



Introduction
In Mexico, the last National Survey demonstrated that obesity prevalence in women of reproductive age increased by 9.6% from 2012 to 2021, with the prevalence of obesity in 2021 in this group at 41.1%, Reference Shamah-Levy, Romero-Martínez and Barrientos-Gutiérrez1 indicating that a significant proportion of women who are planning to be pregnant have obesity. Overall, this disease is considered one of the leading public health problems in the country. Obesity exposes fetuses to an obesogenic environment in utero, which leads to an increased risk of several diseases in the progeny (fetal programing). Reference Ahmed, Liang and Chi2–Reference De Los Santos, Reyes-Castro, Coral-Vázquez, Méndez and Leal-García4
It has been described that diet-induced obesity leads to skeletal muscle deregulation and subsequent development of muscle atrophy, a condition associated with decreased protein synthesis and accelerated degradation of myofibrillar and soluble proteins. This occurs through various mechanisms such as ubiquitin-proteasome pathway over activation, oxidative stress, myonuclear apoptosis, and autophagy, among others. Reference Sishi, Loos, Ellis, Smith, Du Toit and Engelbrecht5–Reference Morales, Monsalves-Álvarez and Tadinada7
Moreover, an obesogenic maternal environment causes impaired skeletal muscle development, characterized by a decrease in lean mass and muscle fibers, an increase in intramuscular lipid content, a reduction in muscle cell proliferation, and impaired markers of postnatal myogenesis in the offspring; notably, a healthy postnatal diet might not reverse any of these effects. Reference Bayol, Simbi and Stickland8–Reference Mikovic, Brightwell and Lindsay10
It has been described that the AKT/PI3K signaling cascade is relevant to muscle catabolic/anabolic balance since this activation induces phosphorylation of FOXO1, causing its translocation to the cytoplasm, inhibiting its function as a transcription factor for MURF and MAFbx, which are ubiquitin ligases specific for degradation of multiple skeletal muscle proteins. Besides, activation of AKT/PI3K induces activation of p70S6K, which is related to the development of muscle fibers. Reference Egerman and Glass11 Because of an obesogenic maternal diet, a decrease in AKT phosphorylation in skeletal muscle has been demonstrated in animal models. Reference Salto, Girón and Manzano12 This effect over the pivotal AKT activation, related to an obesogenic maternal diet, suggests that several downstream molecules could be affected, like p70S6K, FOXO1, MAFbx, and MURF, which have been evaluated in direct obesity models, but not in maternal obesity (MO) models. Furthermore, NFkB participates in muscle atrophy induced by a high-fat diet (HFD). Reference Sishi, Loos, Ellis, Smith, Du Toit and Engelbrecht5
On the other hand, the impact of obesity during pregnancy in offspring is necessary to identify specific components in food, like the natural phytochemicals, that might help prevent several chronic disorders, including obesity and its comorbidities. Reference Cremonini, Iglesias and Kang13 One of these products is (−)-epicatechin (Epi), a flavonoid that has shown beneficial effects on skeletal muscle. Reference Gutierrez-Salmean, Ciaraldi and Nogueira14,Reference Lee, Leem and Go15 Likewise, it has been demonstrated, in a model of obesity due to a HFD, that Epi has a protective effect against skeletal muscle loss and physical performance decline induced by the lipotoxicity state associated with obesity. Reference Munguia, Ramirez-Sanchez, Meaney, Villarreal, Ceballos and Najera16 In addition, our group Reference De los Santos, Coral-Vázquez and Menjivar9 evidenced that prolonged treatment with Epi (13 weeks) could increase lean mass in offspring descendants of MO in the context of fetal programing; however, the possible mechanisms performed by Epi in this phenomenon were not explored.
Considering that these mechanisms that predispose to muscle damage in the offspring of obese mothers are not well known we previously demonstrated that the offspring of obese mothers present obesity and metabolic disorders. Reference De los Santos, Coral-Vázquez and Menjivar9,Reference Rodríguez-González, De Los Santos and Méndez-Sánchez17 We investigated whether the offspring of mothers with obesity, induced by a HFD, presented disorders in the expression of genes related to atrophy (Murf1, MAFbx, NFκB and Foxo1) or in protein synthesis (p70S6K-alpha) in the gastrocnemius muscle, and if these disorders were modified with the administration of the flavonoid (−)-epicatechin using two different protocols (short and long). We hypothesized that the treatment with (−)-epicatechin will increase the expression of genes related to skeletal muscle protein synthesis and will decrease the expression of genes associated with pathological atrophy of this tissue in offspring obese by programing in comparison with the offspring without treatment with this flavonoid. However, we did not find that Epi modified the expression of genes involved in the skeletal muscle’s atrophy and/or protein synthesis.
Materials and methods
Animal model
The Salvador Zubirán National Institute of Medical Sciences and Nutrition Animal Experimentation Ethics Committee previously approved this study (INCMNSZ) with the approval number CINVA: UIO-1892-17/19-1. Regarding the manipulations and procedures done with the animals, these were conducted following the guidelines of the Official Mexican Standard NOM-062-ZOO-1999.
A detailed procedure to administrate the chow diet, HFD, and the standardization of the F0 mother’s phenotype to produce the F1 offspring has been described by our group. Reference De los Santos, Coral-Vázquez and Menjivar9 Furthermore, a prior publication also outlined the procedure for administering the flavonoid (−)-epicatechin. Reference De los Santos, Coral-Vázquez and Menjivar9
Briefly, female albino Wistar rats bred in the animal care center of the INCMNSZ were maintained at 22–23°C under controlled lighting (lights on 07:00–19:00 h) and were fed with Chow diet (Purina 5001, Labdiet, St Louis, MO, U.S.A.). At the age of 14–16 weeks, when they weighed 200–240 g, females were randomly bred with males from other mothers.
The study was performed in two stages. The first stage was completed at weaning (day 21), in which one female, F0 pup from each litter, was randomly assigned to either a control (C, n = 6) group, that received the laboratory Chow diet, or to a maternal obesity group (MO, n = 6) group fed a HFD with an energy content of 4.9 kcal/g. A total of four rats for each group were included.
