


No CrossRef data available.
Article contents
Crowding accelerates the rotation of a bacterial rotor
Published online by Cambridge University Press: 18 September 2024
Abstract
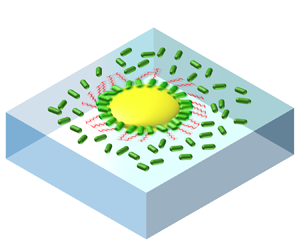
Understanding the propulsion of a swimmer in a large group of individuals holds the key to unravelling the intriguing dynamics of active matter collective motion. Here, we develop a two-dimensional (2-D) self-assembled rotor, powered by bacterial flagella. At a water–air interface, the average direction of rotation of a rotor is fixed. When the chiral rotor is put into a 2-D bacterial suspension, we examine the average and fluctuation of the angular velocity of the rotor. Remarkably, the average angular velocity of a rotor is found to increase up to 3 times when the density of surrounding bacterial suspension increases and the increase is nonlinear. In a dense suspension of bacteria, the existence of a rotor disrupts vortices in the surrounding active turbulence, and the acceleration of the rotor is independent of the activity level of the surrounding free bacteria. The nonlinear acceleration thus results from hydrodynamic interaction with surrounding crowdedness that can be quantitatively explained by hydrodynamic simulation. The simultaneity between the acceleration of rotor and free bacteria in active turbulence suggests that crowding-induced acceleration may promote the onset of instability. The result will inspire new active-matter-based microfluidic devices with improved transport properties.
JFM classification
Information
- Type
- JFM Papers
- Information
- Copyright
- © The Author(s), 2024. Published by Cambridge University Press
References
Aranson, I.S., Sokolov, A., Kessler, J.O. & Goldstein, R.E. 2007 Model for dynamical coherence in thin films of self-propelled microorganisms. Phys. Rev. E 75, 040901.CrossRefGoogle ScholarPubMed
Beppu, K., Izri, Z., Sato, T., Yamanishi, Y., Sumino, Y. & Maeda, Y.T. 2021 Edge current and pairing order transition in chiral bacterial vortices. Proc. Natl Acad. Sci. USA 118, e2107461118.CrossRefGoogle ScholarPubMed
Bianchi, S., Saglimbeni, F., Frangipane, G. & Di Leonardo, R. 2022 Flagellar elasticity and the multiple swimming modes of interfacial bacteria. Phys. Rev. Res. 4, L022044.CrossRefGoogle Scholar
Cisneros, L.H., Kessler, J.O., Ganguly, S. & Goldstein, R.E. 2011 Dynamics of swimming bacteria: transition to directional order at high concentration. Phys. Rev. E 83, 061907.CrossRefGoogle ScholarPubMed
Cortez, R., Fauci, L. & Medovikov, A. 2005 The method of regularized Stokeslets in three dimensions: analysis, validation, and application to helical swimming. Phys. Fluids 17, 031504.CrossRefGoogle Scholar
Darnton, N., Turner, L., Breuer, K. & Berg, H.C. 2004 Moving fluid with bacterial carpets. Biophys. J. 86, 1863–1870.CrossRefGoogle ScholarPubMed
Dewangan, N.K. & Conrad, J.C. 2019 Rotating oil droplets driven by motile bacteria at interfaces. Soft Matt. 15, 9368-9375.CrossRefGoogle ScholarPubMed
Di Leonardo, R., Angelani, L., Arciprete, D.D., Ruocco, G., Iebba, V., Schippa, S., Conte, M.P., Mecarini, F., De Angelis, F. & Di Fabrizio, E. 2010 Bacterial ratchet motors. Proc. Natl Acad. Sci. USA 107, 9541–9545.CrossRefGoogle ScholarPubMed
