Introduction
Updated knowledge of Early Jurassic marine gastropods from the Andean region of Argentina has been provided recently by Ferrari (Reference Ferrari2009, Reference Ferrari2011, Reference Ferrari2012, Reference Ferrari2013, Reference Ferrari2014, Reference Ferrari2015) for the Chubut Basin, and by Damborenea and Ferrari (Reference Damborenea and Ferrari2008) and Ferrari et al. (2014, Reference Ferrari, Damborenea, Manceñido and Griffin2015) for the Neuquén Basin. Those contributions supplied an updated systematic database of the gastropod taxonomic composition in the region. These studies indicate that gastropods are well represented in the Early Jurassic of Argentina by three major taxa: Vetigastropoda, Caenogastropoda and Architectibranchia (=Opisthobranchia), although Patellogastropoda Lindberg, Reference Lindberg1986 and other representatives of Heterobranchia Haszprunar, Reference Haszprunar1988 are also present but less commonly found and less diversified in the marine deposits. The systematic and palaeobiogeographical knowledge about Argentinean vetigastropods was discussed recently by Ferrari (Reference Ferrari2015). The present paper is focused on the Caenogastropoda and Architectibranchia species and attempts to improve and update the systematic knowledge of these particular taxa in the Neuquén Basin, and to reassess the Jurassic gastropod collections currently existing in Argentina, supplying new evidence on Early Jurassic marine gastropods in South America. Nineteen caenogastropod and architectibranch species are reported here from the Neuquén Basin, among which seven are new to science; these are Procerithium subimbricatum n. sp., Microschiza weaveri n. sp., Naricopsina aequabilis n. sp., Naricopsina ballentae n. sp., Pictavia rothi n. sp., Oonia acuta n. sp., and Cylindrobullina brevispira n. sp. Seven other caenogastropods are testified for the first time in the area including Cryptaulax redelii Ferrari, Procerithium nulloi (Ferrari), Rhabdocolpus patagoniensis (Ferrari), Rhabdocolpus? sp., Pseudomelania feruglioi Ferrari, Pseudomelania aff. baugieriana (d’Orbigny), and Anulifera chubutensis Ferrari, extending their known palaeobiogeographical distribution in the Andean region of Argentina. The occurrences of Globularia catanlilensis (Weaver), Microschiza? americana (Möricke), Procerithium bodenbenderi (Behrendsen), and the architectibranchs Striactaeonina transatlantica (Behrendsen) and Cylindrobullina ovata (Behrendsen) is confirmed based on new material from the type and other localities in the Early Jurassic marine beds at the Neuquén Basin.
Geologic setting
A geological framework of the Argentinean Neuquén and Chubut basins was summarized by Ferrari (Reference Ferrari2015, p. 920, see references therein). The gastropod material described below comes from several localities in the Neuquén Basin, at the Neuquén and Mendoza provinces (Fig. 1.2).

Figure 1 (1) Map of the western part of South America showing the study area at the Neuquén Basin during the Hettangian-Toarcian. (2) Location map of the main localities with gastropod species described in the text. AML-N=Ayo. Malo; APR-N=Ayo. El Pedrero; ACHL-N=Ayo. Las Chilcas; PAY-N=Puesto Araya; PA-N=Paso del Portezuelo Ancho; APA-N=Arroyo del Portezuelo Ancho; AS-N=Ayo. Serrucho; CPH-N=Co. Puchenque; CÑC-N=Cañada Colorada; AÑR-N=Ayo. Ñiraico; ER-N=Estación Rajapalo; CLI-N=Catán Lil; ESI-N=Ea. Santa Isabel; CC-N=Carrán Curá; CR-N=Co. Roth; S-N=Sañicó (modified from Ferrari, Reference Ferrari2015).
In Mendoza Province, Hettangian to Toarcian sediments of the Atuel–western Malargüe depocentre contain gastropods at Arroyo Malo (Fig. 1.2, locality AML-N), Arroyo El Pedrero (Fig. 1.2, locality APR-N), Arroyo Las Chilcas (Fig. 1.2, locality ACHL-N), Puesto Araya (Fig. 1.2, locality PAY-N), Paso del Portezuelo Ancho (Fig. 1.2, locality PA-N), Arroyo del Portezuelo Ancho (Fig. 1.2, locality APA-N), Arroyo Serrucho (Fig. 1.2, locality AS-N), Cerro Puchenque (Fig. 1.2, locality CPH-N), and Cañada Colorada (Fig. 1.2, locality CÑC-N). Gastropods were also found in Pliensbachian−Toarcian deposits in the Cuyo Group beds within the Neuquén embayments in northern and southern Neuquén Province, at Arroyo Ñiraicó (Fig. 1.2, locality AÑR-N), Estación Rajapalo (Fig. 1.2, locality ER-N), Catán Lil (Fig. 1.2, locality CLN-N), Estancia Santa Isabel (Fig. 1.2, locality ESI-N), Carrán Curá (Fig. 1.2, locality CC-N), Cerro Roth (Fig. 1.2, locality CR-N), and Sañicó (Fig. 1.2, locality S-N).
Materials and methods
The gastropod material here described was collected by S. Damborenea, A. Riccardi, M. Manceñido, J. Echevarría (Museo de Ciencias Naturales de La Plata), S. Lanés (Universidad de Buenos Aires), C. Gulisano (YPF), and the present author during several fieldtrips to the Neuquén Basin. Stratigraphical sections for most of the localities yielding gastropods in the Neuquén Basin were described by Damborenea et al. (1975), Damborenea (Reference Damborenea1987), and Damborenea and Manceñido (2005). Specimens described here were found in the Early Jurassic at different stratigraphic levels, spanning the Hettangian to early Toarcian, according to the current ammonite biozonation (Riccardi Reference Riccardi2008a, Reference Riccardib; Riccardi et al., Reference Riccardi, Damborenea, Manceñido and Leanza2011).
Most of the Argentinean gastropod material described here is housed in the Museo de Ciencias Naturales de La Plata (MLP) collection and in the Museo Paleontológico Carmen Funes, Plaza Huincul (MCF-PIPH). Material described by Weaver (Reference Weaver1931), which is currently housed in the Burke Museum of Natural History and Culture (Seattle, USA) (BM), was revised and casts are kept in the La Plata Museum collections. Additional gastropod material described by Gründel (Reference Gründel2001a) from the Early Jurassic of Chile and Argentina which is currently stored at the Museum für Naturkunde, Berlin (MB.Ga.) was also revised, subsequently compared with the specimens here analyzed and included in this paper. The Argentinean specimens were prepared by technical staff (Leandro Canessa and Roberto Pfeiffer) at the MEF laboratory, and by Santiago Bessone at the CENPAT laboratory. Latex cast were obtained from specimens preserved as external moulds and the material was coated with ammonium chloride to enhance sculpture details for photography. Photographs were taken using a digital camera, and using scanning electronic microscopy (SEM) at ALUAR (Pto. Madryn).
Repositories and institutional abbreviations
MLP=Museo de Ciencias Naturales de La Plata, Buenos Aires, Argentina; MCF-PIPH=Museo Paleontológico Carmen Funes, Plaza Huincul, Neuquén, Argentina; MB.Ga.=Museum für Naturkunde Humboldt-Universität zu, Berlin, Germany; MPEF-PI=Museo Paleontológio Egidio Feruglio; BM=Burke Museum of Natural History and Culture, Seattle, USA; CENPAT=Centro Nacional Patagónico, Puerto Madryn, Chubut, Argentina; ALUAR=Aluminio Argentino, Puerto Madryn, Chubut, Argentina; YPF=Yacimientos Petrolíferos Fiscales.
Systematic paleontology
Superorder Caenogastropoda Cox, 1960
Order Sorbeoconcha Ponder and Lindberg, Reference Ponder and Lindberg1997
Suborder Cerithiomorpha Golikov and Starobogatov, Reference Golikov and Starobogatov1975
Superfamily Cerithioidea Fleming, 1822
Family Procerithiidae Cossmann, Reference Cossmann1906
Subfamily Procerithiinae Cossmann, Reference Cossmann1906
Genus Cryptaulax Tate, Reference Tate1869
Type species
Procerithium (Xystrella) protortile Cox, 1965, pro Cerithium tortile Hébert and Eudes-Deslongchamps, Reference Hébert and Eudes−Deslongchamps1860, from the Middle Jurassic (Callovian) of France.
Occurrence
Cosmopolitan; Upper Triassic–Lower Cretaceous.