The six F0 female albino Wistar rats were placed with proven male breeders on postnatal day 120 and conceived during the next cycle. To minimize consumption of the HFD, by males, during the mating period, males were placed with females at night and removed during the morning. Lactating mothers were maintained on their pregnancy diet (Figure 1). Both diets were adjusted and administrated following the recommendations to ensure adequate support for growth, pregnancy, and lactation phases outlined by the American Institute of Nutrition (AIN-93G). Reference Reeves, Nielsen and Fahey18
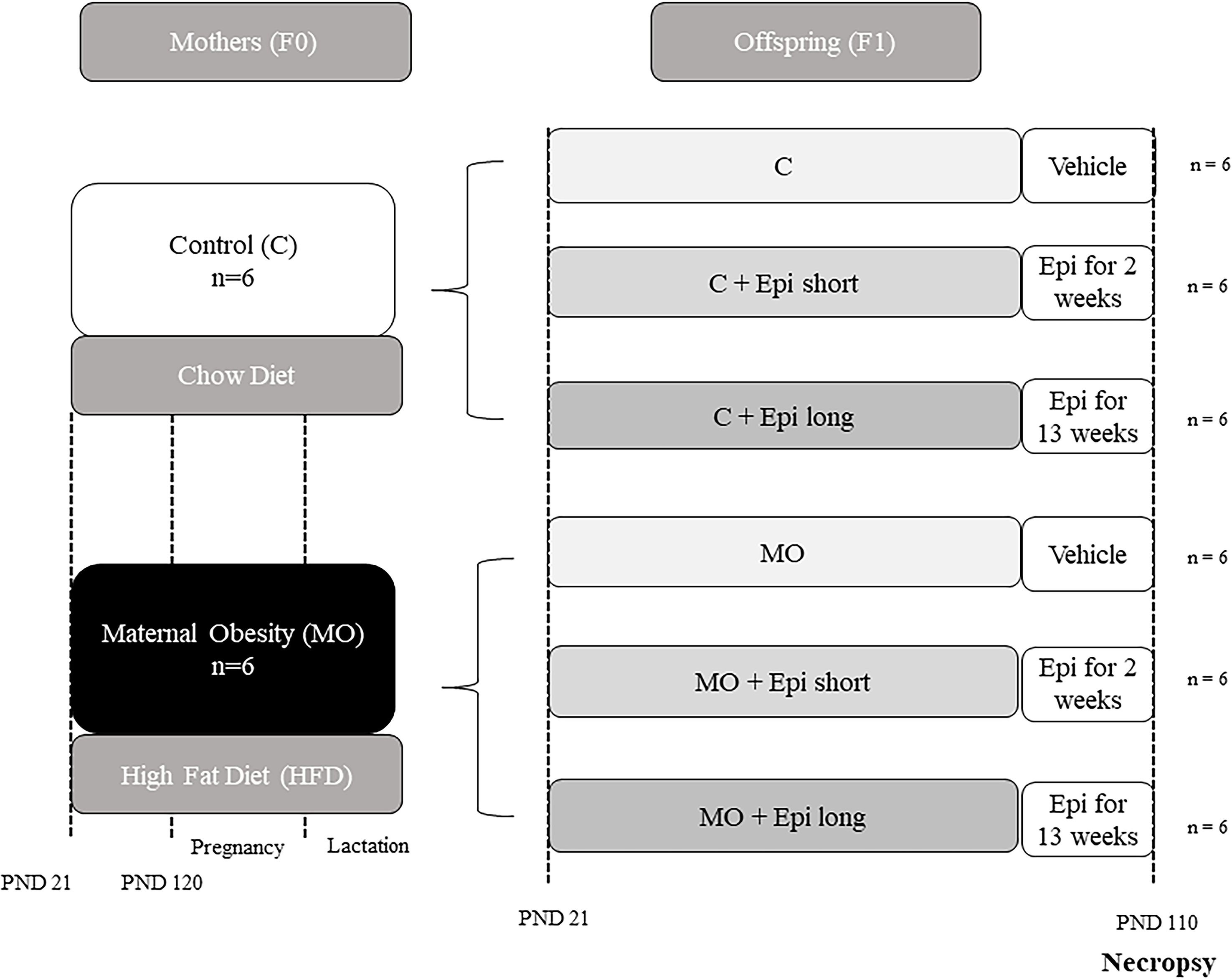
Figure 1. Timeline for the study of two interventions with Epi (short and long), in male offspring of obese mothers at 110 postnatal days (F1). Obesity model. Four female rats (F0) per group were fed with control or high-fat diet at weaning and during pregnancy and lactation. At postnatal day 21, twelve males per group (C and MO) from different litters were randomly allocated to be treated with water or with (−)-epicatechin twice per day for 2 or 13 weeks (C, C + Epi short, C + Epi long, MO, MO + Epi short, MO + Epi long, n = 6 rats per group).
For the stage two, F1 offspring, litter size and pup weight were recorded at birth. The anogenital distance was measured to identify males and females. To guarantee the homogeneity of all litters of F1 offspring on postnatal day 2, all litters studied were adjusted to 10 pups with equal numbers of males and females. Offspring were weaned at postnatal day 21, housed two rats per cage, and fed chow diet throughout the study. Only male offspring were studied. Three male offspring from C or MO mothers from different litters were randomly selected to the control group (vehicle) or to the (−)-epicatechin intervention groups (short or long) (Figure 1).
