



Introduction
Dairy production in temperate regions is characterized by seasonal-calving, pasture-based systems which endeavour to match pasture growth with feed demand (Läpple et al., Reference Läpple, Hennessy and O'Donovan2012; Hennessy et al., Reference Hennessy, Delaby, van den Pol-van Dasselaar and Shalloo2020). However, intensive grassland dairy production systems are characterized as being reliant on nitrogen (N) fertilizer use to support forage dry matter (DM) growth and high animal stocking rates, resulting in losses of N to the surrounding environment via leaching, volatilization and gaseous emissions (Delaby et al., Reference Delaby, Finn, Grange and Horan2020; Jaramillo et al., Reference Jaramillo, Sheridan, Soder and Dubeux2021). Grazing forages containing clover and herb species require lower N inputs to grow forage and therefore can reduce N loss from pasture (Grace et al., Reference Grace, Boland, Sheridan, Lott, Brennan, Fritch and Lynch2018; Carlton et al., Reference Carlton, Cameron, Di, Edwards and Clough2019; Moloney et al., Reference Moloney, Sheridan, Grant, O'Riordan and O'Kiely2021). Additionally, a reduction in urinary N (UN) and improved dietary N use efficiency (NUE) has been reported for cows grazing forages containing herb species (Dodd et al., Reference Dodd, Dalley, Wims, Elliott and Griffin2018; Wilson et al., Reference Wilson, Bionaz, MacAdam, Beauchemin, Naumann and Ates2020). Dairy cows are inefficient users of dietary N, with an estimated nitrogen use efficiency (NUE) ranging from 0.2 to 0.4 (Whelan et al., Reference Whelan, Mulligan and Pierce2013; Fadul-Pacheco et al., Reference Fadul-Pacheco, Pellerin, Chouinard, Wattiaux, Duplessis and Charbonneau2017). Therefore interest is growing in integrating herbs such as chicory and plantain into grazing dairy production systems by either using them as a summer feed and/or transitioning them into permanent pastures (Mangwe et al., Reference Mangwe, Bryant, Beck, Beale, Bunt and Gregorini2019).
In Ireland, compact calving results in cows entering late lactation simultaneously. In late lactation, milk yield declines and milk processability characteristics, such as rennet coagulation time (RCT), milk pH and ethanol stability (ES), deteriorate (Visentin et al., Reference Visentin, De Marchi, Berry, McDermott, Fenelon, Penasa and McParland2017; Doran et al., Reference Doran, Mulligan, Lynch, O'Sullivan, Fahey, McKay, Brady, Grace, O'Rourke and Pierce2021). For processors, the irregularity in milk supply and processability reduces their ability to react to market demand, as some products, such as cheese, cream and milk powder, cannot be manufactured due to poor milk quality, often leading to the storage of product to meet demand out of season (Donnelly and Horne, Reference Donnelly and Horne1986; Auldist et al., Reference Auldist, Walsh and Thomson1998; Timlin et al., Reference Timlin, Tobin, Brodkorb, Murphy, Dillon, Hennessy, O'Donovan, Pierce and O'Callaghan2021). Altering the supply of dietary energy and protein in late lactation can positively influence milk production, composition and processability characteristics (Stumpf et al., Reference Stumpf, Fischer, McManus, Kolling, Zanela, Santos, Abreu and Montagner2013), and therefore is of interest to both producers and processors. Additionally for processors, it is of interest to alter the fatty acid (FA) profile of milk, as it has an important role in the quantity and quality of milk produce manufactured. For instance, saturated FAs have a large influence over the texture and spreadability of butter (Bobe et al., Reference Bobe, Hammond, Freeman, Lindberg and Beitz2003), while FAs such as linoleic, linolenic and vaccenic acid have been shown to have beneficial effects on human health, enhancing the value of dairy produce (Johnson and Fritsche, Reference Johnson and Fritsche2012; Toral et al., Reference Toral, Monahan, Hervás, Frutos and Moloney2018). Milk from pasture-fed cows has been reported to have higher concentrations of total protein and casein, and increased concentrations of saturated FAs (C11:0 – C17:0) and unsaturated FAs (C18:2n-6 trans, C18:3n-3, C20:1, C20:4n-6 and C18:2 cis-9,trans-11) when compared to milk from cows offered total – mixed rations (O'Callaghan et al., Reference O'Callaghan, Hennessy, McAuliffe, Kilcawley, O'Donovan, Dillon, Ross and Stanton2016; Gulati et al., Reference Gulati, Galvin, Lewis, Hennessy, O'Donovan, McManus, Fenelon and Guinee2018). However, little is known about the impact of feeding multispecies forages on milk processability characteristics. Therefore, the objective of this experiment was to compare the effects of a perennial ryegrass, perennial ryegrass-white clover and multispecies forage on the milk production and milk processability characteristics of late lactation dairy cows. It was hypothesized that the inclusion of clover and herb species in a perennial ryegrass based grazing forage would result in an increase in the unsaturated FA concentration of milk from cows grazing these forages compared to those grazing perennial ryegrass only.
Materials and methods
Experimental design
This experiment was a randomized complete block design, with three dietary treatments. Fifteen primiparous and thirty multiparous lactating dairy cows in late lactation were selected from the autumn calving herd at University College Dublin (UCD) Lyons Farm and randomly assigned one of three dietary treatments (n = 15 per treatment). Cows were blocked on DIM (234 DIM; ± 17) and parity, before being balanced for milk yield, milk composition (fat (g/kg), protein (g/kg) and milk solids yield (kg/d)) and body condition score (BCS). Milk parameters for balancing cows were averaged from the previous two monthly milk recordings carried out in March and April.
The experiment was conducted over 56-d, which included a 14-d adaptation period during which cows were adapted to their respective dietary treatment. After the pre-experimental period, a 35-d measurement period occurred where milk production parameters, forage chemical composition, rumen fermentation parameters and blood metabolite concentrations were measured. Following this, cows (n = 3 per group) were grazed in replicate groups (n = 5 per treatment) for 7-d to allow the estimation of forage DMI.
Dietary treatments
Cows were randomly assigned to one of three forage-based dietary treatments as follows;
1. PRG: cows grazing a perennial ryegrass monoculture forage, supplemented with approximately 3 kg of DM/d a grass-silage based total-mixed ration (TMR; grass silage, straw and molasses), plus a dairy concentrate pellet fed at 2.7 kg of DM/cow per day (fed in the milking parlour, half offered during each milking);
2. PRGWC: cows grazing a perennial ryegrass and white clover forage supplemented with a TMR and concentrate, as above; and
3. MULTI: cows grazing a mixed forage of perennial ryegrass, timothy, white clover, red clover, plantain and chicory supplemented with a TMR and concentrate, as above.