Cryptaulax redelii Ferrari, Reference Ferrari2012

Figure 2 (1−7) Cryptaulax redelii Ferrari, Reference Ferrari2012. (1) MLP 35459, lateral view. (2−7) MLP 12193/a. (2, 3) Lateral and apertural views, (4, 5) lateral views, (6, 7) ornament detail. (8−10) Procerithium nulloi (Ferrari, Reference Ferrari2009). (8) MLP 35464, lateral view. (9) MLP 12163, lateral and apertural view. (10) MCF-PIPH 567, lateral view. (11) Procerithium bodenbenderi (Behrendsen, Reference Behrendsen1891), MLP 35462, lateral view. (12, 13) Procerithium subimbricatum n. sp., MLP 35460, holotype, lateral views. (14−17) Procerithium (Rhabdocolpus) patagoniensis Ferrari, Reference Ferrari2012, MLP 35465. (14−17) Lateral views; (18) Rhabdocolpus? sp., MLP 35517, lateral view. (19−27) Microschiza weaveri n. sp. (19, 20) MLP 25020 (cast of BMNH 44091/SA1029), paratype, lateral views. (21−24) MLP 35468, holotype. (21) lateral view; (22, 23) lateral and apertural views; (24) apertural detail. (25−27) MLP 35467, paratype. (25) Lateral view; (26) apical view; (27) lateral and apertural. (28) Microschiza? americana, MLP 25021 (cast of BMNH 269/SA1029) lateral view. (29−35) Pseudomelania feruglioi Ferrari, Reference Ferrari2013. (29) MLP 35472, lateral and apertural views. (30, 31) MLP 35473, lateral and apertural views. (32) MLP 35469, lateral and apertural view. (33) MLP 19638, lateral view. (34−37) MLP 35459. (34) Three specimens in lateral views; (35) juvenile and mature teleoconch in latera view; (36) juvenile teleoconch detail; (37) basal detail. (38, 39) Pseudomelania aff. baugieriana (d’Orbigny, 1850) MLP 12174, fragmentary specimen in lateral views. (40) Anulifera chubutensis Ferrari, Reference Ferrari2013, MLP 35516, lateral view.
2012 Cryptaulax redelii Ferrari; Reference FerrariFerrari, p. 327, fig. 2I−K.
Holotype
MPEF-PI 3508, from Lomas Occidentales, Chubut Province, Argentina in the Mulanguiñeu Formation (Ferrari, Reference Ferrari2012, fig. 2I−K).
Occurrence
Cerro Roth and Estación Rajapalo, Pliensbachian, Neuquén Province. Previously known from Lomas Occidentales, Pliensbachian-Toarcian, Chubut Province, Argentina (Ferrari, Reference Ferrari2012).
Description
See original description of the species in Ferrari (Reference Ferrari2012, p. 327).
Materials
MLP 12193a/b, 35459; three specimens preserved as external moulds.
Dimensions (mm)
MLP 12193/a: height=17.0; width=8.7; apical angle=47°*. MLP 12193/b: height=16.2; width=9.4; apical angle=42°*. MLP 35459: height=8.7; width=3.9; apical angle=51°. *=fragmentary data.
Remarks
According to the diagnosis proposed by Ferrari (Reference Ferrari2012, p. 327), the specimens here analyzed certainly represent members of Cryptaulax redelii. Ferrari (Reference Ferrari2012, p. 327) reported this species from the Early Jurassic (Pliensbachian-Toarcian) of the Chubut Province. Cryptaualx weaveri Ferrari and Damborenea (2015, p. 634, fig. 2. 16−19), from the early Bajocian of the Neuquén Basin, differs from C. redelii in having a high number of primary and secondary spiral cords on mature teleoconch whorls, and more conspicuous nodes at the crossing points of spiral and axial elements.
For comparison with other similar Cryptaulax species from Argentina and western Tethys, see Ferrari (Reference Ferrari2012).
Genus Procerithium Cossmann, Reference Cossmann1902
Type species
Procerithium quinquegranosum Cossmann, Reference Cossmann1902, from the Early Jurassic (Hettangian) of France.
Occurrence
Europe, Africa, New Zealand, Asia, Antarctica (?), and South America; Lower Jurassic (Hettangian)–Lower Cretaceous (Barremian).
Remarks
Representatives of Procerithium were characterized by Gründel (Reference Gründel1999). According to the author, this genus groups “slender shells, with numerous whorls, outline of the whorls straight to convex, with distinct sutures, on later whorls the number of spiral and axial elements is variable with nodes at the crossing points, base weakly convex, angulated to the upper part of the whorls, with one strong spiral at the periphery and 1-2 weaker spirals toward the centre.” In the present paper, the diagnosis of Gründel (Reference Gründel1999) is followed.
Procerithium nulloi (Ferrari, Reference Ferrari2009)
2009 Cryptaulax nulloi Reference FerrariFerrari, p. 457, fig. 3F−H.
2012 Procerithium nulloi (Ferrari); Reference FerrariFerrari, p. 328, figs. 2L−T, 4A−F.
Holotype
MPEF-PI 1870, from Puesto Currumil, Chubut Province, Argentina in the Osta Arena Formation (Ferrari, Reference Ferrari2012, figs. 2P−Q).
Occurrence
Cerro Roth and Subida a Sañicó, Pliensbachian, Neuquén Province, Argentina. Previously known from Puesto Currumil and Lomas Occidentales, Pliensbachian−Toarcian, Chubut Province, Argentina (Ferrari, Reference Ferrari2009, Reference Ferrari2012).
Description
See original description of the species in Ferrari (Reference Ferrari2009, p. 457; Reference Ferrari2012, p. 328).
Materials
MLP 12163, 35464, MCF-PIPH 567; five fragmentary and recrystallized specimens; one external mould.
Dimensions (mm)
MCF-PIPH 567: height=22.7; width=6.4; apical angle=26º. MLP 35464-1: height=11.0; width=3.3; apical angle=28º. MLP 35464-2: height=11.5; width=4.0. MLP 35464-3: apical angle=36º.
Remarks
The specimens reported here fits with the characterization of the species proposed by Ferrari (Reference Ferrari2009, p. 457, fig. 3F−H, as Cryptaulax nulloi; Ferrari, Reference Ferrari2012, p. 328, figs. 2L−T, 4A−F), from the Early Jurassic of the Chubut Province, Argentina.
For comparisons of Procerithium nulloi with other similar species of South America and western Tethys, see Ferrari (Reference Ferrari2009, Reference Ferrari2012).
Procerithium bodenbenderi (Behrendsen, Reference Behrendsen1891) new combination
1891 Cerithium bodenbenderi Reference BehrendsenBehrendsen, p. 382, pl. 22, fig. 4.
1922 Cerithium bodenbenderi Behrendsen; Reference BehrendsenBehrendsen, p. 168, pl. 1, fig. 4.
Holotype
A well-preserved specimen, originally described by Behrendsen (Reference Behrendsen1891) from the Early Jurassic of Paso del Portezuelo Ancho, Mendoza Province, Argentina (Behrendsen, Reference Behrendsen1891, p. 382, pl. 22, fig. 4). Behrendsen’s original material, which should be housed at Göttingen, Germany, could not be examined.
Occurrence
Cerro Puchenque and Paso del Portezuelo Ancho, Pliensbachian, Mendoza Province, Argentina.
Description
Dextral, turriculate, high-spired, and medium-sized shell. The teleoconch comprises up to 10 (or more) flattened whorls, with a height of 34.0 mm and a width of 9.0 mm. The apex forms an angle of 30º, although earliest teleoconch whorls are missing in the available specimens. Suture is deeply impressed in a spiral furrow. The ornament is spiral predominantly, consisting of one to two primary and strongly nodular cords located in adapical and abapical position on the outer face; a second order of weaker spiral cords appear between the primary ones, and in the middle of mature whorls are present in numbers of 12−14. The presence of two strong nodular spiral keels bordering the sutures gives the outer face of the shell a slightly concave appearance. The base is flat, ornamented by weak spiral cords; the aperture is holostomatous and subcircular.
Materials
MLP 35462, 35463; two fragmentary and recrystallized specimens.
Dimensions (mm)
MLP 35462: height=34.0; width=9.0; apical angle=40º. MLP 35463: height=43.0; width=10.5.
Remarks
Procerithium bodenbenderi was originally described by Behrendsen (Reference Behrendsen1891) under the genus Cerithium. This species is here reassigned to the genus Procerithium, following the updated characterization of Gründel (Reference Gründel1999) (see also Ferrari, Reference Ferrari2012).
Behrendsen’s species is very similar in ornament to Procerithium nulloi (Ferrari). The differences between both species were clearly stated out by Ferrari (Reference Ferrari2012): “Procerithium nulloi is very similar to Cerithium bodenbenderi Behrendsen (Reference Behrendsen1891, p. 382, pl. 22, fig. 4) from the Early Jurassic of Mendoza Province (Argentina). They share the same general shell morphology, with a high-spired shell, slightly concave whorls and nodose rows bordering the sutures on each mature whorl. However, C. bodenbenderi is much larger, has more conspicuous nodes and a greater number (12–14) of spiral ribs on mature whorls.”
P. bodenbenderi has a paleogeographical distribution limited to the Mendoza Province, whereas the northernmost occurrence of P. nulloi in the Andean region of Argentina reaches the southern Neuquén Province.
Procerithium subimbricatum new species
Holotype
MLP 35460; one specimen preserved as external mould.
Diagnosis
Sub-imbricate, turriculate; subhorizontal sutural ramp; spiral ornament predominant on the outer face; adult teleoconch with 2 primary spiral cords; 2 secondary spirals cords between the primary ones on mature whorls; nodes at the crossing points of spiral and axial elements.
Occurrence
Paso del Portezuelo Ancho, late Pliensbachian, Mendoza Province, Argentina.