Drescher, K., Dunkel, J., Cisneros, L.H., Ganguly, S. & Goldstein, R.E. 2011 Fluid dynamics and noise in bacterialcell-cell and cell-surface scattering. Proc. Natl Acad. Sci. USA 108, 10940–10945.CrossRefGoogle ScholarPubMed
Dunkel, J., Heidenreich, S., Drescher, K., Wensink, H.H., Bär, M. & Goldstein, R.E. 2013 Fluid dynamics of bacterial turbulence. Phys. Rev. Lett. 110, 228102.CrossRefGoogle ScholarPubMed
Giomi, L. 2015 Geometry and topology of turbulence in active nematics. Phys. Rev. X 5, 031003.Google Scholar
Guo, S., Samanta, D., Peng, Y., Xu, X. & Cheng, X. 2018 Symmetric shear banding and swarming vortices in bacterial superfluids. Proc. Natl Acad. Sci. USA 115, 7212-7217.CrossRefGoogle ScholarPubMed
Hatwalne, Y, Ramaswamy, S., Rao, M. & Simha, R.A. 2004 Rheology of active-particle suspensions. Phys. Rev. Lett. 92, 1118101.CrossRefGoogle ScholarPubMed
Happel, J. & Brenner, H. 1983 Low Reynolds Number Hydrodynamics: With Special Applications to Particulate Media. Springer Science & Business Media.CrossRefGoogle Scholar
Hernández-Ortiz, J.P., de Pablo, J.J. & Graham, M.D. 2007 Fast computation of many-particle hydrodynamic and electrostatic interactions in a confined geometry. Phys. Rev. Lett. 98, 140602.CrossRefGoogle Scholar
Koch, D.L. & Subramanian, G. 2011 Collective hydrodynamics of swimming microorganisms: living fluids. Annu. Rev. Fluid Mech. 43, 637–659.CrossRefGoogle Scholar
Laganenka, L., Colin, R. & Sourjik, V. 2016 Chemotaxis towards autoinducer 2 mediates autoaggregation in Escherichia coli. Nat. Commun. 7, 12984.CrossRefGoogle ScholarPubMed
Lauga, E., DiLuzio, W.R., Whitesides, G.M. & Stone, H.A. 2006 Swimming in circles: motion of bacteria near solid boundaries. Biophys. J. 90, 400–412.CrossRefGoogle ScholarPubMed
Leshansky, A.M. 2009 Enhanced low-Reynolds-number propulsion in heterogeneous viscous environments. Phys. Rev. E 80, 051911.CrossRefGoogle ScholarPubMed
Li, H., Shi, X-Q., Huang, M., , Chen, X., Xiao, M., Liu, C., Chate, H. & Zhang, H.P. 2019 Data-driven quantitative modeling of bacterial active nematics. Proc. Natl Acad. Sci. USA 116, 777–785.CrossRefGoogle ScholarPubMed
Liu, B., Breuer, K.S. & Powers, T.R. 2014 Propulsion by a helical flagellum in a capillary tube. Phys. Fluids 26, 011701.CrossRefGoogle Scholar
López, H.M., Gachelin, J., Douarche, C., Auradou, H. & Clément, E. 2015 Turning bacteria suspensions into superfluids. Phys. Rev. Lett. 115, 028301.CrossRefGoogle ScholarPubMed
Mathijssen, A.J.T.M., Doostmohammadi, A., Yeomans, J.M. & Shendruk, T.N. 2016 Hydrodynamics of micro-swimmers in films. J. Fluid Mech. 806, 35–70.CrossRefGoogle Scholar
Peng, Y., Liu, Z. & Cheng, X. 2021 Imaging the emergence of bacterial turbulence: phase diagram and transition kinetics. Sci. Adv. 7, eabd1240.CrossRefGoogle ScholarPubMed
Rabani, A., Ariel, G. & Be'er, A. 2013 Collective motion of spherical bacteria. PLoS One 8, e83760.CrossRefGoogle ScholarPubMed