Table 1 shows the chemical composition of each treatment offered, including grazing forages, grass-silage TMR and concentrate pellet. Cows were offered the TMR to prevent bloating which is a risk associated with high clover forages. The TMR consisted of grass silage (2 kg DM), straw (0.75 kg DM) and molasses (0.25 kg DM), and was offered to cows post milking each morning. Cows were separated into their respective treatment groups in the cubicle barn before the TMR was offered via an open feed space. The TMR was mixed and fed out using a diet feeder (Keenan, Alltech Farming Solutions, Carlow, Ireland), which weighed the feed as it was fed out, allowing equal amounts of the TMR to be offered to each treatment group. Silage, straw and TMR samples were collected daily, while concentrate samples were collected weekly. Samples were then dried at 55°C for 72 h to allow determination of DM, before being pooled according to week.
Table 1. Chemical composition of the treatment forages, TMR and concentrate offered to cows during the experiment

Values presented for the treatment forages (grass, grass and white clover, and grass, legume and herb) are Bonferroni adjusted means ± standard error of the mean (S.E.M.)
Forage establishment and management
Forages were previously established on UCD Lyons Research Farm in May (2.4 ha) and August (10.35 ha) of 2019. Two land blocks were each separated into three equal sections and randomly sown so that approximately 4.25 ha was sown with each forage type. All forages received slurry (9.8 kg N/ha) in January 2021. Nitrogen fertilizer, in the form of urea with a urease inhibitor (0.38 N; 30 kg N/ha), was applied in March. Half of each forage type was grazed by the existing dairy herd in April, with the other half harvested for silage. No further cutting or grazing of forages occurred until the experiment began on 25 May.
In May 2021, each forage area was fenced into equal sized paddocks of approximately 0.8 ha for grazing. Permanent fences separated the paddocks and electric fences were used to separate daily pasture allocations. Cows were offered an estimated pasture allocation of 12 kg of DM/cow per day. Daily pasture allowances were offered based on pre-grazing forage mass and adjusted daily based on pasture DM and estimated daily pasture growth. Daily pasture DM was initially estimated from data reported on the PastureBase Ireland website. Pre-grazing forage samples were collected and dried in a forced air oven at 55°C for 72 h to determine forage DM, with this figure then used to adjust forage DM estimations. Animals were offered fresh forage allocations each day post morning and evening milking. Plastic water troughs were moved daily into the grazed area to allow ad libitum access to water.
Pre-grazing forage mass was estimated for each forage type at the beginning of the experiment and every 2–3 days thereafter as new paddocks entered the grazing rotation. An area (0.25 m2) was cut using a quadrat and shears to 4 cm at three random locations in the area to be grazed. The samples were weighed, mixed and a 100 g sub-sample collected for DM determination and subsequent chemical analysis, and a 150 g sub-sample gathered to estimate the botanical composition of the forage. The sample was separated into component species before being dried at 55°C for 72 h to determine the DM proportion of the species in each forage type. In total, 15 forage separations were carried out on the PRG forage, 13 on the PRGWC forage, and 14 on the MULTI forage over the course of the experiment. Post-grazing residuals were measured daily using a rising plate meter (RPM; plate diameter of 355 mm and area density of 3.2 kg/m2; Jenquip, Feilding, New Zealand). A total of 50 platemeter measurements were recorded in a W pattern on the area after being grazed.
Intake measurement period
From day 50–56, all animals were grazed in replicate groups of three animals per group (n = 5 groups per treatment) to allow the estimation of forage intake, as described by Dodd et al. (Reference Dodd, Dalley, Wims, Elliott and Griffin2018) and Muir et al. (Reference Muir, Ward and Jacobs2015). During this measurement period paddocks were break-fed in 24-hour breaks, beginning after the morning milking. Break sizes were modified daily, based on harvested DM and estimated daily forage growth, to allow approximately 12 kg DM intake/cow/d.
Initial pre-grazing forage mass was measured by cutting two plots of 5 m × 2 m with a lawnmower to 4 cm height. The cut forage was then weighed and subsampled for DM determination. This was repeated once the group were half-way through the paddock. Group forage DMI was estimated by determining pre and post-grazing forage mass with a RPM by collecting 100 measurements in each daily allocation of pasture during the measurement period. A total of 20 RPM readings were recorded across the area in each of the replicate breaks pre and post grazing, giving a total 100 readings across the entire plot. The RPM was calibrated by linear regression against pasture mass at varying heights by collecting 30 quadrats (each 0.25 m2, 15 pre-grazing and 15 post-grazing quadrats) per forage treatment. Quadrats were cut to 3 cm residual height with electric hand shears along a diagonal transect in the area to be measured. Calibration equations for each forage type are as follows;
-
PRG – Forage cover (kg DM/ha) = 188.61 (platemeter height (cm)) + 230.94 (r2 = 0.71)
-
PRGWC – Forage cover (kg DM/ha) = 187.12 (platemeter height (cm)) + 290.29 (r2 = 0.77)
-
MULTI – Forage cover (kg DM/ha) = 141.88 (platemeter height (cm)) + 145.17 (r2 = 0.76)
Apparent DM intake of replicates was calculated from forage disappearance between pre- and post-grazing forage and area allocated such that:
Forage DMI = (pre-grazing mass (kg DM/ha) – post-grazing mass (kg DM/ha)) × area/no. cows.
Data and sample collection
Animal measurements
Cow bodyweight (BW) was recorded at the beginning and end of the experiment using an electronic scale (Tru-Test, New Zealand). Body condition score was assessed by a single, experienced operator, using a scale of 1 to 5 with 0.25 increments according to Edmonson et al. (Reference Edmonson, Lean, Weaver, Farver and Webster1989), following morning milking every 14-days.
On day 15, 29 and 43, blood and rumen samples were harvested from all animals. Blood samples were collected via jugular venipuncture for urea N and glucose. Blood samples were harvested into a 4-mL glucose Vacutainer containing sodium fluoride for glucose analysis (BD Vacutainer Systems, Plymouth, UK), which were stored on ice and centrifuged at 2100 × g for 20 min at 4°C for extraction of blood plasma. The blood plasma was frozen at −20°C, pending further analysis.
Rumen fluid samples were harvested using a rumen scoop oral oesophageal sampler (Prof-Products, Ontario, Canada). Rumen fluid samples were analysed immediately for pH (Phoenix Instrument EC-25 Ph/Conductivity Portable Meter). Once collected, samples were strained through four layers of cheesecloth, and a 4-mL aliquot was collected using an automatic pipette, mixed with 1 mL of trichloroacetic acid, and cooled on ice. These were stored at −20°C pending analysis for VFA and rumen ammonia.