Description
Dextral, turriculate, medium-sized and high-spired shell. The apex is broken and earliest teleoconch whorls are missing. The protoconch is not preserved. The fragmentary teleoconch has a height of 18.2 mm and a width of 7.7 mm, and consists of seven slightly imbricated whorls. The ramp is flat to inclined approximately 45º from sutures; toward the outer face the whorl become flat and vertical. Suture is clearly impressed in a spiral furrow. The shell is ornamented by spiral, axial and nodular elements. The ramp bears weak and regularly spaced axial ribs. The outer face of whorls is ornamented by four regularly spaced spiral cords; two primary cords are located in an adapical and abapical position on the outer face; two other secondary cords are median and located between the primary ones. The primary spiral cords become stronger toward mature growth stages. Strong and regularly spaced axial ribs intercept the spiral elements on the outer face forming nodes at the crossing points. Nodes are more conspicuous and acute at the intersection with the primary spiral cords. On last whorl, nodes are even stronger and a third spiral and nodular cord appears below the abapical one, delimiting the base. The base is flat to slightly convex, ornamented by 1 spiral cord. The apertural characters are missing.
Etymology
Referred to the strongly sub-imbricated teleoconch whorls.
Remarks
In the present paper, the concepts of Gründel (Reference Gründel1999, Reference Gründel2003) and Schulbert and Nützel (2013) are followed to include the specimen here described into the genus Procerithium. The variable number of spiral and axial elements on mature whorls, together with the presence of nodes at the crossing points, are considered as the main diagnostic characters; however, none of the protoconch features are preserved in the available material.
Procerithium subimbricatum n. sp. differs from Procerithium nulloi (Ferrari, Reference Ferrari2009; see above) in being larger, having an imbricate outline shell, a sub-horizontal sutural ramp, and axial ribs on the shell surface. P. subimbricatum resembles Procerithium (Rhabdocolpus) patagoniensis Ferrari; however, the new species has a sub-imbricate shell outline, a more reticulate ornament pattern, secondary spiral cords on the middle of whorls, and more developed nodes at the crossing points of axial and spiral elements.
Genus Rhabdocolpus Cossmann, Reference Cossmann1906
Type species
Melania scalariformis Deshayes, 1830 from the Middle Jurassic of France.
Occurrence
Europe, New Zealand, Antarctica and South America; Late Triassic?–Late Jurassic.
Remarks
The characterization of Rhabdocolpus was stated out by Gründel (Reference Gründel1999) and Bandel et al. (Reference Bandel, Gründel and Maxwell2000). Both authors included within Rhabdocolpus high-spired and flattened to slightly convex outline shell, with a horizontal to strong sloping sub-sutural ramp, opisthocyrt axial ribs on the teleoconch, forming nodes at the crossing point with spiral elements, and base with an adapical channel and lacking abapical notch. The genus is common in the Jurassic of Europe, and Haas (Reference Haas1953) reported the first occurrence of Rhabdocolpus in the Upper Triassic of the Pucará group (South America). In the present paper, the diagnosis of Gründel (Reference Gründel1999) and Bandel et al. (Reference Bandel, Gründel and Maxwell2000) are followed.
Rhabdocolpus patagoniensis (Ferrari, Reference Ferrari2012) new combination
2009 Cryptaulax cf. damboreneae Reference FerrariFerrari, p. 454, fig. 3E.
2012 Procerithium (Rhabdocolpus) patagoniensis Reference FerrariFerrari, p. 329, figs. 4G−P, 5A−K.
2015 Procerithium (Rhabdocolpus) patagoniensis Ferrari; Reference FerrariFerrari, p. 84, fig. 5O−R.
2015 Procerithium (Rhabdocolpus) patagoniensis Ferrari; Reference Ferrari and BessoneFerrari and Bessone, p. 353, fig. 3L−M.
Holotype
MPEF-PI 3517, from Lomas Occidentales, Chubut Province, Argentina in the Mulanguiñeu Formation (Ferrari, Reference Ferrari2012, figs. 4G−J).
Occurrence
Arroyo Las Chilcas, Río Atuel region, late Sinemurian (Orthechioceras-Paltechioceras Zone), Mendoza Province, Argentina. Previously known from Lomas Occidentales and La Casilda, Pliensbachian−Toarcian, Chubut Province, Argentina (Ferrari, Reference Ferrari2012).
Description
Dextral, turriculate, small-sized and high-spired shell. The protoconch is not preserved. The teleoconch has a mean height of 4.7 mm and a mean width of 2.6 mm, and consists of six flattened to slightly convex whorls. The mean apical angle is of 59º, although earliest teleoconch whorls are missing. A narrow sub-sutural ramp gives the periphery of the shell a step-like outline. Suture is weakly impressed. The axial ornament is predominant on all teleoconch whorls and consists of opisthocyrt to straight ribs, which run from suture to suture. The axial ribs are strong and slightly orthocline on juvenile whorls, becoming more spaced and opisthocyrt toward mature growth stages. The axial elements intercept five to six regularly spaced spiral cords on teleoconch whorls, forming nodes at the crossing points. The nodes on the adapical spiral cords are spiny and stronger than the others. The base is convex and ornamented by four visible regularly spaced spiral cords. The aperture is fragmentary and subcircular.
Materials
MLP 35465, 35466; three specimens preserved as negative moulds.
Dimensions (mm)
MLP 35465-1: height=4.6; width=2.5; apical angle=57°. MLP 35465-2: height=4.9; width=3.0; apical angle=60°. MLP 35466: height=4.7; width=2.5; apical angle=60°.
Remarks
Ferrari (Reference Ferrari2012, p. 329) and Ferrari and Bessone (Reference Ferrari and Bessone2015, p. 353) described this species as Procerithium (Rhabdocolpus) patagoniensis from the Early Jurassic (Pliensbachian−Toarcian) of the Chubut Province. Here it is transferred to the genus Rhabdocolpus following the diagnosis of Gründel (Reference Gründel1999) and Bandel et al. (Reference Bandel, Gründel and Maxwell2000). Representatives of Rhabdocolpus have been reported recently from the early Bajocian of the Neuquén Basin with the species Rhabdocolpus (Infacerithium) excavatus Ferrari and Damborenea (2015, p. 635, fig. 2.20−22). The Bajocian form, however, has a less step-like outline shell with more convex teleoconch whorls, a less horizontal sub-sutural ramp, and more rounded and conspicuous nodes at the crossing point of spiral and axial ribs. Cryptaulax cf. damboreneae Ferrari (Reference Ferrari2009, p. 545, fig. 3E), from the early Toarcian of Cerro Puchenque locality, was originally described by the author as a conespecific species of Cryptaulax damboreneae; however, it has a larger shell, orthocline to slightly opistocline axial ribs, and stronger spiral cords. Cryptaulax cf. damboreneae is here included as a synonymous of Rhabdocolpus patagoniensis (Ferrari, Reference Ferrari2012) (see above).
For comparisons with other similar procerithiids species, see Ferrari (Reference Ferrari2012).
Rhabdocolpus? sp.
Occurrence
Cerro Puchenque, Pliensbachian, Mendoza Province, Argentina.
Description
Dextral, turriculate, high-spired, and medium-sized shell. The fragmentary teleoconch has a height of 23.2 mm and a width of 7.5 mm, and consists of ten step-like whorls. The sub-sutural ramp is narrow and horizontal, and sutures are weakly incised in a spiral furrow on the subsutural area. The upper portion of whorls is delimited by a convex, nodular swollen belt, which gives the periphery of the shell a step-like appearance. Nodes are conspicuous, rounded, and more or less spaced. The nodular swollen belt also bears fine, separated, and almost imperceptible spiral cords. Whorls become flat to slightly concave toward the outer face. Basal and apertural characters are missing.
Materials
MLP 35517; one fragmentary specimen.
Remarks
The single specimen described is assigned to Rhabdocolpus based on the presence of a high-spired shell and an horizontal sub-sutural ramp; however, the typical axial ribs on the whorls flank intercepted by spiral cords and forming nodes at the crossing points, and the adapical channel on base are not visible in the available material; thus, the specimen is left in open nomenclature. Rhabdocolpus? sp. differs from Rhabdocolpus patagoniensis in being larger and in lacking the strong opisthocyrt axial ribs intercepted by spiral cords on the shell surface.
Suborder Hypsogastropoda Ponder and Lindberg, Reference Ponder and Lindberg1997
Superfamily Littorinoidea Gray, Reference Gray1840
Family Purpurinidae Zittel, Reference von Zittel1895
Genus Microschiza Gemmellaro, Reference Gemmellaro1878
Type species
Turbo philenor d’Orbigny, 1847, from the Early Jurassic (Sinemurian) of France.
Occurrence
Early Jurassic−Early Cretaceous; Europe, Japan, South America.
Remarks
Microschiza was originally proposed by Gemmellaro (Reference Gemmellaro1878) as a subgenus of Chemitzia, to distinguish sub-imbricated and bucciniform shells, with a low spire, aperture oval, strongly expanded abapically and acute posteriorly, and ornamented by sinusoidal axial ribs. Later authors (Cossmann, Reference Cossmann1909; Delpey, Reference Delpey1937; Wenz, 1938; Kase, Reference Kase1984) included Microschiza within Pseudomelaniidae, even though Fischer and Weber (Reference Fischer and Weber1997) assigned the genus to the family Purpurinidae, on the basis of its similarities with the genus Purpurina d’Orbigny, 1850. However, the systematic position of Microschiza still remains unclear. In the present paper, the proposal of Fischer and Weber (Reference Fischer and Weber1997) is followed.