Reinken, H., Nishiguchi, D., Heidenreich, S., Sokolov, A., Bär, M., Klapp, S.H.L. & Aranson, I.S. 2020 Organizing bacterial vortex lattices by periodic obstacle arrays. Commun. Phys. 3, 76.CrossRefGoogle Scholar
Rodenborn, B., Chen, C.-H., Swinney, H.L., Liu, B. & Zhang, H.P. 2013 Propulsion of microorganisms by a helical flagellum. Proc. Natl Acad. Sci. USA 110, E338–E347.CrossRefGoogle ScholarPubMed
Ryu, W.S., Berry, R.M. & Berg, H.C. 2000 Torque-generating units of the flagellar motor of Escherichia coli have a high duty ratio. Nature 403, 444–447.CrossRefGoogle ScholarPubMed
Saintillan, D. & Shelley, M.J. 2008 Instabilities and pattern formation in active particle suspensions: kinetic theory and continuum simulations. Phys. Rev. Lett. 100, 178103.CrossRefGoogle ScholarPubMed
Schwarz-Linek, J., Valeriani, C., Cacciuto, A., Cates, M.E., Marenduzzo, D., Morozov, A.N. & Poon, W.C.K. 2012 Phase separation and rotor self-assembly in active particle suspensions. Proc. Natl Acad. Sci. USA 109, 4052-4057.CrossRefGoogle ScholarPubMed
Simha, R.A. & Ramaswamy, S. 2002 Hydrodynamic fluctuations and instabilities in ordered suspensions of self-propelled particles. Phys. Rev. Lett. 89, 058101.CrossRefGoogle Scholar
Sokolov, A., Aranson, I.S., Kessler, J.O. & Goldstein, R.E. 2007 Concentration dependence of the collective dynamics of swimming bacteria. Phys. Rev. Lett. 98, 158102.CrossRefGoogle ScholarPubMed
Turner, L., Ryu, W.S. & Berg, H.C. 2000 Real-time imaging of fluorescent flagellar filaments. J. Bacteriol. 182, 2793–2801.CrossRefGoogle ScholarPubMed
Vizsnyiczai, G., Frangipane, G., Bianchi, S., Saglimbeni, F., Dell'Arciprete, D. & Di Leonardo, R. 2020 A transition to stable one-dimensional swimming enhances E. coli motility through narrow channels. Nat. Commun. 11, 2340.CrossRefGoogle ScholarPubMed
Xu, H., Dauparas, J., Das, D., Lauga, E. & Wu, Y. 2019 Self-organization of swimmers drives long-range fluid transport in bacterial colonies. Nat. Commun. 10, 1792.CrossRefGoogle ScholarPubMed
Yin, Y., Yu, H.-T., Tan, H., Cai, H., Chen, H.-Y., Lo, C.-J. & Guo, S. 2022 Escaping speed of bacteria from confinement. Biophys. J. 121, 4656–4665.CrossRefGoogle ScholarPubMed
Yoshinaga, N. & Liverpool, T.B. 2018 From hydrodynamic lubrication to many-body interactions in dense suspensions of active swimmers. Eur. Phys. J. E 41, 76.CrossRefGoogle ScholarPubMed
Zhang, B., Leishangthem, P., Ding, Y., Xu, X. 2021 An effective and efficient model of the near-field hydrodynamic interactions for active suspensions of bacteria. Proc. Natl Acad. Sci. USA 118, e2100145118.Google ScholarPubMed
Zhu, L., Lauga, E. & Brandt, L. 2013 Low-Reynolds-number swimming in a capillary tube. J. Fluid Mech. 726, 285–311.CrossRefGoogle Scholar

Huang et al. supplementary movie 1
A rotor with radius R=17 μm surrounded by free bacteria (FB) at area fraction ϕ=0.03 and activity aF=20 μm/s.

File
21 MB
Huang et al. supplementary movie 2
A rotor with radius R=15 μm surrounded by FB at ϕ=0.4 and aF=20 μm/s.
File
40.9 MB
Huang et al. supplementary movie 3
A rotor with radius R=15 μm surrounded by FB at ϕ=0.32 and aF=10 μm/s.
File
20 MB
Huang et al. supplementary material 4
Huang et al. supplementary material
File
4 MB