


It is believed that, due to a variety of physiological and structural differences, the hindgut of ruminants may be more susceptible to factors affecting gut barrier integrity compared to the rumen (Sanz-Fernandez et al., Reference Sanz-Fernandez, Daniel, Seymour, Kvidera, Bester, Doelman and Martin-Tereso2020). Given what is known about the importance of hindgut health in non-ruminant species, specifically targeting the ruminant hindgut with prophylactic interventions is an opportunity that merits exploration. We have previously observed increases in yields of milk fat (Seymour et al., Reference Seymour, Carson, Daniel, Sanz-Fernandez, Martín-Tereso and Doelman2020, Reference Seymour, Sanz-Fernandez, Daniel, Martín-Tereso and Doelman2021) and/or protein (Sanz-Fernandez et al., Reference Sanz-Fernandez, Seymour, Daniel, Doelman and Martín-Tereso2022; Seymour et al., Reference Seymour, McKnight, Carson, Sanz-Fernandez, Daniel, Metcalf, Martín-Tereso and Doelman2022) when supplementing lactating dairy cattle with approximately 16 g/d hydrogenated fat-embedded calcium gluconate (HFCG), a prebiotic compound specifically targeting the increased production of acetate and/or butyrate in the hindgut (Asano et al., Reference Asano, Yuasa, Kunugita, Teraji and Mitsuoka1994; Tsukahara et al., Reference Tsukahara, Koyama, Okada and Ushida2002, Reference Tsukahara, Hashizume, Koyama and Ushida2006). However, the underlying mechanism(s) of these responses remain unclear at present. The objectives of the current study were to evaluate the changes in yields of milk and milk components, and alterations in concentrations of blood and fecal metabolites, in a potentially dose-dependent response to HFCG. We hypothesized that milk fat content and yield, and concentrations of fecal butyrate and plasma non-esterified fatty acids would be increased by HFCG in a linear dose-dependent manner.
Materials and methods
This experiment was conducted at the Trouw Nutrition Agresearch Dairy Facility (Burford, ON) between May and December 2015. All animal procedures were approved by the Institutional Animal Care and Use Committee at Trouw Nutrition Agresearch, in accordance with Canadian Council on Animal Care guidelines. Forty-eight lactating Holstein cattle (9 primiparous, 39 multiparous) were enrolled in the experiment at 21 ± 2 d after calving, however, two multiparous cattle were subsequently removed due to unrelated health reasons. Animals were individually housed in tie stalls for the duration of the experiment. Both water and feed in the form of a total mixed ration (Table 1) were offered ad libitum. Fresh feed was delivered daily at approximately 0900 h, and orts were weighed to determined voluntary dry matter intake (DMI). Animals were milked in place twice daily at approximately 0600 h and 1600 h.
Table 1. Formulation and chemical composition of total mixed ratios
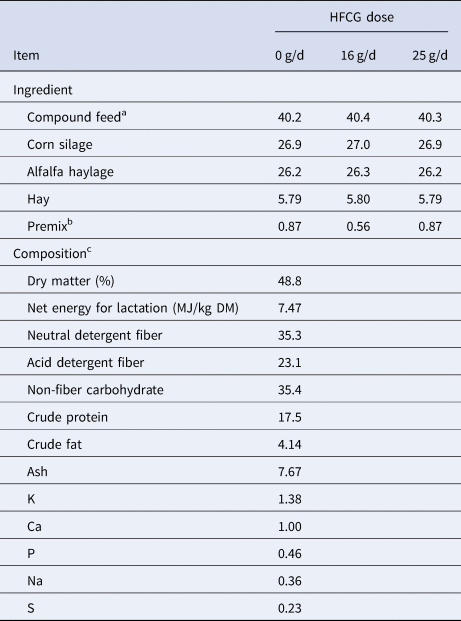
Values are presented on a percent dry matter basis unless indicated otherwise.
a Contained (% as-fed) ground corn (37.4), fine rolled corn (17.3), corn gluten feed (10.4), corn dried distillers grains (8.32), pork meal (4.99), bypass soybean meal (5.08; Top Soy, Trouw Nutrition, Puslinch, ON), bakery waste (3.33), palm fat supplement (2.13; APF + , Trouw Nutrition), fat supplement (1.25; Stay Fat, Darling Ingredients Canada, Cambridge, ON), salt (1.16), sodium sesquicarbonate (1.07), calcium carbonate (1.06), magnesium oxide (0.69), blood meal (0.54), urea (0.33), mineral premix (0.24; Nutri-Plex Micro NS, Trouw Nutrition), organic acid supplement (0.16; RM104, Trouw Nutrition), selenium (0.13), yeast (0.10; BioPower SC10, Lallemand Animal Nutrition, Montréal, QC), methionine (0.09; Alimet, Novus International Inc., St. Charles, MI), monensin (0.02; Rumensin, Elanco Animal Health, Greenfield, IN)
b 0 g/d: contained (% as-fed) limestone (54.9), rice hulls (42.5), soybean oil (2.60); 16 and 25 g/d: contained limestone (49.7), rice hulls (37.3), hydrogenated fat-embedded calcium gluconate (12.0; Trouw Nutrition) and soybean oil (1.00)
c Composition of HFCG-supplemented TMRs did not differ significantly from control
A 3 × 3 replicated Latin square design was used to evaluate performance responses to two levels of HFCG relative to a negative control over three periods of 28 d each. Cattle were blocked by calving date and treatment sequences were randomly assigned to animals within blocks. HFCG supplement (50% hydrogenated palm oil, 40% calcium gluconate, 10% calcium carbonate; Trouw Nutrition) was incorporated into a premix (Table 1) and then added to the base ration to achieve a target dose of either 0 g/d (negative control), 16 g/d, or 25 g/d of HFCG. Samples of blood, milk and feces were collected over the last 3 d of each period, as described in the Supplementary File. Data were analyzed as described in the Supplementary File.
Results and discussion
Responses in DMI and milk production in multiparous cows are presented in Table 2. Milk fat content was significantly increased in response to 16 g/d HFCG (0.19%; P < 0.05), resulting in a 6.2% increase in milk fat yield (P < 0.05). As DMI was unaffected by treatment, the increased yields resulted in a 6% increase (P < 0.05) in gross feed efficiency. Yields of milk fat, 4% fat-corrected and energy-corrected milks, as well as gross feed efficiency and milk fat content, displayed evidence of a potential non-linear dose response to HFCG supplementation, as indicated by the lack-of-fit partition (Table 2). Increased milk fat yield has previously been observed in lactating dairy cattle supplemented with 16 g/d HFCG (Seymour et al., Reference Seymour, Carson, Daniel, Sanz-Fernandez, Martín-Tereso and Doelman2020, Reference Seymour, Sanz-Fernandez, Daniel, Martín-Tereso and Doelman2021), which was previously attributed to the increased incorporation of pre-formed fatty acids of endogenous origin. In the present study, no differences were observed in circulating acetate, beta-hydroxybutyrate or non-esterified fatty acids (online Supplementary Table S1). We did not record bodyweight or body condition score to assess the potential mobilization of body reserves. As such, the nature of the milk fat response remains unclear at this point in time. In primiparous animals, no differences were observed in production parameters (online Supplementary Table S2), however, fecal butyrate concentration displayed evidence of a potentially non-linear dose response (online Supplementary Table S3). However, this study lacked sufficient statistical power to make any valid claims pertaining specifically to the responses (or lack thereof) in heifers, and as such these results should be interpreted with caution. All responses observed at 25 g/d HFCG supplementation did not differ from control.
Table 2. Dry matter intake and production responses in multiparous Holstein cows (n = 37) supplemented with 3 levels of hydrogenated fat-embedded calcium gluconate (HFCG)