The genus Microschiza is not a very common genus within the Mesozoic gastropod faunas, and has a relatively narrow distribution range, being only mentioned in Early Jurassic and Early Cretaceous deposits of Europe and Japan, respectively. The new species described below represent the first occurrence of Microschiza in the Jurassic of Argentina.
Microschiza weaveri new species
1931 Amberleya cf. americana Möricke; Reference WeaverWeaver: p. 370. (pars)
2001a Microschiza sp. Reference GründelGründel, p. 60, pl. 4, figs. 15−16.
Holotype
MLP 35468; one recrystallized specimen.
Paratypes
MLP 35467; one recrystallized specimen. BMNH 44091/SA1029 of Weaver´s collection and originally assigned by Weaver (Reference Weaver1931) to Amberleya cf. Americana (cast MLP 25020).
Diagnosis
Globular and slightly bucciniform shell; convex whorls; sutural ramp narrow and horizontal; prosocyrt to sinusoidal axial ribs; aperture holostomatous and oval; outer lip strongly convex; columellar lip thickened.
Occurrence
Catán Lil, Plienbachian, Neuquén Province, Argentina (Weaver collection); Arroyo Las Chilcas, Río Atuel region, late Sinemurian (Orthechioceras−Paltechioceras Faunule), Mendoza Province; Cerro de Cuevitas, Early Jurassic (Sinemurian), Chile (Gründel, Reference Gründel2001a).
Description
Dextral, globular to slightly bucciniform, gradate, medium- to large-sized, and low-spired shell. The protoconch is not preserved. The teleoconch has a mean height of 41.6 mm and a mean width of 33.6 mm, and consists of three to four strongly convex whorls, but slightly concave in the middle, and last whorl greatly expanded. The apex forms an angle of 122º. The sub-sutural ramp is narrowly horizontal. Suture is incised in a concave furrow. The axial ornament is predominant and visible on last whorl, and consists of prosocyrt to sinusoidal and strong axial ribs. The base is strongly convex to angular and smooth. The aperture is holostomatous and forms an adapical notch. The outer lip is convex and the columellar lip thickened as a callus. The basal lip is expanded abapically.
Etymology
Dedicated to C. Weaver, who contributed to the paleontological knowledge of Jurassic marine gastropods from the Andean region of Argentina and who collected one of the paratypes.
Materials
MB.Ga.785; one recrystallized teleoconch. Specimen originally assigned by Gründel (Reference Gründel2001a) to Microschiza sp., from the Early Jurassic (Sinemurian) of Chile.
Dimensions (mm)
MLP 35468, holotype: height=41.4; width=32.2; apical angle=122°. MLP 35467, paratype: height=24.0; width=20.2; apical angle=115°. MLP 25021, paratype: height=64.5; width=55.0; apical angle=131°. MB.Ga.785: height=36.6; width=27.0; apical angle=122º.
Remarks
The material here described shows the diagnostic characters of Microschiza, such as bucciniform and sub-imbricate shell, with the ornament consisting on sinusoidal axial ribs or furrows, oval aperture.
The type species Microschiza philenor (d’Orbigny) (Fisher and Weber, Reference Fischer and Weber1997, p. 128, pl. 23, figs. 22−23), from the Early Jurassic (Sinemurian) of France, is smaller than the Argentinean form, has a rather higher-spired shell with a more acute apex; a more gradate shell shape; a more developed ornament pattern with axial and spiral elements forming nodes at intersections points.
The Cretaceous species, Microschiza japonica (Nagao) (in Kase, Reference Kase1984; p. 110, pl. 4, figs. 5−7), from the Early Cretaceous of Japan, is similar to M. weaveri n. sp.; however, M. japonica is smaller, has the shell surface sculptured by axial furrows which are variable in strength and number, and its axial furrow usually has nodes on the depression of body whorls. The last character is missing in the Argentinean species. Microschiza aff. heybroeki Delpey, Reference Delpey1940 (Kase, Reference Kase1984; p. 112, pl. 14, fig. 9a−b), from the Early Cretaceous of Japan, differs from M. weaveri in having nodes at the shouldered edge of whorls and in lacking axial ribs.
Microschiza? americana (Möricke, Reference Möricke1894) new combination
1894 Amberleya americana Reference MörickeMöricke: p. 29, pl. 4, fig. 8a−b.
1931 Amberleya cf. americana Möricke; Weaver: p. 370. (pars)
Type material
Specimens described by Möricke (Reference Möricke1894, p. 29, pl. 4, fig. 8a−b) from the Lower Jurassic beds at Mina Amolanas, Chile. The material was originally housed at Strassburg University but could not be examined.
Occurrence
Catán Lil, Pliensbachian, Neuquén Province, Argentina.
Description
Dextral, gradate, medium-sized, and moderately high-spired shell. The teleoconch has a height of 51.3 mm and a width of 39.4 mm. The apex forms an angle of 64º. The horizontal sub-sutural ramp is narrowly developed. The outer face is flat to slightly concave in the middle. On the periphery of whorls, a row of rounded nodes is visible. Suture is weakly incised. The base is flat to angular and slightly concave. The aperture is missing. The columellar lip is thickened.
Materials
Cast MLP 25021 (original material BMNH 269/SA1029 of Weaver’s collection); one fragmentary specimen.
Remarks
The material here analyzed was assigned by Weaver (Reference Weaver1931) to Amberleya cf. americana, although it seems to be a member of Microschiza, because it has a bucciniform and sub-imbricated shell shape. However, the prosocyrt and sinusoidal axial ribs, together with the strongly oval aperture, characteristic of Microschiza, are not visible in the present material, and thus it is left in open nomenclature. Ferrari (Reference Ferrari2009) stated out that Amberleya itself is of rather disputable identity, and that descriptions and illustrations available in Möricke (Reference Möricke1894, p. 29, pl. 4, fig. 8a−b) show that Amberleya americana differs significantly from the type species Amberleya bathonica Cox and Arkell, 1950 (see Ferrari et al., 2014, fig. 2). Ferrari et al. (2014) discussed the systematic status of the genus Amberleya and restricted the usage of the name Amberleya to shells with small umbilicus, a single abapical keel and adapical spiral cord, ornamented by blunt nodes and smooth base; this restricts it largely to the type species A. bathonica.
Microschiza? americana (Möricke, Reference Möricke1894) differs from Microschiza weaveri in having a slightly more gradate and higher-spired shell, narrower apical angle, less convex whorls, more developed nodular elements, and lacking sinusoidal and prosocyrt axial ribs.
Order Ptenoglossa Gray, Reference Gray1853
Superfamily Pseudomelanoidea R. Hornes, Reference Hornes.1884
Family Pseudomelaniidae R. Hornes, Reference Hornes.1884
Genus Pseudomelania Pictet and Campiche, 1862
Type species
Pseudomelania gresslyi Pictet and Campiche, 1862, by subsequent designation by Wenz (1938); from the Lower Cretaceous (Neocomian) of Switzerland.
Occurrence
Triassic−Cretaceous (Kaim, Reference Kaim2004). Europe, Asia, Africa, Madagascar, New Zealand and America.
Pseudomelania feruglioi Ferrari, Reference Ferrari2013
2013 Pseudomelania feruglioi Reference FerrariFerrari, p. 582, figs. 2F−I, 3A−D.
?2015 Pseudomelania feruglioi Ferrari; Reference Ferrari and BessoneFerrari and Bessone, p. 354, fig. 3O.
Holotype
MPEF-PI 4155, from Lomas Occidentales, Chubut Province, Argentina in the Mulanguiñeu Formation (Ferrari, Reference Ferrari2013, fig. 2F).
Emended diagnosis
Dextral, anomphalous and turriculate shell; teleoconch up to ten whorls; juvenile teleoconch slightly convex or step-like with a convex swollen belt in the adapical sutural area; mature teleoconch with a strong adapical spiral keel on each whorl; opisthocline growth lines on the middle of mature whorls, and prosocyrt on the lower portion of whorls and on base; aperture oval forming a narrow siphonal canal.
Occurrence
Cerro Roth and Estación Rajapalo, Pliensbachian, Neuquén Province; Cerro Puchenque, Arroyo Serrucho and Arroyo del Portezuelo Ancho, Pliensbachian, Mendoza Province, Argentina. Previously known from Lomas Occidentales and Betancourt, Pliensbachian−Toarcian, Chubut Province, Argentina (Ferrari, Reference Ferrari2013).
Description
Dextral, phaneromphalous, turriculate, high-spired and medium- to large-sized shell. The teleoconch has a mean height of 39.9 mm and mean width of 12.4 mm. The mean apical angle is of 37°, although earliest teleoconch whorls are missing in the better preserved specimens. In the most complete specimens, the teleoconch is composed of eight whorls. The earliest whorls are slightly step-like with a convex swollen belt in the adapical sutural area; the whorls become flat to slightly concave toward mature growth stages. Suture is incised and clearly visible on juvenile whorls. Ornament is poorly developed on juvenile whorls, and stabilizes toward mature teleoconch with an adapical spiral keel on each whorl. Opisthocline collabral lines are visible on the middle of mature whorls and become prosocyrt on the lower portion of whorls and on base. The base is strongly convex and the aperture holostomatous, oval and acute adapically, with the columellar lip strongly expanded abapically forming a narrow siphonal canal.