For the (−)-epicatechin treatment, two distinct protocols were implemented: one from day 21 to day 110 postnatal (lasting 13 weeks, referred to as the long protocol), Reference Cheng, Xu and Zhao19 and the other from day 95 to day 110 postnatal (comprising two weeks, referred to as the short protocol). Reference Ramirez-Sanchez, De los Santos and Gonzalez-Basurto20,Reference De Los Santos, García-Pérez and Hernández-Reséndiz21 The male pups were either administered a vehicle (water) or 1 mg/kg of body weight of (−)-epicatechin (Sigma-Aldrich, St Louis, MO, U.S.A.) via oral catheter twice daily (Figure 1). Reference Gutierrez-Salmean, Ciaraldi and Nogueira14,Reference Ramirez-Sanchez, De los Santos and Gonzalez-Basurto20,Reference De Los Santos, García-Pérez and Hernández-Reséndiz21
To form the experimental groups, six male offspring from different litters from both the F0 groups (originating from either control mothers (C) or obese mothers (MO) were randomly allocated to one of the six experimental groups, which were as follows: offspring F1 of control mother (C)), offspring F1 of control mother and two weeks of postnatal (−)-epicatechin treatment (C + Epi short), offspring F1 of control mother and 13 weeks of postnatal (−)-epicatechin treatment (C + Epi long), offspring F1 of obese mother (MO), offspring F1 of obese mother and two weeks of postnatal (−)-epicatechin treatment (MO + Epi short), and offspring F1 of obese mother and 13 weeks of postnatal (−)-epicatechin treatment (MO + Epi long) (total of male F1 rat = 32) (Figure 1). It’s important to note that no adverse effects resulting from this dietary intervention were recorded, and no rats perished before the conclusion of the experimental protocol.
Gastrocnemius tissue RNA extraction
On postnatal day 110, the rats were euthanized, and skeletal muscle tissue (gastrocnemius) was collected from each animal. RNA extraction from the gastrocnemius muscle tissue samples was performed using TRIzol reagent (Invitrogen®, Carlsbad, CA, U.S.A.), following the manufacturer´s instructions.
Gene expression analysis
RNA (100 ng) was used for the evaluation of gene expression using the AgPath-ID™ One-Step RT-PCR Reagents (Applied Biosystems™, Carlsbad, CA, U.S.A.), following instructions provided by the manufacturer. This procedure allowed for the assessment of the gene expression of Muscle RING-finger protein-1 (Murf1), Muscle atrophy F-box (MAFbx), Forkhead box protein O1 (Foxo1), Nuclear factor kappa-light-chain-enhancer of activated B cells (NFkB), and Ribosomal protein S6 kinase beta-1 (p70S6K-alpha) in gastrocnemius tissue, only in C, MO, C + Epi long and MO + Epi long groups using reverse transcription-quantitative polymerase chain reaction (RT-qPCR). TaqMan on Demand Gene Expression Probes from Applied Biosystem (Applied Biosystems™, Carlsbad, CA, U.S.A.), were employed for this analysis: assay ID: Murf1 (Rn00590197_m1), MAFbx (Rn00591730_m1), Foxo1 (Rn01494868_m1), NFκB (Rn01502266_m1) and p70 S6K-alpha (Rn00583148_m1). The values were normalized to the relative amounts of β-actin (Assay ID: Rn00667869) for each sample, allowing for comparing and interpreting gene expression levels in the different experimental groups.
Reverse transcription-quantitative PCR (RT-qPCR) was carried out using a LightCycler® 480 Instrument (Roche Diagnostics Ltd, Risch-Rotkreuz, Switzerland). The data obtained were normalized using an endogenous control, β-actin, and relative quantification was determined using the 2^(-Delta Delta CT) procedure.
Western blotting
Protein expression was analyzed across all experimental groups, which included C, MO, C + Epi short, C + Epi long, MO + Epi short, and MO + Epi long. Fifty mg of protein extracts of gastrocnemius, obtained by homogenization in RIPA lysis buffer (Santa Cruz Biotechnology, Inc., Dallas, TX, U.S.A.) and supplemented with protease and phosphatase inhibitors, were submitted to SDS-PAGE gel electrophoresis. They were subsequently transferred to a nitrocellulose membrane 0.45 µm (Bio-Rad Laboratories, Life Science Group, Hercules, CA, U.S.A). The proteins were detected using the following primary antibodies: recombinant Anti-MURF1 + MURF3 + MURF2 (ab172479) (Abcam Inc., Waltham, MA, U.S.A.); recombinant Anti-Fbx32 (ab168372) (Abcam Inc., Waltham, MA, U.S.A.); anti-Foxo1 (C29H4, Rabbit mAb#2880), (CellSignaling, Danvers, MA, U.S.A.); anti-Phospho-Foxo1 (Ser256) (E1F7T, Rabbit mAb #84192), (CellSignaling, Danvers, MA, U.S.A.); anti-NF-κB p65 (C22B4, Rabbit mAb #4764), (CellSignaling, Danvers, MA, U.S.A.); anti-Phospho-NF-κB p65 (Ser536) (93H1, Rabbit mAb#3033), (CellSignaling, Danvers, MA, U.S.A.); anti-p70S6 Kinase (49D7, Rabbit mAb #2708), (CellSignaling, Danvers, MA, U.S.A.); anti-Phospho-p70 S6 Kinase (Thr389) (108D2, Rabbit mAb #9234), (CellSignaling, Danvers, MA, U.S.A.). Anti-Gadph (PA1-988), (ThermoFisher Scientific, Rockford, IL, U.S.A.) was used as a loading control. Protein detection was carried out using the appropriate secondary antibody: HRP-goat anti-rabbit IgG (111-095-003) or H.R.P.- anti-mouse IgG (715-035-150) (Jackson Immuno Research Laboratories, Inc., West Grove, PA, U.S.A.). The proteins were detected by chemiluminescence with the SuperSignal™ West Femto Chemiluminescent Substrate kit (ThermoFisher Scientific, Rockford, IL, U.S.A.). Digital image adquisition was obtained with C-DiGit Blot Scanner (LI-COR Biosciences, Lincoln, NE, U.S.A.) using LI-COR Image Studio software (http://www.licor.com/bio/products/software/image_studio_lite/); additionally, ImageJ software (U.S. National Institutes of Health, Bethesda, Maryland, USA), was employed for further analysis and quantification of the protein band intensities.