Milk measurements
Milk yield was recorded each day. Cows were milked twice daily at approximately 07:00 and 15:00 in a 60 – unit rotary milking parlour (Dairymaster, Causeway, Kerry, Ireland). Measurement of milk yield and composite milk sample collection was carried out using the Weighall milk meter system (Dairymaster, Causeway, Kerry, Ireland). During the sampling period, milk samples were collected from consecutive am and pm milkings weekly for composite analysis. Samples from each animal were pooled by day in proportion to yield before being stored at – 20°C pending further analysis. During week 7 of the experiment, additional pooled samples from individual cows were prepared for milk FA analysis. Forage samples from the same period were also gathered and prepared for forage FA analysis.
Sample analyses
Dried samples of forage, silage, straw and concentrate were ground in a hammer mill fitted with a 1-mm screen (Lab Mill, Christy Turner, Ltd., Ipswich, UK). Sample DM was determined after drying overnight at 105°C (16 h minimum) (AOAC International, 2005, method 930.15). The ash content of forage and concentrate samples was determined after incineration of a 3 g sample in a muffle furnace (Nabertherm GmbH, Lilienthal, Germany) at 550°C for 5-hours (AOAC International, 2005a; method 942.05). The N content of forage and concentrate samples was measured using a Leco FP 828p Analyzer instrument (Leco Instruments UK, Cheshire, UK) and CP (N × 6.25) subsequently calculated. Neutral detergent fibre and ADF content were determined using the method of Van Soest et al. (Reference Van Soest, Robertson and Lewis1991) using the Ankom 220 Fiber Analyzer (Ankom Technology, Fairport, NY). For analysis of NDF, concentrate samples were analysed with a thermo-stable α-amylase and 20 g of NaSO3 was added to neutral detergent solution. Neutral detergent fibre and ADF are expressed inclusive of residual ash. DM digestibility (DMD) of forage was estimated using ADF according to the equation; DMD = 88.9 – (0.779 × ADF %) (Linn and Martin, Reference Linn and Martin1991). Starch content of the concentrate was determined using the Megazyme Total Starch Assay Procedure (product no. K-TSTA, Megazyme International Ltd., Wicklow, Ireland). This assay is a two-step enzymatic process where a-amylase is first added to the sample, followed by amyloglucosidase at a later step. The sample solutions are then read against a reagent blank using a spectrophotometer (absorbance = 510 nm; UVmini – 1240, Shimadzu, Japan). Water-soluble carbohydrate of forage was analysed according to the method used by Birch and Mwangelwa (Reference Birch and Mwangela1974). Briefly, forage samples were soaked in distilled water before being centrifuged. Concentrated sulphuric acid and 5% aqueous phenol were added to the supernatant and incubated at 35°C before water – soluble carbohydrate (WSC) concentration was measured by spectrophotometry (absorbance = 485 nm; UVmini – 1240, Shimadzu, Japan). Ether extract was measured using a Soxtec instrument (Tecator) according to the method of AOAC 107 (AOAC, 1970). Non-fibre carbohydrates (NFC) were calculated using forage NDF, CP, ash and EE according to the equation described in Wilson et al. (Reference Wilson, Bionaz, MacAdam, Beauchemin, Naumann and Ates2020) such that NFC = 100 – ((% NDF – 2) + CP + Ash + EE).
Concentrations of milk fat, protein, lactose and somatic cell count (SCC) were determined in a commercial milk laboratory (Independent Milk Laboratories, Cavan, Ireland) using mid-infrared spectrophotometry (CombiFoss 5000, Foss Analytical A/S, Hillerød, Denmark). Milk solids yield (kg/d) was calculated as the sum of daily fat yield (kg/d) and daily protein yield (kg/d). Energy corrected milk (ECM) was calculated according to the equation (0.3273 × Milk yield kg) + (7.65 × Protein kg) + (12.97 × Fat kg) (Tyrrell and Reid, Reference Tyrrell and Reid1965). Fat corrected milk (FCM) was calculated according to Gaines (Reference Gaines1928), such that 4% FCM = (0.4 × Milk yield kg) + (15 × Fat yield kg).
Milk processability analysis was carried out weekly and included analysis of milk pH, RCT and ES. Fresh milk was used for all milk processability analysis, with milk refrigerated at 4°C pending analysis. A sub-sample of the milk collected for weekly milk composite analysis was used for milk processability analysis. Within each treatment, cows were randomly assigned to one of five subgroups by blocking on parity and balancing for DIM, BCS and milk production parameters. Individual cow milk samples from each treatment were pooled in replicate groups of three per group (n = 5, per treatment). The purpose of the subgroups was to facilitate milk processability analysis. All milk processability analysis was carried out in the same laboratory, using the same equipment and by the same technician who had been previously trained to visually determine milk flocculation.
Milk pH was determined using a portable pH meter (Phoenix Instrument EC-25 pH/Conductivity Portable Meter). The RCT was determined according to a modified version of the method described by Berridge (Reference Berridge1952). Five mL of rennet (Hansen's Naturen 145 rennet; Chr. Hansen Holding A/S, Hørsholm, DK) was diluted with 100 mL of distilled water to give a 1/20 rennet dilution. This solution (0.5 mL) was added to a test-tube containing a 5 mL sample of milk which had been preheated to 30°C in a water bath. Upon addition of the rennet solution to the preheated milk, a timer was simultaneously started. The milk sample was slowly inverted twice before being attached to a rotating holder and immersed in the water bath at a 30° angle with rotation set to maximum speed (4 rpm). Coagulation of milk was determined to have occurred when flocks of renneted standard milk substrate appeared on the wall of the rotating test-tube and the time taken for coagulation to occur was recorded. Milk ES was determined according to the method reported by Guo et al. (Reference Guo, Wang, Li, Qu, Jin and Kindsted1998). Briefly, equal volumes of the milk were mixed with an ethanol solution (ranging in concentration from 62% to 84%, v/v) at room temperature. The ES of milk was determined at the maximum concentration of ethanol solution that did not cause the appearance of precipitate in the sample being analysed.
FA analysis of whole milk and herbage samples was conducted in the AFBI Newforge Lane Laboratory (Agri-Food and Biosciences Institute, Belfast, Northern Ireland). Milk was collected from individual cows during the afternoon milking on day 43 and the morning on day 44, then pooled in proportion to morning and afternoon milk yield. Corresponding forage samples were gathered on day 43 and 44 from the pasture allocation offered to cows, dried in a forced air oven at 55°C for 72 h and then ground in a hammer mill fitted with a 1-mm screen (Lab Mill, Christy Turner, Ltd., Ipswich, UK). Dried samples were bulked 50:50 before being sent for analysis. A total of 45 milk samples and 3 forage samples were sent for FA analysis. All samples were analysed in randomized batches, in duplicate. The average of the duplicate values was then used for statistical analysis.