Materials
MLP 19638, 25097, 35459, 35469, 35470, 35471, 35472, 35473, 35474, 35475, 35476, 35477, 35478; MCF-PIPH 693, 681, 688, 557, 541; 18 recrystallized specimens.
Dimensions (mm)
Table 1 Dimensions (mm) of Pseudomelania feruglioi Ferrari, Reference Ferrari2013. −=missing data; j=juvenile.

Remarks
Pseudomelania feruglioi Ferrari (Reference Ferrari2013) was previously reported by Ferrari (Reference Ferrari2013) from the Early Jurassic of the Chubut Province. The specimens from the Chubut Province, however, do not show the step-like earliest whorls with the convex swollen belt in the adapical sutural area, and have instead slightly convex juvenile whorls. Moreover, the Chubut specimens lack the clearly visible opisthocline growth lines in the middle of mature whorls and the prosocyrt lines on base, and lack the columellar lip forming a narrow siphonal canal. These differences are considered here as intraspecific variability within Pseudomelania feruglioi, and have been included in the emended the diagnosis of the species (see above).
The species resembles Pseudomelania procera Eudes-Deslongchamps, 1842 (in d’Orbigny, 1850, p. 41, pl. 239, figs. 2−3), from the Middle Jurassic (Bajocian) of England and France, although the European species is larger and has a narrower apical angle.
For comparisons with similar and coeval Pseudomelania species, see Ferrari (Reference Ferrari2013, p. 584).
Pseudomelania aff. baugieriana (d’Orbigny, 1850)
Occurrence
Carrán Curá, Early Jurassic (Pliensbachian), Neuquén Province, Argentina.
Description
Dextral and turriculate shell. The fragmentary teleoconch is an internal mould and consists of two whorls; the ramp of whorls is narrow. The outer face becomes flat to slightly convex. The periphery of whorls is bordered by a row of small and rounded nodes. The base is strongly convex and the aperture oval with the outer lip convex and the columellar lip concave.
Materials
MLP 12174; one recrystallized specimen.
Remarks
Pseudomelania aff. baugieriana is similar to Pseudomelania baugieriana (d’Orbigny, 1850) (p. 40. pl. 243, fig. 5; Fischer and Weber, Reference Fischer and Weber1997, p. 17, pl. 2, fig. 16), from the Early Jurassic (Toarcian) of France; however, the Argentinean material shows a row of weak nodes on the periphery of the shell; the former character is missing in P. baugieriana (d’Orbigny). Pseudomelania lorieri (d’Orbigny, 1850) (p. 38, pl. 238, fig. 1; Fischer and Weber, Reference Fischer and Weber1997, p. 16, pl. 1, fig. 28), from the Early Jurassic (Toarcian) of France, resembles also the species here described; but, the European form has more convex teleoconch whorls, with the last one less expanded, and lacks the peripheral row of nodes. Pseudomelania repeliniana (d’Orbigny, 1850) (p. 39, pl. 238, fig. 9; Fischer and Weber, Reference Fischer and Weber1997, p. 16. pl. 2, figs. 14−15), from the Toarcian of France, has less convex whorls and a more developed axial ornament than the Argentinean species. Pseudomelania (P.) disparilis (d’Orbigny, 1850) (p. 47, pl. 243, fig. 6; Fischer and Weber, Reference Fischer and Weber1997, p. 21, pl. 2, fig. 9), from the Middle Jurassic (Bajocian) of France, is very similar to P. aff. baugieriana, but the European species has less convex whorls and a more developed horizontal ramp.
Family Protorculidae Bandel, Reference Bandel1991
Genus Anulifera Zapfe, Reference Zapfe1962
Type species
Zygopleura (Anulifera) variabilis Zapfe, Reference Zapfe1962, from the Rhaetian (Late Triassic) of Austria.
Occurrence
Late Triassic–Middle Jurassic. Europe, Asia, South America.
Anulifera chubutensis Ferrari, Reference Ferrari2013
2013 Anulifera chubutensis Reference FerrariFerrari, p. 585, fig. 3F−G.
Holotype
SEGEMAR 25000 (cast MPEF-PI 4107), from Lomas Occidentales, Chubut Province, Argentina in the Mulanguiñeu Formation (Ferrari, Reference Ferrari2013, fig. 3F).
Occurrence
Estancia Santa Isabel, Pliensbachian, Neuquén Province. Previously known from Lomas Occidentales, Pliensbachian−Toarcian, Chubut Province, Argentina (Ferrari, Reference Ferrari2013).
Description
See original description in Ferrari (Reference Ferrari2013, p. 585).
Materials
MLP 35516; one fragmentary and recrystallized specimen.
Remarks
The material here analyzed certainly belongs to the species Anulifera chubutensis (see Ferrari Reference Ferrari2013, p. 585). Anulifera sigmoidea Ferrari and Damborenea (2015, p. 636, fig. 2.23−31) recently reported from the early Bajocian of the Neuquén province differs from A. chubutensis in being smaller, having a more slender outline shell, a slightly concave mature whorls, and sigmoidal growth lines on the outer face.
For comparisons with other similar species of the genus, see Ferrari (Reference Ferrari2013).
Superfamily Campaniloidea Douvillé, Reference Douvillé1904
Family Ampullinidae Cossmann in Cossmann & Peyrot, Reference Cossmann and Peyrot1919
Genus Globularia Swainson, Reference Swainson1840
Type species
Ampullaria sigaretina Lamarck, Reference de Lamarck1804, from the Eocene of France.
Occurrence
Triassic?/Jurassic−Holocene. Cosmopolitan.
Globularia catanlilensis (Weaver, Reference Weaver1931)

Figure 3 (1−10) Globularia catanlilensis (Weaver, Reference Wenz1931). (1−5) MLP 12171. (1−2, 5) Lateral views; (3) apical view; (4) lateral and apertural views. (6−10) MCF-PIPH 563. (6−9) Lateral views; (10) apical view. (11−21) Naricopsina aequabilis n. sp. (11) MLP 25098, paratype, lateral view. (12−15) MLP 25096, holotype. (12, 13) Lateral view; (14) apertural view; (15) apical view. (16, 17) MLP 35484, lateral views. (18, 19) MLP 35486, lateral views. (20, 21) MLP 27884, lateral views. (22−26) Naricopsina ballentae n. sp., MLP 35494, holotype. (22, 23) lateral views; (24, 25) apical views; (26) lateral and apertural views. (27−31) Pictavia rothi n. sp. (27, 28) MLP 12170. (27) Lateral view; (28) basal and apertural views. (29−31) MLP 12169, paratype. (29, 31) Lateral views; (30) basal and apertural views.
1931 Natica catanlilensis Reference WeaverWeaver, p. 378, pl. 42, figs. 277−278.
1934 Natica aff. catanlilensis, Feruglio, Reference Feruglio1934, p. 49, pl. 5, figs. 2a, b, 3a, b.
1942 Natica catanlilensis Wahnish, Reference Wahnish1942, p. 61, pl. 2, figs. 1a, b.
2013 Globularia cf. catanlilensis (Weaver, Reference Weaver1931); Reference FerrariFerrari, p. 587, figs. 3H, 4A−C.
Holotype
BM 280/1029, from the Early Jurassic of Catán Lil at Southerm Neuquén (Weaver, Reference Weaver1931, p. 378, pl. 42, figs. 277−278). Cast of the holotype: MLP 25017.
Occurrence
Cerro Roth, Carrán Curá, Estancia Santa Isabel and Catán Lil, Sinemurian−Pliensbachian, Neuquén Province, Argentina; Puesto Araya, Pliensbachian, Mendoza Province, Argentina.
Description
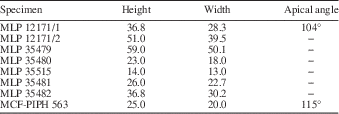
Dextral, globular, anomphalous, naticiform, low-spired and medium- to large-sized shell. The teleoconch has a mean height of 33.9 mm and a mean width of 27.7 mm. The apex forms an angle of 109º. The spire is composed of five strongly convex whorls; last whorl is more expanded than the spire representing the 80% of the shell height. The sub-sutural ramp is narrowly horizontal to slightly convex. The outer face of last whorl is strongly convex in specimen MCF-PIPH 563 and weakly concave in the adapical part of last whorl in specimen MLP 12171. These differences are considered here as intraspecific variability. Sutures are deeply impressed in a concave spiral furrow. The ornament is predominantly spiral and clearly visible on last whorl. The spiral elements consist of very thin and regularly spaced cords; between cords, weak spiral furrows appear. Collabral growth lines are not clearly visible. The base is strongly convex. The umbilicus is closed or partially covered by a columellar callus. The aperture is holostomatous and oval, with the outer lip convex.
Materials
MLP 12171, 35479, 35480, 35481, 35482, 35515; MCF-PIPH 563; seven recrystallized specimens.
Dimensions (mm)
Table 2 Dimensions (mm) of Globularia catanlilensis (Weaver, Reference Weaver1931). −=missing data.