Statistical analysis
The data were tested to determine normal distribution using the Kolmogorov-Smirnov test. Data are expressed as mean ± standard deviation of 6 individual experimental observations for gene expression. A two-way ANOVA was performed to determine the influence of the two variables over the model (maternal diet and Epi treatment) and possible interactions between them. To address the effect size, percentage of variation of the model attributable to each variable was calculated using the omega-square measure, and the ones with significant were reported. It was followed by a one-way ANOVA and a Tukey post hoc test to determine differences between individual groups.
For protein expression, data is presented as median ± range of 6 individual experimental observations; a Kruskal-Wallis non-parametric test was then conducted, followed by a Dunn’s post hoc test. Data were analyzed with GraphPad Prism 6.0 software (GraphPad Software, San Diego, CA). Significant differences were defined by a p < 0.05.
Sample size and power were calculated according to the algorithm of the percentage of success and error test (by group/by trial): n = log B/log p; n = 6.39 per group/test.
Results
Effect of (−)-epicatechin long treatment in the expression of mRNA of genes related to muscle atrophy or protein synthesis in F1 male offspring of obese wistar rats
The relative expression for the genes of interest was performed with qRT-PCR, using β-actin as a reference gene. All measurements were taken as duplicates, and the 2−ΔΔCt method was applied for data analysis. After the analysis by two-way ANOVA, we found an influence of the Epi long treatment over the model by decreasing the Murf1 gene expression in both groups treated with the flavonoid (C + Epi long and MO + Epi long) (p = 0.036) accounting for 13.08% of the variation of the model (Figure 2, Panel A). Moreover, we also found an influence of the Epi long treatment over the NFκB expression by decreasing the fold increase in both groups treated with the flavonoid (C + Epi long and MO + Epi long) (p = 0.038) accounting for 15.03% of the variation of the model (Figure 2, Panel C); however, we did not find any interaction between the variables.
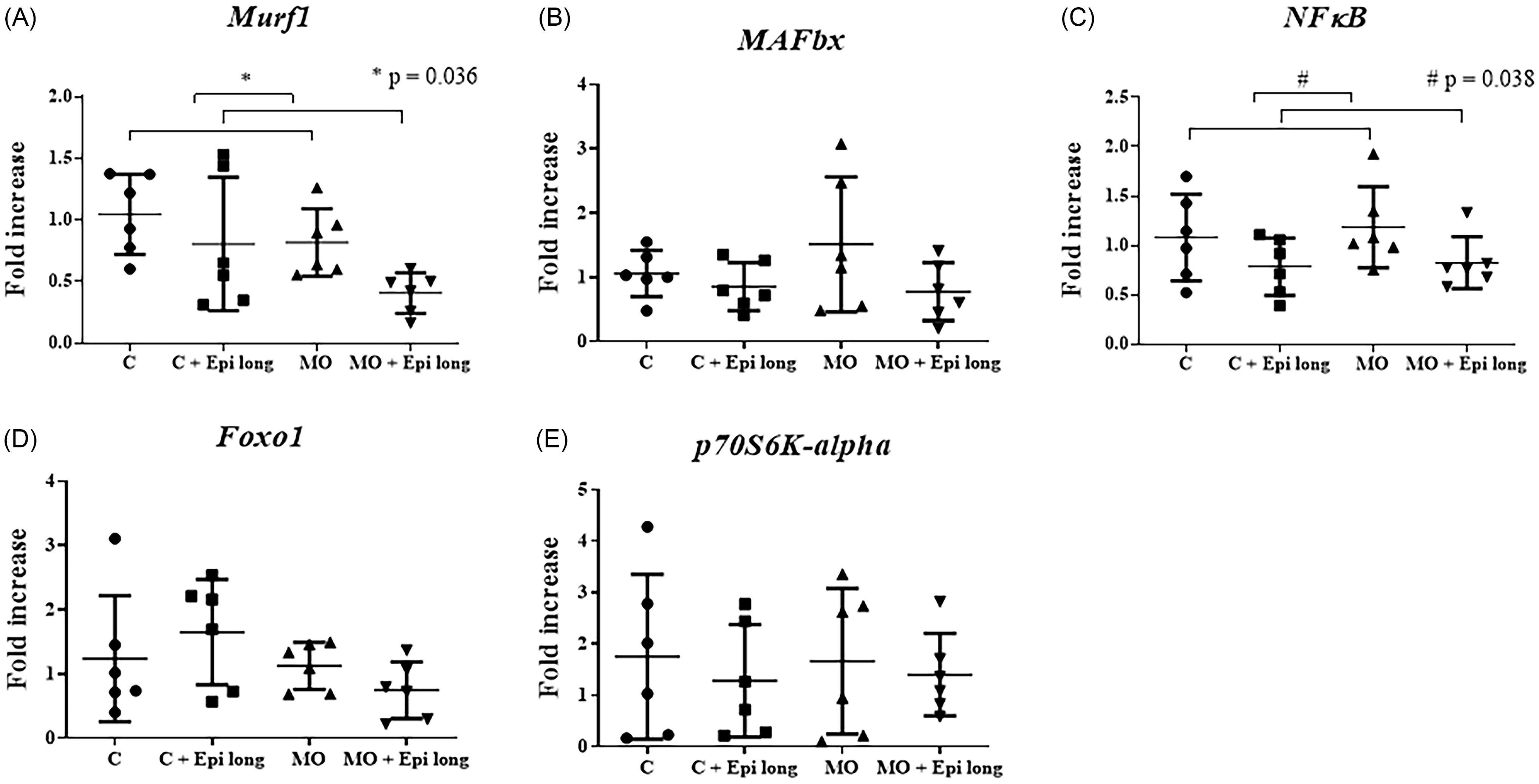
Figure 2. Effect of long-treatment with Epi on relative mRNA expression of genes related to atrophy and protein synthesis in the gastrocnemius muscle, of male offspring of obese mothers at 110 postnatal days. ( a ) Murf1, ( b ) MAFbx, ( c ) NFkB, ( d ) Foxo1, and ( e ) p70S6K-alpha mRNA expression. Epi long treatment decreasing the Murf1 gene expression in C + Epi long and MO + Epi long groups (p = 0.036) (Figure 2, Panel A). Also, Epi long treatment decreased the NFκB expression, by the fold increase in C + Epi long and MO + Epi long groups (p = 0.038) (Figure 2, Panel C). Data are expressed as median and range and were analyzed by Kruskal-Wallis followed by post hoc Dunn test for pair comparison (n = 6 rats per group). C = male rats offspring of control mothers; C + Epi long= male rats offspring of control mothers treated with Epi 13 weeks; MO = male rats descended from obese mothers; MO + Epi long= male rats descended from obese mothers treated with Epi 13 weeks.