Fatty acid methyl esters (FAME) in forage were extracted according to the protocol described in Palmquist and Jenkins (Reference Palmquist and Jenkins2003), while FAME in milk were analysed using the chloroform method described in Bligh and Dyer (Reference Bligh and Dyer1959). Total FAME concentrations were measured using an Agilent 7890 GC with Flame Ionisation Detector (FID) fitted with a CP-Sil 88 capillary column for FAME (100 m × 0.25 mm: 0.2 μm, Agilent technologies, USA). Individual FAMEs were identified by comparison of their retention times with those of pure methyl ester standards (Supelco 37 Component FAME Mix, Sigma-Aldrich, USA). Fatty acid methyl esters in milk are reported as a percentage of identified FAME in each sample. Total FA values in forage are reported in mg/g and were calculated for each sample using internal standard C19:0 with applied corrections using experimental and calculated conversion ratios.
Blood samples were analysed in the UCD veterinary pathology laboratory (School of Veterinary Medicine, Dublin, Ireland) using Randox kits (Randox Laboratories Ltd, Crumlin, UK) according to manufacturer's instructions. Blood urea in serum was measured via enzymatic procedure (kit no. UR1068) while glucose was analysed using the hexokinase test (kit no. GL3816).
Rumen fluid was thawed overnight in a refrigerator at 4°C before being centrifuged 2100 × g for 10 min at 4°C prior to analysis. One ml of supernatant was diluted with 4 ml distilled H2O and then centrifuged at 1600 × g for 15 min at 4°C. Next, 200 μl of supernatant was combined with 3 reagents (Reagent 1 = 13 g of NaOH + 4 g (di) Sodium EDTA made up to 1L with dH2O; Reagent 2 = 10 g phenol + 0.05 g Sodium Nitroprusside made up to 1L with dH2O; Reagent 3 = 5 g NaOH + 10 ml Sodium Hypochlorite solution made up to with dH2O). Rumen ammonia concentrations were then determined using a spectrophotometer (absorbance = 680 nm; UVmini – 1240, Shimadzu, Japan). Rumen fluid samples were also prepared for VFA analysis by mixing 250 μl of the same supernatant used for NH₃-N determination with 3.75 ml of distilled water and 1 ml of internal standard solution (0.5 g 3-methyvaleric acid in 1000 ml of 0.15 M oxalic acid). The resulting solution was centrifuged at 2100 rpm for 5 min at room temperature and then filtered through a syringe tip filter (polytetrafluoroethylene, 25 mm diameter, 0.45 μm) into 2 ml gas chromatography (GC) vials. Concentration of VFA's was determined using Scion 456-GS (Scion Instrument, Scotland, UK) fitted with a DB-FFAP capillary column (15 m × 0.53 mm: 1.00 μm, Agilent Technologies, USA).
Statistical analysis
Analysis of data was carried out using the MIXED procedure of SAS (version 9.1.3, SAS Institute Inc., Cary, NC). Initially, a univariate regression model was used where each predictor variable was included as a single fixed effect, and variables that had P ⩽ 0.25 were selected for use in the multivariate analysis. Subsequently, the model was developed using a manual stepwise model building procedure. All factors with a P ⩽ 0.25 were retained in the final model. Parity was retained in the model as a fixed effect due to its biological significance. Fixed-effect testing was based on the F-test with denominator degrees of freedom approximated by the Satterthwaite's procedure. After fitting the models for each dependent variable, residuals were plotted against predicted values to check for normality and homogeneity of variance by histograms, QQ-plots, and formal statistical tests as part of the UNIVARIATE procedure of SAS. All data analysed were deemed to have a normal distribution.
When repeated measures were included in a model, a covariance structure was subsequently fitted to the model. The type of variance-covariance structure used was chosen from compound symmetry, unstructured, autoregressive, heterogeneous first order autoregressive, or Toeplitz variance-covariance structures, and the model with the lowest Bayesian information criterion value was selected as appropriate for the analysis. The PDIFF option and the Bonferroni test were applied as appropriate to evaluate multiple comparisons. Effects with a P-value <0.25 were retained in the model. Statistical significance was declared at P ≤ 0.05 and a tendency was assumed at 0.05 < P ≤ 0.10. Results are presented as least square means ± standard error of the mean (SEM).
The model for forage nutritive value was as follows;

where Yij is the response variable being analysed, μ is the mean, Ti is the fixed effect of treatment, Wj is the fixed effect of week, Ti × Wj is the fixed effect of the interaction between treatment and week, and еij is the residual error. Week was included as a repeated measure.
Cow was considered the experimental unit for milk production, milk composition, blood and rumen data analysis. The model for these analyses was as follows;

where Yijklm is the response variable being analysed, Ti is the fixed effect of treatment, Wj is the fixed effect of week, Bk is the fixed effect of block, Pl is the fixed effect of parity, Cm is the random effect of cow, Ti × Wj is the fixed effect of interaction between treatment and week, and еijklm is the residual error. Days in milk and days in calf were also included as covariates in the model. Milk production, milk quality, blood and rumen function were analysed using repeated measures. For analysis of milk yield, day was included as a repeated measure. Week was included as a repeated measure for milk composition analysis. Time point was used as a repeated measure for analysis of blood and rumen parameters, and cow BCS and BW.
Sub-group was considered the experimental unit for milk processability and DMI analysis. The model for these analyses was as follows;

where Yijk is the response variable being analysed, μ is the mean, Ti is the fixed effect of treatment, Wj is the fixed effect of week, Sk is the random effect of sub-group, Ti × Wj is the fixed effect of interaction between treatment and week, and еijk is the residual error. For DMI analysis, day was included as a repeated measure instead of week and days in calf and days in milk were included as covariates.
Cow was considered the experimental unit for milk FA analysis however this analysis did not include a repeated statement. The model for this analysis was as follows;

where Yijkl is the FA being analysed, μ is the mean, Ti is the fixed effect of treatment, Bj is the fixed effect of block, Pk is the fixed effect of parity, Cl is the random effect of cow, Ti × Bj is the fixed effect of the interaction between treatment and block, and еijkl is the residual error.