Remarks
Ferrari (Reference Ferrari2013) suggested that the species originally referred to Natica from the Jurassic of South America should no longer be considered as real representatives of Naticidae and thus were transferred by the author to the genus Globularia. In the present paper, the classification of Ferrari (Reference Ferrari2013) is followed.
The material here described agree with the diagnostic characters of Globularia, including a globular, naticiform, low-spired, and convex shell, with weak spiral ribs, and the closed umbilical area.
Globularia catanlilensis (Weaver) was original described as Natica catanlilensis Weaver (Reference Weaver1931, p. 378, pl. 42, figs. 277−278) from the Early Jurassic of Catan-Lil locality, Neuquén Province, Argentina. Natica klingamanni Weaver (Reference Weaver1931; p. 374, pl. 42, figs. 279−280) differs from G. catanlilensis in having a more elongated shell, a narrower spiral angle, a narrower basal angle, a developed collabral ornament with fine growth lines which become sinuous on the outer face of the whorls, and sutures not very distinct. Globularia andina Ferrari and Damborenea (2015, p. 637, fig. 3.1−3.13), from the early Bajocian of the Neuquén Basin, differs from G. catanlilensis in having a more gradate shell outline, a more developed horizontal sub-sutural ramp, very fine prosocline growth lines on the outer face of last whorl but lacking spiral cords, and a columellar lip thickened as a callus. Globularia cf. catanlilensis (Weaver) was originally considered by Ferrari (Reference Ferrari2013) as conespecific to Globularia catanlilensis (Weaver). However, after analyzing in detail the shell characters of both species they share identical morphology and ornament pattern, and Globularia cf. catalilensis is thus retained here as synonymous of G. catanlilensis.
For comparisons with other similar species, see also Ferrari (Reference Ferrari2013, p. 587).
Genus Naricopsina Chelot, Reference Chelot1886
Type species
Lobostoma guarangeri Davoust, Reference Davoust1855, from the Middle Jurassic (Bathonian) of Europe.
Occurrence
Early Jurassic (Sinemurian)−Upper Cretaceous (Maastrichtian). Europe, Africa, Asia, America.
Remarks
In the present paper, the genus Naricopsina is considered as a member of Ampullinidae (see Kaim et al., Reference Kaim, Beisel and Kurushi2004; Ferrari, Reference Ferrari2013). According to Gründel (Reference Gründel2001b, Reference Gründel2003) the representatives of this genus share a naticiform shell, whose spire is depressed to distinct, with the edge of the aperture not thickened and orthocline, with teleoconch whorls convex, without any sculpture or with weak spiral sculpture. The growth lines are partially thickened, more or less prosocline, straight or sinuous, a narrow but distinct umbilicus is developed, the aperture is broadly oval and the inner lip has a callus that sometimes covers the umbilicus. The species described below fits with the characterization of Gründel (Reference Gründel2001b).
Naricopsina aequabilis new species
Holotype
MLP 25096; one recrystallized specimen.
Paratype
MLP 25098; one recrystallized specimen.
Diagnosis
Globose, step-like shell; narrow, horizontal sub-sutural ramp; outer face convex; last whorl more expanded than the spire; ornament lacking; umbilicus widely opened or covered by a callus; aperture oval.
Occurrence
Subida a Sañicó, Estancia Santa Isabel and Arroyo Ñiraico, Pliensbachian−early Toarcian, Neuquén Province; Arroyo Serrucho, Cerro Tricolor, Cañada Colorada, Arroyo El Pedrero, Cerro Puchenque and Puesto Araya, late Sinemurian (F. de Orthechioceras/Paltechioceras) to late Toarcian, Mendoza Province, Argentina.
Description
Dextral, globose, naticiform, medium-sized and low-spired shell. The protoconch is not preserved. The teleoconch is composed of four strongly convex and step- like whorls. The mean height of the teleoconch is 24.4 mm and a mean with is 20.5 mm. The apex forms an angle of 103º, although earliest teleoconch whorls are missing in the available specimens. Teleoconch whorls have a narrow and horizontal sub-sutural ramp; the whorls become strongly convex toward the outer face. Suture is deeply incised in a concave spiral furrow. The shell is externally smooth. The base is convex. The umbilicus is widely open or partially covered by a callus. The aperture is holostomatous and broadly oval with the adapical edge acute and the basal edge convex.
Etymology
Latin, adjective aequabilis=uniform; referring to the uniform ornament pattern and the absence of sinuous collabral ribs on the shell surface, which is a diagnostic character of the genus.
Materials
MLP 12164, 15552a/b, 27884, 35483, 35484, 35485, 35486, 35487, 35488, 35489, 35490, 35491, 35492, 35493; sixteen recrystallized specimens.

Remarks
The material here described is assigned to Naricopsina on the basis of a naticiform shell shape, lacking a distinct sculpture, the teleoconch whorls strongly convex, the aperture broadly oval, and a callus in the inner lip which sometimes covers the umbilicus (see Gründel, Reference Gründel2001b, Reference Gründel2003; Kaim et al., Reference Kaim, Beisel and Kurushi2004).
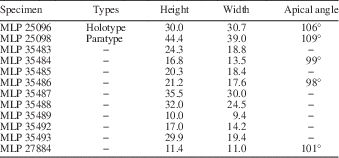
In comparison with other similar Argentinean species, Naricopsina aequabilis n. sp. is very similar to Natica cf. bulimoides (Deshayes), from the Early Cretaceous of the Agrio Formation, Neuquén Province, Argentina (Weaver, Reference Weaver1931, p. 374, pl. 43, figs. 289−290). Both share similar general shell morphology, with a globose shape, spire whorls with a very narrow sub-sutural ramp, aperture oval and the inner lip callused and deflected; however, the species assigned to Natica has a narrower spire angle, is larger than N. aequabilis, and the inner lip is separated from the last whorl by a V-shaped depression. Natica cf. philippi Möricke (Jaworski, Reference Jaworski1926b, p. 196; Weaver, Reference Weaver1931, p. 377), from the Early Jurassic of Argentina, show resemblance with N. aequabilis; however, according to the available descriptions, N. cf. philippi has a spire rather higher than N. aequabilis, an open umbilicus, and a narrower apical angle. Naricopsina laevis (Stoliczka, Reference Stoliczka1861) (Szabó, Reference Szabó2009, p. 98, fig. 91), from the Early Jurassic (Sinemurian) of Europe, resembles N. aequabilis, but, the European species is smaller, and has unequally thin and strongly prosocline growth lines. Naricopsina guerangeri (Davoust, 1885) (Gründel, Reference Gründel2001b, p. 62, pl. 5, figs. 3−7), from the Middle Jurassic of Europe, is comparable to the Argentinean species; however, N. guerangeri has a lower spire and is smaller than N. aequabilis, has orthocline to slightly sinuous and distinctly prosocline growth lines, with their apical part prosocyrt, and fine spiral elements (about 20) on base. Naricopsina montreuilensis (Hébert and Eudes-Deslongchamps, Reference Hébert and Eudes−Deslongchamps1860) (Gründel, Reference Gründel2001b, p. 64, pl. 5, figs. 8−12; Gründel, Reference Gründel2003, p. 186, pl. 14, figs. 8−9, pl. 15, figs. 1−4), from the Middle Jurassic (Callovian) of Europe, differs from N. aequiabilis in being smaller and in having spiral elements on the shell surface. Naricopsina? sinuosa (Gründel, Reference Gründel2001b, p. 65, pl. 6, figs. 1−6; Gründel, Reference Gründel2003, p. 186, pl. 15, figs. 6−9), from the Middle Jurassic (Callovian) of Europe, resembles the species here described, even though the European form is much smaller than the Argentinean species, and has fine and slightly opisthocyrt growth lines near the adapical suture and on the base. The material described by Szabó and Jaitly (Reference Szabó and Jaitly2004, p. 19, pl. 2, figs. 37−39) as Naricopsina cornelia (Laube, Reference Laube1867), from the Callovian (Middle Jurassic) of India, is also comparable to N. aequabilis; the Indian form, however, has a wider apical angle, and irregularly spaced and strongly prosocline growth lines, with weak spiral lines crossing the collabral elements.
Naricopsina ballentae new species
Holotype
MLP 35494; one recrystallized specimens.
Paratypes
MLP 35495, 35496; two recrystallized specimens.
Diagnosis
Globose, naticiform, very low-spired shell; sub-sutural ramp lacking; sutures slightly impressed; outer face convex; ornament lacking; last whorl more expanded than the spire and angular toward base; umbilical area widely opened; aperture circular to oval.
Occurrence
Carrán Curá, late Pliensbachian, Neuquén Province, Argentina.
Description
Dextral, globose, naticiform, medium-sized and low-spired shell. The apex forms an angle of 115°. The protoconch is not preserved. The teleoconch has a mean height of 14.4 mm and a mean width of 18.0 mm, and consists of four strongly convex whorls; last whorl markedly more expanded than the spire. Sub-sutural ramp is missing. Suture is incised in a spiral furrow. The shell is smooth. Last whorl is convex on the outer face and become slightly angular to the periphery. The base is convex with a widely opened umbilical area. The aperture is circular to slightly oval, with the outer lip strongly convex and the columellar lip thickened.