MAFbx, Foxo1, and p70S6K-alpha mRNA expression showed no changes secondary to maternal diet or Epi treatment in any experimental group (C, C + Epi long, MO and MO + Epi long).
Afterward, we performed a one-way ANOVA, followed by the post hoc Tukey test for pair comparison (n = 6 rats per group), finding no differences in Murf1, MAFbx, NFκB, Foxo1, or p70S6K-alpha mRNA expression among offspring of C, MO, C + Epi long and MO + Epi long groups (Figure 2, Panel A-to-E).
Effect of (−)-epicatechin long and short treatment in the expression of proteins related to muscle atrophy or synthesis of proteins in F1 male offspring of obese wistar rats
Western blot assays did not show changes in the level of Murf1 and MAFbx proteins related to muscle protein degradation through the proteasome, both in the control group (C) and in the offspring of obese mothers (MO). Likewise, administration of the flavonoid did not modify the expression of these proteins, neither with the treatment of 2 weeks (short protocol), nor with the treatment of 13 weeks (long protocol) (Figure 3, Panel A-to-D).
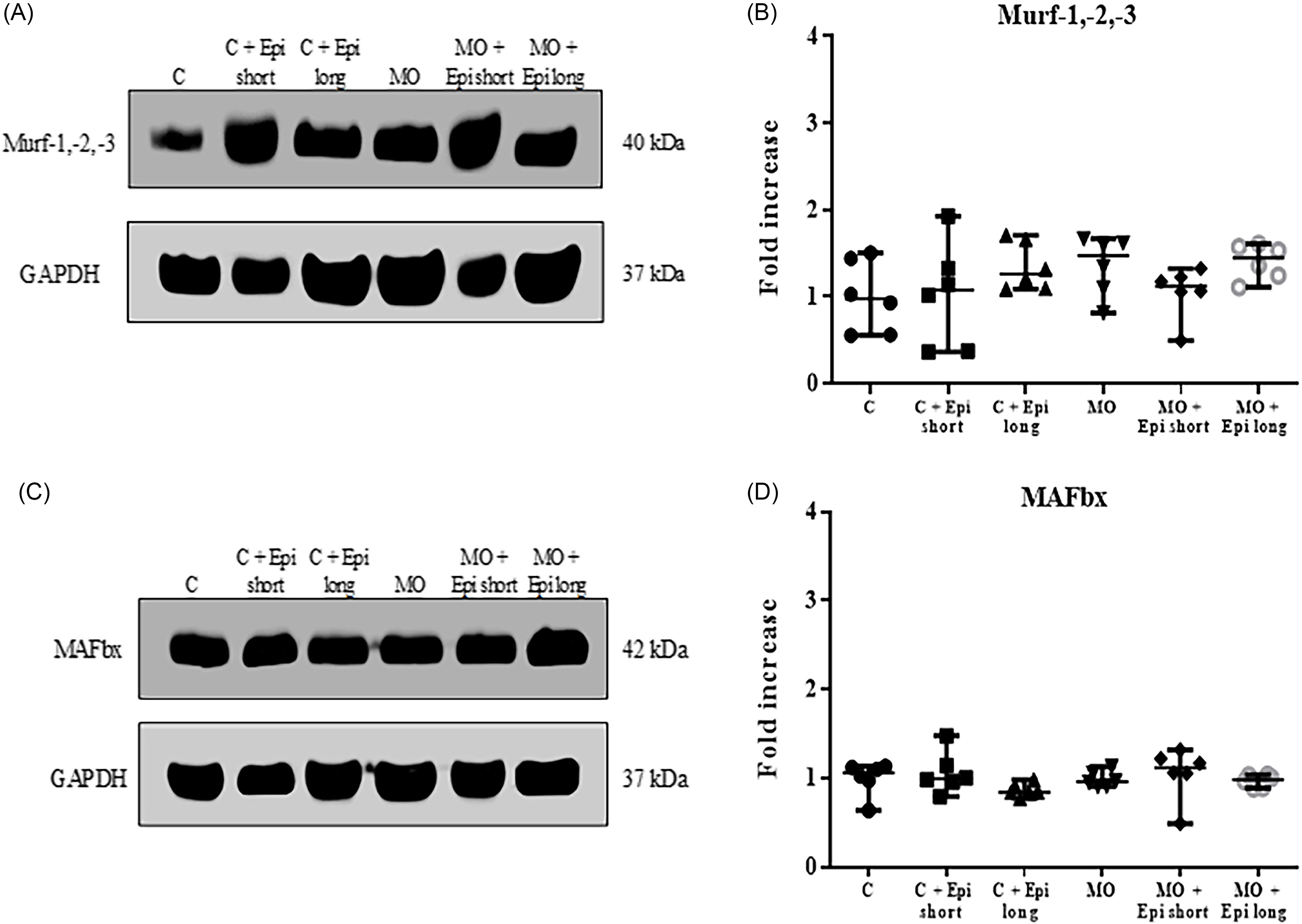
Figure 3. Effect of short-and-long-treatment with Epi on proteins related to atrophy in the gastrocnemius muscle of male offspring of obese mothers at 110 postnatal days. ( a ) and ( c ) Representative immunoblotting of protein of Murf1 and MAFbx, GAPDH was used as a loading control. ( b ) and ( d ) densitometry analysis of Murf1 and MAFbx protein expression in gastrocnemius muscle. Data are expressed as median and range and were analyzed by Kruskal-Wallis followed by post hoc Dunn test for pair comparison (n = 6 rats per group). C = male rats offspring of control mothers; C + Epi short= male rats offspring of control mothers treated with Epi 2 weeks; C + Epi long= male rats offspring of control mothers treated with Epi 13 weeks; MO = male rats descended from obese mothers; MO + Epi short= male rats descended from obese mothers treated with Epi 2 weeks; MO + Epi long= male rats descended from obese mothers treated with Epi 13 weeks.