Results
Diet chemical composition
The chemical composition of the treatment forages, TMR and dairy concentrate offered to cows during the experiment are shown in Table 1. Mean forage DM was lower for the grass and white clover (162 g/kg), and grass, legume, herb forages (162 g/kg) compared to grass only (217 g/kg). Crude protein was significantly higher (P = 0.007) for grass and white clover (182 g/kg DM), and grass only (128 g/kg DM) when compared to the grass, clover, herb forage. The NDF concentration of grass and white clover (413 g/kg DM; P = 0.002) was significantly lower than both the grass only (496 g/kg DM) and grass, legume, herb (464 g/kg DM) forages. The concentration of non-fibre carbohydrates (NFC) was similar between forage types, though the grass only forage had the highest WSC concentration (202 g/kg DM; P = 0.001) of all forage types. The DMD concentration of the grass only (733 g/kg DM) and grass and white clover (740 g/kg DM) forages was significantly higher (P < 0.001) than that of the grass, legume, herb forage (713 g/kg DM).
The concentration of C14:0 in the grass (0.084 mg/ml) and grass and white clover (0.084 mg/ml) forages were significantly higher than the grass, legume, herb forage (0.065 mg/ml) (Table 2). Both the grass and white clover and grass, legume, herb forages had a significantly higher (P < 0.05) concentrations of C17:0, C18:2 cis – 9, 12, and saturated fatty acids than the grass only forage. The polyunsaturated fatty acid (PUFA) (13.62 mg/ml) and total fatty acid concentration (19.18 mg/ml) of grass only was higher (P < 0.05) than the grass and white clover (12.60 mg/ml and 18.47 mg/ml) and grass, legume, herb forages (12.66 mg/ml and 18.46 mg/ml).
Table 2. The fatty acid concentration (mg/ml) of grass, grass and white clover, and grass, legume and herb forages
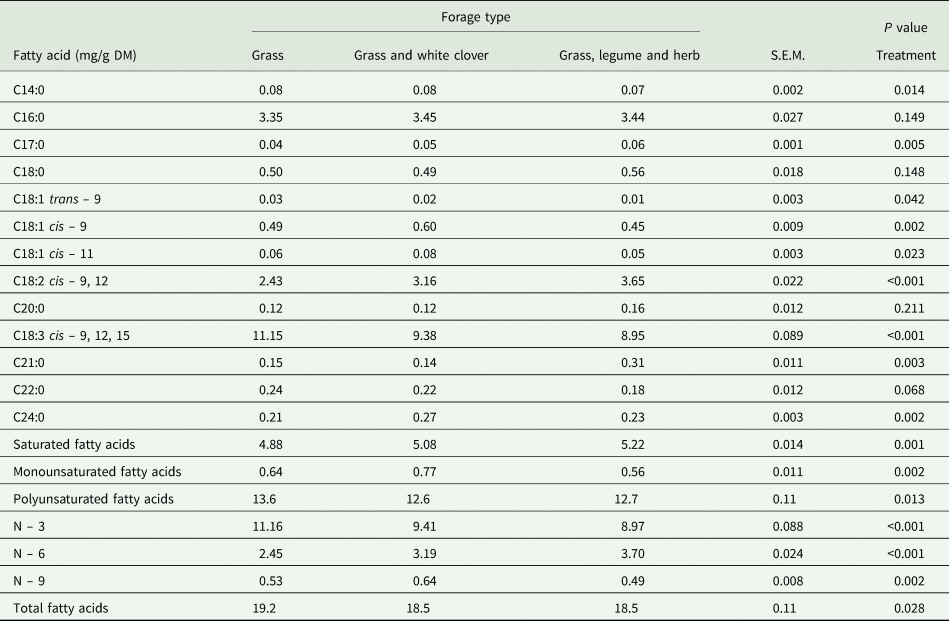
Values presented are Bonferroni adjusted means ± standard error of the mean (S.E.M.)
Forage botanical composition and intake
Mean pre-grazing forage mass (kg DM/ha) for each forage type during weeks 1 −5 of the experiment was as follows; grass only = 1502 kg DM/ha, grass and white clover = 2168 kg DM/ha, and grass, legume, herb = 2033 kg DM/ha. Mean group grazed forage DMI was highest for those offered the grass and white clover forage (11.05 kg DM/d), followed by the grass, legume, herb (10.41 kg DM/d), and grass only (9.69 kg DM/d) forages. On a DM basis, the proportion of perennial ryegrass in the grass only forage was 0.96. The grass and white clover forage was predominately composed of white clover (0.6 as a proportion of DM), with perennial ryegrass comprising 0.4 of the forage on a DM basis. On average, the proportion of species in the grass, legume, herb forage was as follows; 0.26 perennial ryegrass, 0.08 timothy, 0.5 red clover, 0.05 white clover, 0.04 chicory and 0.04 plantain on a DM basis.
Cow BCS, milk production and milk FA profile
Cow BW and BCS were unaffected by treatment (P > 0.05). Milk yield was significantly higher for cows offered PRGWC (18.07 kg/d) and MULTI (17.84 kg/d) compared to PRG (17.62 kg/d) (Table 3). Milk protein yield was significantly higher for PRGWC (0.75 kg/d; P = 0.012) compared to PRG (0.67 kg/d). Milk pH was also higher for PRGWC (6.79) compared to PRG (6.74; P = 0.004) and MULTI (6.75; P = 0.014). Both the fixed effect of day, and the interaction of day and treatment, had a significant effect (P ≤ 0.001) on all yield (kg/d) parameters reported, while parity had a significant effect on daily milk yield (kg/d) and milk solids yield (kg/d). There was no difference in ECM, 4% FCM, milk fat and lactose yield (kg/d), milk composition (g/kg), milk solids production (kg/d), RCT (mins) or ES (%) between treatments.
Table 3. Milk production (kg/d), milk composition (g/kg) and milk processability results of cows offered PRG, PRGWC or MULTI

Values presented are Bonferroni adjusted means ± standard error of the mean (S.E.M.)
The concentration of saturated fatty acids (mg/ml) in milk, including C6:0, C8:0, C10:0, C11:0, C12:0, C13:0, C14:0, and C16:0, was unaffected by treatment (P > 0.05) (Table 4). The concentration of C18:1 trans – 9 in the milk of cows offered MULTI (0.256 mg/ml) was higher than that of cows offered PRGWC (0.211 mg/ml; P = 0.017). The concentration of C18:1 trans – 11 was significantly higher (P = 0.018) for cows offered PRG (1.00 mg/ml) compared to either PRGWC (0.768 mg/ml) or MULTI (0.774 mg/ml). Cows offered MULTI had a significantly higher concentration of C18:2 cis – 9, 12 (P = 0.002), C18:3 cis – 9, 12, 15 (P < 0.001), and PUFA (P = 0.001) compared to PRG, and a numerically higher concentration of these fatty acids compared to PRGWC. The ratio of N6: N3 in milk was lowest (P < 0.001) for PRGWC (1.08) compared to either PRG (1.26) or MULTI (1.16).