Etymology
Dedicated to Dr. Sara Ballent (1950-2011), renowned micropalaeontologist from Museo de Ciencias Naturales de La Plata (MLP).
Dimensions (mm)
MLP 35494, holotype: height=21.7; width=25.0; apical angle=115°. MLP 35495, paratype: height=6.6; width=11.5. MLP 35496, paratype: height=15.0; width=17.6.
Remarks
Naricopsina ballentae n. sp. differs from Naricopsina aequabilis in having a less step-like shall shape, less impressed sutures, and a lower spire.
Naricopsina montreuilensis (Hébert and Eudes-Deslongchamps, Reference Hébert and Eudes−Deslongchamps1860) (Gründel, Reference Gründel2001b, p. 64, pl. 5, figs. 8−12; Gründel, Reference Gründel2003, p. 186, pl. 14, figs. 8−9, pl. 15, figs. 1−4), from the Middle Jurassic (Callovian) of Europe, is very similar to N. ballentae in shell morphology; however, the European form has a more closed, funnel shaped and narrower umbilical area, a more convex and less angular last teleoconch whorl, fine spiral cords on base and stronger sinuous growth lines. Naricopsina guerangeri (Davoust, Reference Davoust1855) (Gründel, Reference Gründel2001b, p. 62, pl. 5, figs. 3−7), from the Middle Jurassic of Europe, is also comparable to the Argentinean species; however, N. guerangeri has a much lower spire, more convex last teleoconch whorl, and a more oval aperture.
Genus Pictavia Cossmann, Reference Cossmann1925
Type species
Natica pictaviensis d’Orbigny, 1852 (=Natica bajocensis d’Orbigny, 1852) from the Middle Jurassic (Bajocian) of France.
Occurrence
Early Jurassic−Middle Jurassic; Europe, South America.
Remarks
Pictavia shows close resemblance with the representatives of the genus Oonia Gemmellaro, Reference Gemmellaro1878. Gründel (Reference Gründel2001b), however, pointed out that members of Oonia differs from those of Pictavia in lacking a well-developed sub-sutural ramp on teleoconch whorls, and the growth lines (when present) are sinuous and prosocline. In contrast, members of Pictavia show a narrow ramp which may be smooth or ornamented by spiral rows of fine pits, and the growth lines are straight or only weakly sinuous and distinctly prosocline. Based on these characters, in the present paper the classification of Gründel is followed to include the Argentinean material into Pictavia.
Pictavia rothi new species
Holotype
MLP 35497; one recrystallized specimen.
Paratypes
MLP 12169, 15540; two recrystallized specimens.
Diagnosis
Globose to egg-shaped shell; sub-sutural ramp narrowly horizontal; last whorl more expanded than the spire; ornament lacking; base convex; aperture oval with an S-shaped columellar edge; narrow umbilical area.
Occurrence
Cerro Roth and Estación Rajapalo, Pliensbachian, and Arroyo Ñiraico, early Toarcian, Neuquén Province, Argentina.
Description
Dextral, slightly globose to egg-shaped, small-sized, and moderately high-spired shell. Protoconch is not preserved. Teleoconch is composed of four to five convex whorls with a mean height of 24.4 mm and a mean width of 13.0 mm. The apex forms an angle of 85°. Last whorl more expanded than spire whorls. Suture is incised in concave spiral furrow and the sub-sutural ramp is narrowly horizontal. The shell is externally smooth and lacks the typical sinuous and prosocline growth lines.
On the last whorl of specimen MLP 12170, near the outer lip, irregularly spaced axial folds are visible. The base is convex and the aperture oval with an S-shaped columellar edge; adapical edge slightly acute and the abapical edge more expanded. A very narrow umbilical area is developed, which sometimes may be covered by a callus.
Etymology
Dedicated to Dr. Santiago Roth (1850–1924), a renowned Swiss Argentine paleontologist.
Materials
MLP 12170, 15436; two recrystallized specimens.
Dimensions (mm)
MLP 35497, holotype: height=24.9; width=17.6; apical angle=82º. MLP 12169, paratype: height=29.2; width=2.6; apical angle=98º. MLP 15540, paratype: height=23.0; width=13.6; apical angle=76º. MLP 12170: height=20.7; width=18.3.
Remarks
According to Gründel’s (Reference Gründel2001b) diagnosis, the material here described represents members of Pictavia, considering the narrowly developed sutural ramp as the main diagnostic feature (see above).
The type species Natica pictaviensis d’Orbigny (1852, p. 191, pl. 289, figs. 8−10; Fischer and Weber, Reference Fischer and Weber1997, p. 71, pl. 17, figs. 7−8, as Ampullospira [Pictavia] bajociensis), from the Middle Jurassic (Bajocian) of France, is very similar to the Argentinean species; however, the European form has a more slender shell shape, is smaller, has a slightly more developed sub-sutural ramp, and collabral elements.
The analyzed material is also similar to Pictavia calypso (d’Orbigny) (Gründel, Reference Gründel2001b, p. 57, pl. 3, fig. 14, pl. 4, figs. 1−9; Reference Gründel2003, p. 184, pl. 13, figs. 9−10, pl. 14, fig. 1), from the Middle Jurassic (Callovian) of Europe; however, the former species is smaller, and has well developed growth lines on the shell surface, which are intercepted by fine spiral cords. Pictavia punctura (Bean) (Gründel, Reference Gründel2001b, p. 58, pl. 4, figs. 10−15; Reference Gründel2003, p. 185, pl. 14, figs. 2−4), from the Middle Jurassic (Bathonian−Callovian) of Europe, differs from Pictavia rothi n. sp. in having a more developed collabral ornament with fine prosocline to slightly opisthocyrt growth lines, which are more developed on last teleoconch whorl.
Genus Oonia Gemmellaro, Reference Gemmellaro1878
Type species
Melania abbreviata Terquem Reference Terquem1855=Pseudomelania hettangiensis Cossmann Reference Cossmann1909, from the Early Jurassic of France.
Occurrence
Early Jurassic−Middle Jurassic; Europe, South America.
Remarks
As it is pointed out above, Oonia shows extreme resemblance with representatives of Pictavia. The difference between both genera relies in that Oonia lacks a sub-sutural ramp on the outer face of whorls and the growth lines are distinctly sinuous (weakly sinuous in Pictavia) and prosocline. The classification of Gründel (Reference Gründel2001b) is also followed here.
Oonia acuta new species

Figure 4 (1−5) Pictavia rothi n. sp. (1, 2) MLP 15540, paratype, lateral views. (3−5) MLP 35497, holotype, lateral views. (6−14) Oonia acuta n. sp. (6−9) MLP 12159/a, holotype. (6, 7) Lateral views; (8, 9) lateral and apertural views. (10) 12159/b, paratype, lateral view. (11, 12) MLP 35499, lateral views. (13, 14) MLP 35498, lateral views. (15−20) Striactaeonina transatlantica (Behrendsen). (15, 16) MLP 25094a/b, lateral views. (17) MLP 35508, lateral view. (18) MLP 35510, lateral view. (19, 20) MLP 35509, lateral and apertural views. (21−27) Cylindrobullina ovata (Behrendsen, Reference Behrendsen1891). (21) MLP 35514, lateral view. (22−24) MCF-PIPH 546. (22) Lateral view; (23) lateral and apertural views; (24) apical view. (25, 26) MLP 35511, lateral views; (27) MLP 35512, lateral view. (28, 29) Cylindrobullina brevispira n. sp., MLP 35513, holotype. (28) Lateral view; (29) apical view.
?1926 Oonia euspiroides (Gemmellaro, Reference Gemmellaro1878); Jaworski, 1926, p. 200.
?1931 Oonia cf. euspiroides (Gemmellaro, Reference Gemmellaro1878); Weaver Reference Weaver1931, p. 371.
2001a cf. Oonia Reference GründelGründel, p. 59, pl. 5, fig 1−4.
Holotype
MLP 12159/a; one recrystallized specimen.
Paratypes
MLP 12159/b, 35498, 35499; five recrystallized specimens and one negative mould.
Diagnosis
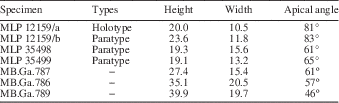
Globose, acute, high-spired shell; juvenile whorls slightly convex; mature whorls strongly convex; last whorls markedly expanded and elongated; spiral ornament predominant on juvenile whorls, opisthocyrt collabral ribs on mature whorls; base convex, aperture elongated and oval; umbilical area covered by a thin columellar callus.
Occurrence
Cerro Roth, Pliensbachian, Neuquén Province; Paso del Portezuelo Ancho, Arroyo Malo and Arroyo Serrucho, late Hettangian to Pliensbachian, Mendoza Province, Argentina. Portezuelo de Pederales and Quebrada Cachina, Early Jurassic (Hettangian−Pliensbachian), Chile (Gründel, Reference Gründel2001a).