Values are presented in units of kg/d unless indicated otherwise
a 16 g/d: 0 g HFCG/d vs. 16 g HFCG/d; 25 g/d: 0 g HFCG/d vs. 25 g HFCG/d; LIN, linear dose response; LOF, lack-of-fit of linear dose response.
b Standard error of the difference.
c Dry matter intake.
d 4% fat-corrected milk: 0.4 × milk yield (kg/d) + 15 × fat yield (kg/d).
e Energy-corrected milk: 0.01 × milk yield (kg/d) + 12.2 × fat yield (kg/d) + 7.7 × protein yield (kg/d) + 5.3 × lactose yield (kg/d).
f Gross feed efficiency.
The precise mode of action of the HFCG supplement remains unclear. It is currently hypothesized that gluconic acid salts support the production of acetate and butyrate, which have been implicated in the promotion of gut health and integrity (as reviewed by Liu et al., Reference Liu, Wang, He, Becker, Zhang, Li and Ma2018; Litvak et al., Reference Litvak, Byndloss and Baumler2018), as well as modulation of whole-body energy metabolism (as reviewed by den Besten et al., Reference den Besten, van Eunen, Groen, Venema, Reijngoud and Bakker2013). However, the main challenges in evaluating the response to this product are twofold: it is difficult to get an accurate determination of in vivo volatile fatty acid synthesis with spot samples due to their rapid uptake by the gastrointestinal epithelium (den Besten et al., Reference den Besten, van Eunen, Groen, Venema, Reijngoud and Bakker2013), and the signalling pathways involved are poorly understood across species (den Besten et al., Reference den Besten, van Eunen, Groen, Venema, Reijngoud and Bakker2013; Litvak et al., Reference Litvak, Byndloss and Baumler2018; Liu et al., Reference Liu, Wang, He, Becker, Zhang, Li and Ma2018). Due to this, it is difficult to draw conclusions as to why responses to 25 g/d HFCG did not differ from control.
In conclusion, supplementing lactating multiparous dairy cows with 16 g/d HFCG increased both the yield and concentration of milk fat. Yields of milk fat, 4% fat-corrected and energy-corrected milks, as well as gross feed efficiency and milk fat content, displayed evidence of a potential non-linear dose response to HFCG supplementation, contrary to our hypothesis of a linear dose response over this range. Additionally, no responses due to treatment were observed for concentrations of fecal butyric acid or plasma non-esterified fatty acid, contrary to our hypothesis. More work is required to characterize the response to HFCG supplementation both at the level of the gastrointestinal epithelium, as well as the downstream signalling pathways in the host animal. Both the nature of the non-linear dose response and potential parity-by-treatment interactions require additional work to confirm.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0022029922000851
Acknowledgements
The authors would like to thank J. A. Metcalf, L. L. McKnight, D. F. Waterman and the staff of Trouw Nutrition Agresearch for their contributions to this study.




 Open access
Open access