Table 4. Mean fatty acid concentration of milk (mg/ml) from cows offered PRG, PRGWC and MULTI

Values presented are Bonferroni adjusted means ± standard error of the mean (S.E.M.)
Blood metabolites and rumen fermentation
Blood urea nitrogen was significantly higher for PRGWC (4.86 mmol/L) and MULTI (4.76 mmol/L) compared to PRG (2.03 mmol/L; P < 0.001). Similarly, blood glucose concentration was higher (P < 0.001) for PRWC (3.33 mmol/L) and MULTI (3.25 mmol/L) than PRG (2.79 mmol/L).
There was no difference in rumen pH between treatments (Table 5). The rumen ammonia concentration of cows offered PRGWC (2.00 mmol/L) and MULTI (1.92 mmol/L) was higher than those offered PRG (0.91 mmol/L; P < 0.001). There was no difference between forage treatments in the rumen fluid concentration (mmol/L) of acetic acid, propionic acid or total VFAs. Both the rumen fluid concentration and proportion of butyric acid was significantly higher for cows offered PRG (P < 0.001) when compared to those offered PRGWC or MULTI. Offering cows PRG resulted in a lower proportion of acetic acid in rumen fluid (61.88) than either PRGWC (66.21) or MULTI (67.45). The proportion of propionic acid in the rumen fluid of cows offered MULTI (17.80) was lower than that of cows offered PRG (19.07; P = 0.002) and PRGWC (18.82; P = 0.015). The A:P ratio differed significantly between all treatments (P < 0.001). MULTI resulted in the highest A:P ratio (3.84), with PRGWC intermediate (3.55), and PRG having the lowest (3.25).
Table 5. Rumen fermentation characteristics of cows offered PRG, PRGWC or MULTI

Values presented are Bonferroni adjusted means ± standard error of the mean (S.E.M.)
Discussion
Forage DMI, milk production and milk processability
The forage DMI of cows grazing the grass and white clover and grass, legume, herb forages was significantly higher than those grazing the grass only forage, despite the higher pre-grazing covers for both the grass and despite clover and grass, legume, herb forages. Cows offered both PRGWC and MULTI also had significantly higher concentrations of blood glucose compared to PRG, suggesting a higher level of dietary digestible energy intake for these animals. The high proportions of white clover (0.6) and red clover (0.5) in the diverse forages may have compensated for the higher pre-grazing covers of these forages as clover species are readily digestible in the rumen (Dewhurst et al., Reference Dewhurst, Delaby, Moloney, Boland and Lewis2009) and are known to have lower concentrations of structural fibre than grass species (Villalba et al., Reference Villalba, Ates and MacAdam2021).The nutrient composition of plant leaf and stem material influences animal grazing selection, with selection differentials typically higher for CP and energy and lower for NDF as animals prefer to consume leaf material over stem (Muir et al., Reference Muir, Ward and Jacobs2015). This could explain the reduced forage DMI, and subsequently milk production, of cows grazing the grass only forage as this forage had the lowest CP and highest NDF of all forage types.
The lower structural fibre concentration of white clover species, coupled with the increased milk production of animals offered PRGWC, likely impacted the milk fat concentration of cows grazing this forage, with PRGWC resulting in a numerically lower milk fat concentration than PRG. Similarly, cows offered MULTI in this experiment also had a reduced milk fat concentration, despite the grass, legume, herb forage having a significantly higher NDF and ADF concentration than the grass and white clover forage. This has been previously reported in cows offered forages containing white clover and plantain (Dineen et al., Reference Dineen, Delaby, Gilliland and McCarthy2018; Nguyen et al., Reference Nguyen, Navarrete, Horne, Donaghy and Kemp2022). Cows offered both PRGWC and MULTI had a significantly lower proportion of butyric acid in their rumen than those offered PRG. Previous authors have reported a reduction in rumen butyric acid concentration when increasing the dietary proportion of clover in silage for dairy cows (Vanhatalo et al., Reference Vanhatalo, Kuoppala, Ahvenjärvi and Rinne2009; Johansen et al., Reference Johansen, Søegaard, Lund and Weisbjerg2017). Butyrate is one of the primary carbon sources for de novo milk fat synthesis in the mammary gland (Bauman and Griinari, Reference Bauman and Griinari2003), and the reduced availability of this in the rumen may have negatively impacted the milk fat concentration of cows offered PRGWC and MULTI.
However, milk protein yield was higher for cows grazing PRGWC and MULTI compared to PRG, likely due to the higher milk yield of these cows as milk protein concentration (g/kg) was similar between treatments. However, the elevated rumen ammonia and blood urea N (BUN) of cows on the PRGWC and MULTI treatments is likely due to an increased dietary CP intake due to the high clover content (proportion of white and red clover >0.5 respectively) in these forages. This suggests these cows would also have a high milk urea nitrogen (MUN) concentration compared to PRG. Both BUN and MUN are known to be positively correlated (Kauffman and St-Pierre, Reference Kauffman and St-Pierre2001; Barros et al., Reference Barros, Reed, Olmos Colmenero and Wattiaux2019). Increased MUN presents an issue for processors as it is positively associated with the proportion of non-protein nitrogen (NPN) in milk protein. Significantly higher NPN has previously been reported in the milk of cows grazing grass-clover forages compared to those grazing a grass monoculture (O'Callaghan et al., Reference O'Callaghan, Hennessy, McAuliffe, Kilcawley, O'Donovan, Dillon, Ross and Stanton2016). Increased proportions of NPN are undesirable for dairy processors, with increased NPN resulting in poorer quality protein and a reduction in the concentration of casein and whey protein in milk, (Reid et al., Reference Reid, O'Donovan, Murphy, Fleming, Kennedy and Lewis2015; O'Callaghan et al., Reference O'Callaghan, Hennessy, McAuliffe, Kilcawley, O'Donovan, Dillon, Ross and Stanton2016). A higher casein content in milk is important for further processing, and milk with a higher casein content is associated with reduced coagulation times, and a firmer curd, which are important for the production of cheddar cheese and yoghurt (Timlin et al., Reference Timlin, Tobin, Brodkorb, Murphy, Dillon, Hennessy, O'Donovan, Pierce and O'Callaghan2021).