On the other hand, a significant reduction of the total NFκB protein was observed in the MO + Epi long treated animals in comparison to the offspring of the C + Epi long (p < 0.05) and a no effect in the expression of NFκB total levels in the descendants of the C + Epi long versus C group (p = 0.06) (Figure 4, Panel A-to-C); in contrast, no significant differences were determined in the phosphorylated/total protein ratio in any of the experimental groups (Figure 4, Panel A-to-C).
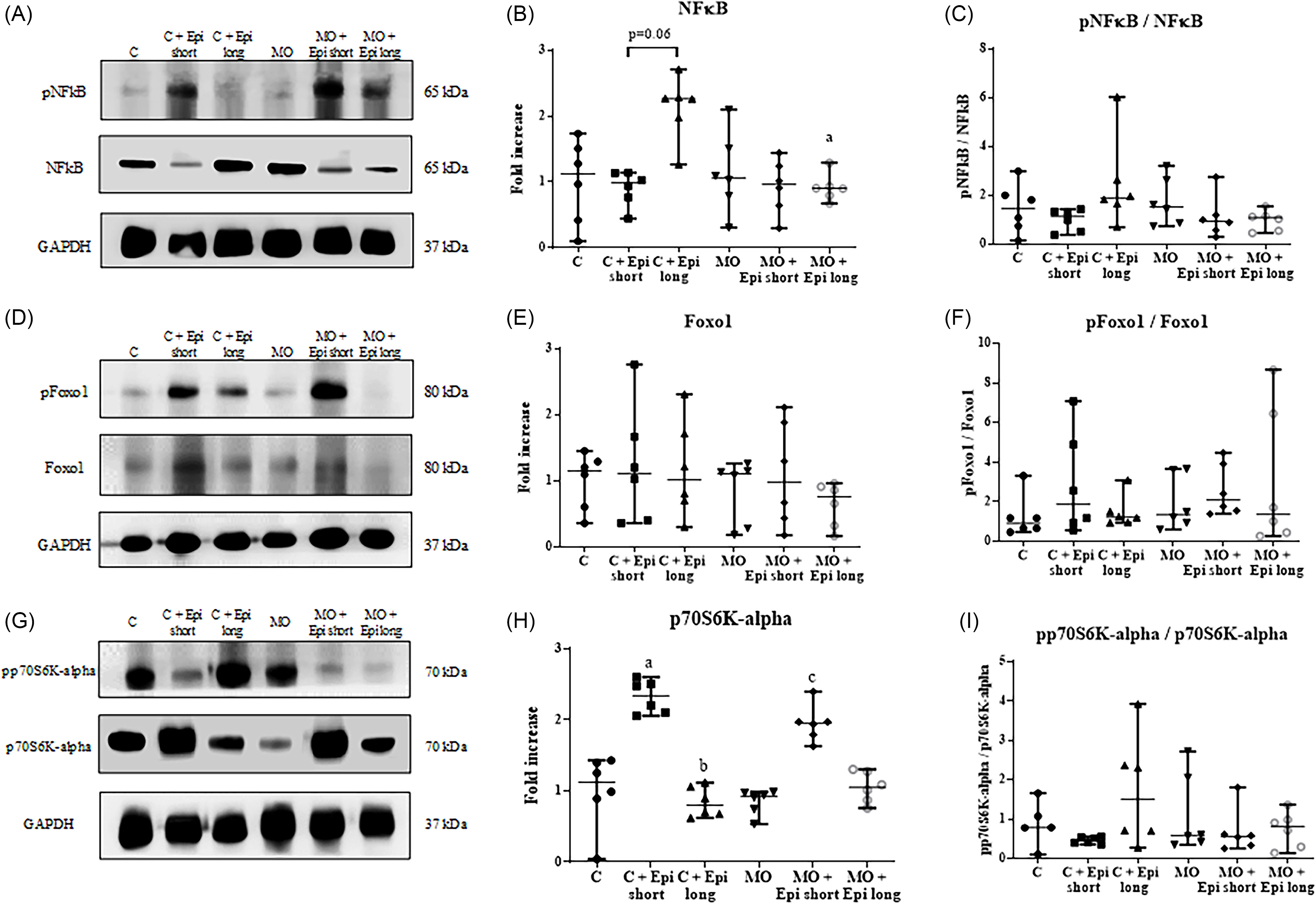
Figure 4. Effect of short-and-long-treatment with Epi on proteins related to atrophy and synthesis of proteins in the gastrocnemius muscle of male offspring of obese mothers at 110 postnatal days. (a), ( d ), and ( g ) Representative immunoblotting of NFκB total and phosphorylated protein, Foxo1 total and phosphorylated protein, and p70 S6K-alpha total and phosphorylated protein. GAPDH was used as a loading control. Densitometry analysis of protein expression ( b ) and ( c ) NFkB total and the relationship between phosphorylated and total NFkB, ( e ) and ( f ) Foxo1 total and the relationship between phosphorylated and total Foxo1, and ( h ) and ( i ) p70 S6K-alpha total and the relationship between phosphorylated and total p70 S6K-alpha in gastrocnemius muscle. ( b ) C + Epi long show a higher NFkB total protein level in comparison to offspring of MO + Epi long. ( h ) The C + Epi short group presented a significant increase in the total expression of p70S6K-alpha compared to the C group; the C + Epi long group showed a significant decrease in the total level of this protein versus the C + Epi short group; besides, the p70S6K-alpha total level significantly increased in the MO + Epi short group in comparison to the MO group. Data are expressed as median and range and were analyzed by Kruskal-Wallis followed with post hoc Dunn test for pair comparison. a p < 0.5 vs. C + Epi long and C group; b p < 0.01 vs. C + Epi short group; c p < 0.05 vs. MO group (n = 6 rats per group). C = male rats offspring of control mothers; C + Epi short= male rats offspring of control mothers treated with Epi 2 weeks; C + Epi long= male rats offspring of control mothers treated with Epi 13 weeks; MO = male rats descended from obese mothers; MO + Epi short= male rats descended from obese mothers treated with Epi 2 weeks; MO + Epi long= male rats descended from obese mothers treated with Epi 13 weeks.
