatments. The baseline matrix model parameters are from an experiment in which thistles were grown with abundant resources and little competition (Jongejans et al. Reference Jongejans, Shea, Skarpaas, Kelly, Sheppard and Woodburn2008). Therefore the high λ values in our results represent the worst-case scenario for an invasion, specifically fast population growth during the initial part of the invasion period in disturbed habitats. As the invasion continues, this invasive species will likely face strong competition, especially during seedling establishment, that can greatly affect its performance and hence reduce population growth (Peterson-Smith and Shea Reference Peterson-Smith and Shea2010; Ruggiero and Shea Reference Ruggiero and Shea2011). Nevertheless, projections of λ and the corresponding c* (which integrates population growth and dispersal) assuming the status quo provide powerful tools for management decisions.
Treatment Hel is the most effective for reducing population growth in the long run (i.e., lowest λ) and for reducing population spread in the long run (i.e., lowest c*). This is because treatment Hel (which includes both an early and a late mow) more successfully reduced lifetime reproduction than treatment Hl, which was penalized by pre-mow capitula that could release seed before treatment even started. Compared with a single late mow, an extra early mow as well as the late mow helps to remove early-dispersing capitula. Therefore, if we consider another short-term management goal, to reduce seed production in the current season, Hel is again the most effective strategy out of all 14 treatments. Furthermore, the extra mow may have hastened depletion of the resource bank of the species (both nutrients and meristem pool; Huhta et al. Reference Huhta, Lennartsson, Tuomi, Rautio and Laine2000) for later regrowth. Due to the large elasticity of the reproductive vital rates for the study system (Jongejans et al. Reference Jongejans, Shea, Skarpaas, Kelly, Sheppard and Woodburn2008), treatment Hel causes a greater reduction in population growth than treatment Hl. In contrast to a previous study of biocontrol agents in which management strategy rankings differ for reducing local population abundance and reducing population spread (Shea et al. Reference Shea, Jongejans, Skarpaas, Kelly and Sheppard2010), the most effective strategy for both longer-term objectives is the same mowing regime. This is because the treatment reducing reproduction the most (Hel) in the present study also affected a dispersal parameter, plant height, immensely. Due to the significant contribution of plant height to spread (Zhang et al. Reference Zhang, Jongejans and Shea2011), treatment Hel thus causes the largest reduction in population spread rate. Furthermore, as the release height of pre-mow capitula in these treatments was assumed to be the same as plant height at the end of the season, the reductions for treatment Hl may be overestimated. Therefore, the advantage of treatment Hel over treatment Hl in controlling spread could be even larger if such variations in dispersal height were taken into account.
Counterintuitively, the treatment with the highest frequency and intensity (treatment Heml) was never the most effective option for achieving any of the three management goals. The treatment comprising an early and a late mow is more effective than mowing three times in terms of reaching all three management goals. This is probably because frequent removal of apical meristems in this treatment induced more flowering stems than the other treatments (Zhang and Shea Reference Zhang and Shea2012). Thus our assessment of a wide range of mowing management interventions not only improves outcomes, but also saves logistical effort.
The most effective management regime based on our spread model outputs focuses on localized population spread, which corresponds to the infilling between established populations and is a key component of invasion spread (Shigesada and Kawasaki Reference Shigesada and Kawasaki1997). However, spread of invasive species on a regional scale also requires extreme long-distance dispersal events (Mortensen et al. Reference Mortensen, Rauschert, Nord and Jones2009; Nathan et al. Reference Nathan, Horvitz, He, Kuparinen, Schurr and Katul2011; Rauschert et al. Reference Rauschert, Mortensen, Bjørnstad, Nord and Peskin2010). For example, human-facilitated long-distance dispersal (e.g., transportation of hay bales contaminated with thistle seeds) may cause new infestations in new areas, which acts on top of the infilling processes occurring on the local scale. Furthermore, spatial and environmental heterogeneity may also complicate regional population spread. Therefore a thorough control effort will require restrictions on both the infilling and colonizing processes.
In this study, we only considered the performance of the targeted weed species, C. nutans. But in reality, management decisions also rely on a suite of other factors, such as forage yield, forage quality, and animal consumption patterns (Fulkerson and Donaghy Reference Fulkerson and Donaghy2001). For example, mowing only once late in the season results in large thistles at the time of the treatment, which may compromise the quality of forage. On the other hand, including an early mow as well as a late mow reduces the size of the thistles, and therefore may improve forage quality. Therefore future studies should include further examination of the effects of weed management practices on the rest of the forage-producing plant community. Furthermore, weed management with late timing often requires handling of more developed fruits compared with management starting earlier in the season (Pyšek et al. Reference Pyšek, Krinke, Jarosik, Perglova, Pergl and Moravcova2007). Because capitula of C. nutans have the potential to continue developing on cut-off stems (RZ, personal observation), managers should carefully destroy the harvested biomass to prevent seed maturation, dispersal, and further infestations. As a consequence, managers intending to control this species should also consider practical implementation aspects.
The choice of an optimal management strategy requires a clear statement of objective (Milner-Gulland and Shea Reference Milner-Gulland and Shea2017; Probert et al. Reference Probert, Shea, Fonnesbeck, Runge, Carpenter, Dürr, Garner, Harvey, Stevenson, Webb, Werkman, Tildesley and Ferrari2016; Shea et al. Reference Shea, Jongejans, Skarpaas, Kelly and Sheppard2010). While “weed control” is often used to describe the management action aimed to eliminate an existing weed population (Buhler Reference Buhler2002), a more precise statement is required in terms of time horizon and spatial extent. For example, does the manager aim at a quick, short-term reduction in plant abundance, or long-term control whose effects may not be obvious during the first few years of management? Does the manager only care about the problem in a limited area where the weed is present, or is he or she also concerned about its spread into adjacent fields? Such questions are important for the appropriate choice of management option. Our study demonstrates that (1) all mowing treatments were more effective than no mowing; (2) nevertheless, some mowing treatments were more effective than other mowing treatments; and (3) a mowing treatment’s effectiveness depends on the specific objective. Intense mowing both early (before flowering) and late (at flowering peak) is the most effective management strategy of the 14 mowing regimes we examined for the long-term control of C. nutans in terms of both local abundance and spatial spread. However, short-term management outcomes from a single, intense, late mow may be acceptable, especially when managers are constrained by limited budgets and labor.
Author ORCID
Katriona Shea, https://orcid.org/0000-0002-7607-8248
Acknowledgments
RZ and KS conceived and designed the experiment. RZ conducted the experiment and analyzed the data. RZ wrote the first draft, and both authors edited this article. Many Shea lab undergraduates helped in the field and the lab. We are grateful for discussion and comments from Ottar Bjørnstad, Eric Post, David Mortensen, Stephen Roxburgh, Adam Miller, Suann Yang, Eelke Jongejans, and Joe Keller. We acknowledge the support of USDA-CSREES (Biology of Weedy and Invasive Plants) NRI grant 2002-35320-12289, NSF grant DEB-0815373 and NSF-NERC award DEB-1556444. No conflicts of interest have been declared.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/inp.2019.18