Milk pH was highest for PRGWC, though still within an acceptable range for processing, and comparable to previous studies carried out in late lactation dairy cows in Ireland (pH 6.6–6.7; McKay et al., Reference McKay, Lynch, Mulligan, Rajauria, Miller and Pierce2019; Doran et al., Reference Doran, Mulligan, Lynch, O'Sullivan, Fahey, McKay, Brady, Grace, O'Rourke and Pierce2021). Both the RCT and ES of milk are positively related to milk pH (Donnelly and Horne, Reference Donnelly and Horne1986; Visentin et al., Reference Visentin, De Marchi, Berry, McDermott, Fenelon, Penasa and McParland2017). However, despite significant differences in milk pH neither ES nor RCT varied significantly between treatments. Milk ES is used in many countries globally as a test of raw milk quality and an indicator of the thermal stability of milk (Da Costa et al., Reference Da Costa, De Paula, Da Silva, Perrone, Stephani and Cappa De Oliveira2022; Pinheiro et al., Reference Pinheiro, Rocha, De Andrade, Rotta, Rezende, Pires and Marcondes2022). As milk is pasteurized before sale, thermal stability is essential in dairy processing. Milk with an acceptable ES (≥72% v/v) is generally considered suitable for ultra-high temperature and milk powder processing (Chavez et al., Reference Chavez, Negri, Taverna and Cuatrín2004; Stumpf et al., Reference Stumpf, Fischer, McManus, Kolling, Zanela, Santos, Abreu and Montagner2013). ES is reported to decrease from 78% to 72% as cows move from early – mid lactation, to late lactation (Li et al., Reference Li, Ye and Singh2019). Supplementing late lactation dairy cows with concentrates while grazing has previously been reported to improve ES (O'Brien et al., Reference O'Brien, Dillon, Murphy, Mehra, Guinee, Connolly, Kelly and Joyce1999; Doran et al., Reference Doran, Mulligan, Lynch, O'Sullivan, Fahey, McKay, Brady, Grace, O'Rourke and Pierce2021), with an increase in dietary energy supply known to have beneficial effects on milk ES (Stumpf et al., Reference Stumpf, Fischer, McManus, Kolling, Zanela, Santos, Abreu and Montagner2013)
Similarly, RCT also extends as lactation progresses with coagulation times increasing from 4–6 min in mid-lactation to 9–15 min in late lactation (O'Brien et al., Reference O'Brien, Crosse and Dillon1996; Visentin et al., Reference Visentin, De Marchi, Berry, McDermott, Fenelon, Penasa and McParland2017). Rennet coagulation of milk is a two-stage process, with the primary phase involving the cleavage of hydrophilic casein on the surface of protein micelles, before the calcium-induced aggregation and gelation of destabilized micelles occurs in phase two (Magan et al., Reference Magan, O′Callaghan, Kelly and McCarthy2021; Britten and Giroux, Reference Britten and Giroux2022). Decreasing pH, increasing temperature, and a higher concentration of calcium ions in milk all improve coagulation and reduce RCT (McSweeney, Reference McSweeney and McSweeney2007). Previous authors investigating the effect of pasture type on RCT reported similar coagulation times of milk when cows were offered grass only or a grass and herb species mixed forage containing chicory and plantain (Sanjayaranj et al., Reference Sanjayaranj, Lopez-Villalobos, Blair, Janssen, Holroyd and MacGibbon2023), or when cows were supplemented with concentrates and alfalfa while grazing ryegrass pasture (Auldist et al., Reference Auldist, Greenwood, Wright, Hannah, Williams, Moate and Wales2016). Though RCT was not significantly different between treatments in this experiment, milk from cows offered PRGWC took almost 100s longer on average to coagulate compared to that from PRG or MULTI. As previously discussed, the urea fraction of milk from cows grazing clover rich forages is higher compared to grass only pastures. Milk is often heated prior to the addition of rennet in order to increase the subsequent cheese yield (Magan et al., Reference Magan, O′Callaghan, Kelly and McCarthy2021). When heated, urea in milk is converted to ammonia, resulting in a buffering effect and an extended heat coagulation times and improved ethanol and thermal stability (Magan et al., Reference Magan, O′Callaghan, Kelly and McCarthy2021). This reaction is pH dependant, and at a pH of 6.6–7.2 the urea concentration does not alter the coagulation reaction (Muir and Sweetsur, Reference Muir and Sweetsur1977). Within this pH range, Karlsson et al. (Reference Karlsson, Lundh, Innings, Höjer, Wikström and Langton2019) found no difference in heat coagulation time or ES with increasing urea concentrations. Similarly in our study, the concentration of urea in milk from PGRWC was perhaps not high enough to significantly improve ES or RCT when compared to PRG or MULTI at a pH of 6.8. The lower RCT of cows offered MULTI could be due to the lower rapidly degradable protein fraction of red clover, despite both red and white clover resulting in similar concentrations of rumen ammonia (Dewhurst et al., Reference Dewhurst, Evans, Scollan, Moorby, Merry and Wilkins2003). In addition, in this experiment cows were all in late-lactation, with stage of lactation likely to supersede the potential effect of pasture type on the ES and RCT of milk despite differences in protein degradability. Irrespective of dietary treatment, milk was of an acceptable quality for processing, indicating that including clover and herb species in grazing forages does not negatively impact milk processability. However, this experiment did not investigate the effect of alternative grazing forages on other characteristics of milk, such as taste. In New Zealand and Australia, it is recommended that chicory remain below 50% of the cows diet in order to reduce the risk of a bitter taint in milk (NSW Department of Industry, 2017). Previous research indicated that the sesquiterpene lactones in some chicory varieties resulted in a ‘bitter’ taint in milk (Barry, Reference Barry1998). Subsequent chicory breeding programmes focused on breeding new varieties with lower levels of sesquiterpene lactones, suitable for use on dairy farms, where the low levels of these chemicals would be most unlikely to cause a taint or bitter aftertaste in the milk (Rumball et al., Reference Rumball, Keogh, Miller and Claydon2003). However, Mangwe (Reference Mangwe2020) found no studies which compared the effects of chicory cultivars on milk taint. Further investigation would be useful to fully assess the effect of multispecies forages on milk processability and sensory characteristics of milk.