Moreover, our results of the expression of the transcription factor Foxo1 showed that the offspring of the C, MO, Epi short- or long-treatment groups did not change this protein’s total or phosphorylation levels in any of the analyzed groups (Figure 4, Panel D-to-F).
Regarding the expression of kinase p70S6K-alpha, we observed that in the control group, with a short treatment of Epi (C + Epi short), a significant increase in the total expression of p70 S6K-alpha was observed when compared to the control group (C) (p < 0.05), but the long treatment of Epi (C + Epi long) showed a significant decrease in the total level of this protein versus the C + Epi short group (p < 0.01). Likewise, we observed that the p70S6K-alpha total levels increased significantly in the MO + Epi short group compared to the MO group (p < 0.05). Nevertheless, all analyzed groups showed significant changes in the proportion of phosphorylated and total protein (C, MO, C + Epi short, C + Epi long, MO + Epi short, and MO + Epi long) (Figure 4, Panel G-to-I).
Discussion
Obesity favors the development of musculoskeletal disorders that include muscle atrophy due to the degradation of muscle fibers, reduction in the size of myofibers, and decrease in the number of satellite cells, in addition to the reduction of type 1 oxidative fiber and an increase in type 1 glycolytic fibers 2X. Reference Blaauw, Schiaffino and Reggiani22–Reference Collins, Herzog and MacDonald24
Besides, in the context of programing, obesity induces in the skeletal muscle of puppies of mothers fed with a HFD during gestation and lactation, a lower percentage of lean mass, a decrease in protein content, an increase in intramuscular inflammation, imbalance of protein synthesis/degradation markers, as well as deregulation in myogenesis. Reference De los Santos, Coral-Vázquez and Menjivar9,Reference Tong, Yan, Zhu, Ford, Nathanielsz and Du25–Reference Cui, Song and Wang27
Given the acute effects that obesity has on offspring’s skeletal muscle programing, it is essential to explore preventive strategies based on natural bioactive compounds such as the flavonoid (−)-epicatechin. In this regard, Epi has been shown to benefit the skeletal muscles of murine models with sarcopenia, murine models and patients with muscular dystrophy, and the skeletal muscles of patients with diabetes. Reference Ramirez-Sanchez, De los Santos and Gonzalez-Basurto20,Reference Taub, Ramirez-Sanchez and Ciaraldi28–Reference Ramírez-Ramírez, Fernández-Valverde and Reséndiz-García30 Likewise, treatment with this flavonoid increased the percentage of lean tissue in the offspring of mothers fed with a HFD. Reference De los Santos, Coral-Vázquez and Menjivar9
We investigated whether the offspring of mothers with obesity, induced by a HFD, presented disorders in the expression of genes related to atrophy (Murf1, MAFbx, NFκB and Foxo1) or protein synthesis in the musculoskeletal tissue (p70S6K-alpha) Reference Sishi, Loos, Ellis, Smith, Du Toit and Engelbrecht5 and if these disorders were modified with the administration of two Epi protocols (short and long).
The high plasticity of skeletal muscle is carried out by regulating anabolic and catabolic processes, which can be disturbed by various diseases, including obesity and metabolic disorders. Sishi et al. Reference Sishi, Loos, Ellis, Smith, Du Toit and Engelbrecht5 demonstrated that obesity increases the expression of proteins related to atrophy, with the ubiquitin-proteasome proteolytic pathway being one of those responsible for regulating the degradation of skeletal muscle proteins through the Murf1 and MAFbx ligases, the activation of the Foxo1 transcription factor or through signaling from NFkB; both pathways, lead to muscle breakdown known as “ubiquitin-dependent protein wastage.” Reference Sishi, Loos, Ellis, Smith, Du Toit and Engelbrecht5,Reference Granado, Martín, Priego, López-Calderón and Villanúa31
Moreover, phosphorylation of the kinase p70S 6K-alpha by the Pi3K/Akt/Mtor axis has been reported as necessary for muscle fibers in order to reach their standard size by increasing protein synthesis. Reference Ohanna, Sobering and Lapointe32
To the best of our knowledge, most reports in the literature are directed at the analysis of genes related to atrophy or protein synthesis in a direct model of obesity, Reference Sishi, Loos, Ellis, Smith, Du Toit and Engelbrecht5,Reference Sun, Huang and Chen33,Reference Cheng, Lin and Liao34 and studies related to the impact of obesity by programing on skeletal muscle, are aimed at analyzing other molecular pathways in skeletal muscle. Reference Bayol, Simbi and Stickland8,Reference Tong, Yan, Zhu, Ford, Nathanielsz and Du25,Reference Cui, Song and Wang27
It has been described that in the gastrocnemius muscle of adult male rats (150 days postnatal), descendants of obese mothers nurtured with a HFD, have a diminished content of muscle proteins and signs of skeletal muscle atrophy, with overexpression of Murf1 and increased phosphorylated NFkB; whereas, the Foxo1 and p70S6K-alpha phosphorylation at Thr389 showed no significant differences in this tissue. These authors proposed that MO negatively impacts skeletal muscle development in the offspring. Reference Pileggi, Segovia and Markworth26
Regarding our results, in contrast with those reported by Pileggi et al., Reference Pileggi, Segovia and Markworth26 we did not find significant differences in the expression of Murf1 and MAFbx mRNA, as well as in the phosphorylated NFkB and p70S6K levels in the gastrocnemius muscle, in the offspring obese by programing in comparison to the control group. Interestingly, Epi administration modified the Murf1 and NFkB mRNA levels but not at the protein levels. This discrepancy between the mRNA expression and the protein levels, could be due to transcriptional, post-transcriptional, translational, and post-translational regulation Reference Schwanhäusser, Busse and Li35 as well as by protein stability, Reference Geiger, Burkhart, Gambaryan, Walter, Sickmann and Zahedi36 or we hypothesize that because we studied young rats, the effect of MO on the muscle tissue of their offspring, first manifests itself in variations in the expression of mRNA, and could being the starting point for the subsequent change in the expression of their proteins in older ages.