Milk fatty acids and rumen fermentation
In this experiment, C18:1 trans-11 (vaccenic acid; VA) was the predominating trans FA in cow's milk. Milk from PRG had a higher concentration of both C18: trans-9 (elaidic acid; EA) and VA, and a numerically higher concentration of CLA. Reducing dietary intake of trans FA is recommended for humans, as they are associated with a range of negative health conditions, including cardiovascular disease, breast cancer and diabetes (Dhaka et al., Reference Dhaka, Gulia, Ahlawat and Khatkar2011; de Souza et al., Reference de Souza, Mente, Maroleanu, Cozma, Ha, Kishibe, Uleryk, Budylowski, Schünemann, Beyene and Anand2015). The primary trans FA in ruminant produce is VA, which is a precursor to beneficial CLA cis-9, trans-11 (Lock et al., Reference Lock, Horne, Bauman and Salter2005). Therefore, foods rich in beneficial CLA will also have a higher content of VA (Lock et al., Reference Lock, Horne, Bauman and Salter2005). The biohydrogenation of the PUFAs C18:2 cis-9, 12 (linoleic acid; LA) and C18:3 cis-6, 9, 12 (ɑ-linolenic acid; ALA) by FA desaturases leads to the formation of VA (Jenkins et al., Reference Jenkins, Wallace, Moate and Mosley2008). Both PRGWC and MULTI resulted in increased concentrations of PUFA, LA and ALA compared to PRG, suggesting reduced biohydrogenation of PUFA in the rumen of these cows. This is consistent with forage analysis, as the PRG forage had the highest concentration of PUFA. Reduced biohydrogenation of PUFAs is desirable, as PUFA are critical nutrients that modulate brain development and cognition, as well as other diseases such as cardiovascular disease and diabetes (Lee et al., Reference Lee, Lee, Kang and Park2016). Both LA and ALA are considered essential FAs as they are necessary for health, but must be provided by foods as they cannot be synthesized in the human body (Kaur et al., Reference Kaur, Chugh and Gupta2014). Wilson et al. (Reference Wilson, Bionaz, MacAdam, Beauchemin, Naumann and Ates2020) reported a reduction in desaturase activity in cows grazing forages containing legume and herb species compared to those grazing grass. Increased PUFA has also been reported in the milk of cows offered forages containing chicory (Muir et al., Reference Muir, Ward and Jacobs2014; Mangwe et al., Reference Mangwe, Bryant, Beck, Fleming and Gregorini2020) and red clover (Lee et al., Reference Lee, Theobald, Tweed, Winters and Scollan2009). A reduction in biohydrogenation has been linked to the combined effect of the plant secondary compounds polyphenol oxidase and tannins in red and white clover (Lee et al., Reference Lee, Theobald, Gordon, Leyland, Tweed, Fychan and Scollan2014; Toral et al., Reference Toral, Monahan, Hervás, Frutos and Moloney2018). In addition, previous authors have reported a higher hourly degradation rate for forages containing plantain and chicory (Minneé et al., Reference Minneé, Waghorn, Lee and Clark2017; McCarthy et al., Reference McCarthy, Lynch, Pierce, Fahey, McDonald, Boland, Sheridan, Markiewicz-Keszycka and Mulligan2023), and white clover (Dewhurst et al., Reference Dewhurst, Evans, Scollan, Moorby, Merry and Wilkins2003) compared to perennial ryegrass dominant forages. The rapidly degradable nature and faster rumen passage rate of herb and clover species has been suggested as an explanation for the increased LA and ALA concentration in the milk of cows offered diverse forages as forage lipids of forage lipids are exposed for less time to lipolysis, and subsequently biohydrogenation, in the rumen (Dewhurst et al., Reference Dewhurst, Evans, Scollan, Moorby, Merry and Wilkins2003; Mangwe et al., Reference Mangwe, Bryant, Beck, Fleming and Gregorini2020). The PRGWC and MULTI forage also had higher concentrations of LA when compared to PRG. Therefore, the differences reported in PUFA concentration in milk are potentially a combination of plant secondary compounds, increased rumen passage rate and variations in FA intake due to forage type.
Though the total concentration of rumen acetic and propionic acid was similar for PRG, PRGWC and MULTI, the acetic: propionic acid ratio (A:P) varied between treatments. Forages rich in WSC and with a low CP concentration are reported to have lower proportions of rumen acetic acid, increased proportions of propionic acid and subsequently a lower rumen A:P ratio (Lee et al., Reference Lee, Merry, Davies, Moorby, Humphreys, Theodorou, MacRae and Scollan2003; Rivero et al., Reference Rivero, Keim, Balocchi and Lee2020). In this experiment, the PRG forage had a significantly higher concentration of WSC and lower CP concentration compared to the PRGWC and MULTI forages, which could help account for differences in the A:P ratio between treatments. Increasing the supply of dietary RDP results in a higher concentration of rumen ammonia and branched chain VFA, such as iso-butyric and iso-valeric acid, formed from the deamination and subsequent catabolism of peptides and branched chain amino acids (Bach et al., Reference Bach, Calsamiglia and Stern2005; Putri et al., Reference Putri, Zain, Warly and Hermon2021). Legumes, such as white and red clover, have a high concentration of soluble protein but are low in fibre (Dewhurst et al., Reference Dewhurst, Delaby, Moloney, Boland and Lewis2009) and differences in the concentration of rumen ammonia and branched chained VFA between treatments are likely a reflection of compositional differences between forage types.
Conclusion
Including clover and herb species in perennial ryegrass grazing forages increased the forage DMI, milk yield and milk protein yield of late lactation dairy cows supplemented with grass silage and concentrates. Irrespective of treatment, milk was of an acceptable level for processing despite PRGWC resulting in an increase in milk pH. However, the elevated rumen ammonia and BUN of cows grazing PRGWC and MULTI species suggests a concurrent increase in the NPN fraction of milk protein compared to those grazing PRG, which could present an issue for dairy processors. Despite this, cows grazing forages containing clover and herb species had a more beneficial milk fatty acid profile, with increased concentrations of PUFA likely due to a reduction in rumen biohydrogenation.
Authors’ contributions
K.M. McCarthy – study design, data collection, sample analysis, statistical analysis, wrote the article. N. Walsh – study design, data collection. E. Serra - data collection, sample analysis. A.G. Fahey – study design, statistical analysis, review and editing of article. M.B. Lynch – review and editing of article. K.M. Pierce – study design, review and editing of article. T.M. Boland – project funding, study design, review and editing of article. H. Sheridan - project funding, study design. M. Markiewicz-Keszycka – sample analysis. F.J. Mulligan – study design, data collection, statistical analysis, writing, review and editing.
Funding statement
The experimental work reported herein was funded by the Department of Agriculture, Food, and the Marine's Competitive Funding Programs (17/S/267) in conjunction with commercial industry.
Competing interests
None.
Ethical standards
All procedures described in this experiment were approved by the Animal Research Ethics Committee at University College Dublin (UCD). These procedures were conducted under experimental license (AE18982/ P182) from the Health Products Regulatory Authority (SI No. 543 of 2012).






 Open access
Open access