Furthermore, in spite that we found that Epi treatment was able to modify the total NFkB and p70 S6K-alpha protein levels, no significant differences were determined in the phosphorylated/total protein ratio in any of the experimental groups, indicating that the administration of this flavonoid could exert an effect on the expression of the total protein, but not on its phosphorylation pattern.
The discrepancies between our results and those reported by Pileggi et al. Reference Pileggi, Segovia and Markworth26 might be due to the age of the animals; in our study, 110 days of postnatal life, equivalent to a young adult (in contrast to rats of 150 days that are mature adults), which could be related to the lack of disorders at this age, without discarding the possibility of presenting them later.
Also, several studies have demonstrated that type II muscular fibers are smaller in the aged population and type I are more conserved, Reference Lexell37,Reference Verdijk, Koopman, Schaart, Meijer, Savelberg and van Loon38 showing differences in the composition of skeletal muscle depending on age. These different types of muscle fibers elicit responses to neural inputs and metabolic stressors, Reference Schiaffino and Reggiani39,Reference Talbot and Maves40 including obesity, showing a shift in the proportion of fibers towards more type IIb fibers. In contrast, the amount of type I and IIa fibers is diminished. Reference Tanner, Barakat and Lynis Dohm41,Reference Maltin42 Considering that animals analyzed in the present study are equivalent to young adults, having a higher proportion of type II fibers, potential disorders that the gastrocnemius muscle may present could still be compensated thanks to the abundance of these fibers.
In conclusion, our results show that male offspring at 110 postnatal days, descendants of obese mothers, do not present disorders in the expression at RNA level or proteins of the atrophy pathway, nor in the protein synthesis in the gastrocnemius muscle. However, treatment with a long protocol of (−)-epicatechin reduces the mRNA of the muscle atrophy genes Murf 1 and NFkB, but these changes are not maintained at the protein level. Further, Epi could not modify the phosphorylated/total protein ratio of the NFkB and p70S6K-alpha. Finally, MAFbx and Foxo1 remain unchanged either at mRNA or protein level by the Epi administration.
Further, our results suggest that the age of the animals could be a determining factor in the effect of MO on musculoskeletal tissue, showing slight changes at this early age; however, research must be extended to analyze these changes over time. On the other hand, Epi administration could affect MO and fetal programing, showing certain effects on proteins related to the balance of anabolic/catabolic pathways in musculoskeletal tissue. For this reason, it would be relevant in future research to evaluate the effect of Epi on older animals and/or with another stimulus, such as a postnatal diet rich in fat. Likewise, these studies on muscle fibers with different glycolytic or oxidative capacities would be important.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S2040174424000187.
Data availability statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
This work was partially supported by the Fondo Sectorial de Investigación para la Educación SEP-CONACYT, Convocatoria de Investigación Científica Básica 2017-2018, CONACyT México, under Grant: A1-S-9740, and by the Newton Fund RCUK- CONACYT-2015 (Research Conuncils UK – CONACyT).
This work was submitted in partial fulfillment of the requirements for the Ph.D. degree of Ana Luisa Álvarez Chávez at the Programa de Doctorado en Ciencias Bioquímicas, Universidad Nacional Autónoma de México.
Ana Luisa Álvarez Chávez received the Apoyo para Ayudantes de Investigador Nivel III o Emérito del Sistema Nacional de Investigadores (S.N.I.), Consejo Nacional de Ciencia y Tecnología (CONACyT), México, fellowship award.
Sergio De los Santos received a postdoctoral fellowship award from the Programa de Becas Posdoctorales de la Dirección General de Asuntos del Personal Académico, División de Investigación, Facultad de Medicina, Universidad Nacional Autónoma de México.
We thank Sebastián De la Rosa, María Elena Tejeda, and Jorge Uribe, from the Unidad de Investigación en Obesidad, Facultad de Medicina, Universidad Nacional Autónoma de México (S.D.R. and M.E.T) and from the Departamento de Biología de Reproducción, Instituto Nacional de Ciencias Médicas y Nutrición “Salvador Zubirán” (J.U.), for their technical assistance in this study.
Author contribution
Study Design: Ana Álvarez-Chávez, Sergio de los Santos, Ramón Mauricio Coral-Vázquez, Elena Zambrano, and Patricia Canto.
Data Collection: Ana Álvarez-Chávez, and Sergio de los Santos.
Statistical Analysis: Ana Álvarez-Chávez, Ramón Mauricio Coral-Vázquez, and Patricia Canto.
Data Interpretation: Ana Álvarez-Chávez, Sergio de los Santos, Ramón Mauricio Coral-Vázquez, Juan Pablo Méndez, and Elena Zambrano, and Patricia Canto.
Manuscript Preparation: Ana Álvarez-Chávez, Juan Pablo Méndez, Ramón Mauricio Coral-Vázquez, and Patricia Canto
Literature Search: Ana Álvarez-Chávez, and Patricia Canto.
Funds Collection: Juan Pablo Méndez, Elena Zambrano, and Patricia Canto.
All authors have approved the final article.
Competing interests
None.
Ethical standard
The research protocol in rodents followed in the present study was approved by the Ethics Committee in Animal Experimentation of the Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán (INCMNSZ) (CINVA: UIO-1892-17/19-1, approved August 27, 2017). In addition, all procedures followed the Guidelines for the Care and Use of Laboratory Animals of the Institute of Laboratory Animal Resources (http://www.nal.usda.gov/awic/animal-welfare-act).