Description
Dextral, globose, medium-sized and moderately high-spired shell, with a strongly oval and acute shape. The protoconch is not preserved. The teleoconch has a mean height of 26.3 mm and a mean width of 15.2 mm, and is composed of seven convex whorls. The teleoconch is higher than wide with an apical angle of 65°. The three earliest teleoconch whorls are flat to slightly convex, becoming strongly convex toward the mature growth stages. Last teleoconch whorl is more expanded than spire and elongated toward the abapical edge. Suture is deeply incised in a spiral furrow. The three to four juvenile whorls have a predominant spiral ornament, consisting on weak and fine irregularly spaced cords. On mature whorls, spiral cords become obsolete and opisthocyrtic collabral ribs are visible. The base is convex and the aperture strongly elongated and oval. The columellar lip forms a thin callus and the umbilical canal is deeply impressed. A small siphonal notch is visible on the basal lip in specimen MLP (33).
Etymology
Latin, adjective acuta=acute; referred to the strongly acute shell.
Materials
MB.Ga.786, 787, 789; three recrystallized specimens. Specimens originally assigned by Gründel (Reference Gründel2001a) to cf. Oonia sp., from the Early Jurassic (Hettangian−Pliensbachian) of Chile.
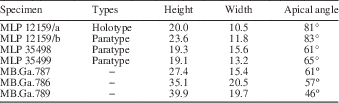
Remarks
Oonia acutovata Gründel (Reference Gründel2001b, p. 60, pl. 4, figs. 16−17, pl. 5, figs. 1−2), from the Middle Jurassic (Bathonian−Callovian) of Europe, differs from O. acuta n. sp. in having a more oval shell and the inner lip forming a narrow callus, and in being smaller.
The genus Oonia was previously reported from Argentina by Jaworski (1926) and Weaver (Reference Weaver1931). Oonia euspiroides (Gemmellaro, Reference Gemmellaro1878) (Jaworski, 1926, p. 200; Weaver, Reference Weaver1931, p. 371 as O. cf. euspiroides), from the Early Jurassic of Portezuelo Ancho (Mendoza, Argentina), shows close resemblance to O. acuta; however, neither Jaworski nor Weaver figured their material. According Gemmellaro’s (Reference Gemmellaro1878) diagnosis, O. euspiroides has well developed growth lines on the shell surface and a narrower spire angle. Considering that O. acuta comes from the same locality (Paso del Portezuelo Ancho) that Oonia euspiroides, here the species described by Jaworski (1926) and Weaver (Reference Weaver1931) are doubtfully included as synonymous of the new species.
Subclass Heterobranchia Gray, Reference Gray1840
Order Architectibranchia Haszprunar, Reference Haszprunar1985 (= Order Opisthobranchia Milne Edwards, Reference Milne-Edwards1848)
Remarks
Gründel and Nützel (Reference Gründel and Nützel2012) pointed out that shells bearing heterobranchs such as modern Acteon or Bullina have been traditionally considered to represent basal opisthobranchs. The authors revised and discussed some Triassic and Jurassic opisthobranch genera, which are related to the modern forms, and based on recent molecular studies they supported the idea that Opisthobranchia is not a monophyletic group (Dinapoli and Klussmann-Kolb, 2010; Schrödl et al., 2011). Thus, they used the name Architectibranchia to include Mesozoic representatives of Opisthobranchia. In the present paper, the classification of Gründel and Nützel (Reference Gründel and Nützel2012) is followed.
Superfamily Acteonoidea d’Orbigny, Reference d’Orbigny1843
Family Tubiferidae Cossmann, 1895
Genus Striactaeonina Cossmann, 1895
Type species
Orthostoma avena Terquem, Reference Terquem1855; Early Jurassic (Hettangian) of Luxembourg.
Occurrence
Early Jurassic−Middle Jurassic; Europe, South America.
Remarks
Based on the characterization of Gründel and Nützel (Reference Gründel and Nützel2012) memebers of Striactaeonina share ‘cylindrical to oval shells, with distinctly elevated spire. The last whorl is moderately convex and not inflated. The whorls have a sub-sutural ramp, which is oblique in most cases. The ramp is demarcated from whorl face by a spiral furrow. Spiral striae are present in whorls flanks and on base. The growth lines are prosocyrt and curving backward in their adapical portion. The aperture is drop- shaped, with the outer lip convex and the inner lip showing a convex parietal and concave columellar portion. Considering these diagnostic characters, the specimens described below are retained into Striactaeonina.
Striactaeonina transatlantica (Behrendsen, Reference Behrendsen1891)
1891 Actaeonina transatlantica Reference BehrendsenBehrendsen, p. 383, pl. 22, fig. 9.
1894 Actaeonina transatlantica Behrendsen, Reference MörickeMöricke, p. 31.
1922 Actaeonina transatraltica Behrendsen; Reference BehrendsenBehrendsen, p. 169, pl. 1, fig. 7.
1926a Striactaeonina transatlantica (Behrendsen); Reference JaworskiJaworski, p. 400.
1926b Striactaeonina transatlantica (Behrendsen); Reference JaworskiJaworski, p. 201.
1931 Striactaeonina transatlantica (Behrendsen); Reference WeaverWeaver, p. 390.
1982 Striactaeonina transatlantica (Behrendsen); Pérez, p. 123, pl. 16, fig. 16.
2001 Striactaeonina transatlantica (Behrendsen); Gründel, p. 65, pl. 6, figs. 10−11.
Type material
Several specimens, originally described by Behrendsen (Reference Behrendsen1891) from the Early Jurassic of Paso del Portezulo Ancho, Mendoza Province, Argentina (Behrendsen, Reference Behrendsen1891, p. 383, pl. 22, fig. 9). Behrendsen’s original material, which should be housed at Göttingen, Germany, could not be examined, and thus no lectotype designation is proposed here.
Occurrence
Cerro Roth, Pliensbachian, Neuquén Province; Arroyo Serrucho, Cerro Puchenque, Paso del Portezuelo Ancho and Arroyo del Portezuelo Ancho, Pliensbachian, Mendoza Province, Argentina.
Description
Dextral, anomphalous, medium-sized and low-spired shell, with an oval to cylindrical shape. The protoconch is not preserved. The teleoconch has a mean height of 31.2 mm and a mean width of 19.2 mm, is strongly stepped in outline and consists of approximately three to four flat to slightly convex whorls. The apex forms an angle of 81º, although earliest whorls are missing in the available specimens. Last teleoconch whorl is more expanded than the spire. Suture is incised and bordered by a weak spiral furrow. The sub-sutural ramp is smooth, horizontal and narrow, and bordered by a peripheral and acute spiral cord. The outer face is flat and becomes slightly convex toward the last whorl and is ornamented by regularly spaced spiral cords; the interspaces between cords develop weak spiral furrows. Collabral ornament is lacking. The base is convex to slightly angular, and the aperture oval.
Materials
MLP 25094a/b, 25095, 35500, 35501, 35502, 35503, 35504, 35505, 35506, 35507, 35508, 35509, 35510; MCF-PIPH 547; 15 recrystallized specimens.
Dimensions (mm)
Table 5 Dimensions (mm) of Striactaeonina transatlantica (Behrendsen, Reference Behrendsen1891). −=missing data.

Remarks
Striactaeonina transatlantica was previously reported in the Early Jurassic of Argentina by several authors (Behrendsen, Reference Behrendsen1891, Reference Behrendsen1922; Möricke, Reference Möricke1894; Jaworski, Reference Jaworski1926a, Reference Jaworskib; Weaver, Reference Weaver1931). Particularly, this species occurs in Pliensbachian deposits of Portezuelo Ancho (Mendoza Province) and Catán Lil (Neuquén Province); but also is present in coeval marine sediments at Qda. Asientos and Jorquera/La Guardia (Chile) (see Pérez, 1982; Gründel, Reference Gründel2001a). Striactaeonina atuelensis Gründel, Reference Gründel2001a (p. 66, pl. 6, figs. 7−8), from the Early Jurassic (early Pliensbachian) of Río Atuel (Mendoza) has recently been revised at the Museum für Naturkunde repository (specimen MB.Ga.803). The species from Chile is very similar to S. transatlantica in general shell morphology and ornament pattern; however, it is slightly smaller than S. transatlantica, has a spire with slightly more convex whorls and a less gradate outline shell, has a narrower sub-sutural ramp, and a wider apical angle of 111°. Actaeonina (Striactaeonina) supraliasica Cox (1965, p. 173, pl. 29, fig. 4a, b, c), from the Early Jurassic (Toarcian) of Kenya, is slightly smaller than the Argentinean form, has a more acute spire and narrower sub-sutural ramp.
Family Cylindrobullinidae Wenz, 1938
Remarks
Gründel and Nützel (Reference Gründel and Nützel2012) argued that Cylindrobullina represents the oldest confirned genus of the family Cylindrobullinidae with it first certain occurrence from the Late Triassic of South America (Haas, Reference Haas1953). Gründel (Reference Gründel2010) redescribed the type species of the genus, Cylindrobullina fragilis (Dunker, Reference Dunker1846), and suggested that the use of the genus had to be changed. Representatives of Cylindrobullina have cylindrical shells with a somewhat elevated spire.
Genus Cylindrobullina von Ammon, Reference von Ammon1878
Type species
Cylindrobullina fragilis (Dunker, Reference Dunker1846), from the Early Jurassic (Hettangian) of North Germany.
Occurrence
Late Triassic−Late Jurassic; Europe, South America.
Remarks
The genus Cylindrobullina shows extreme resemblance with Actaeonia and thus, the assignment of some species to one or the other is usually questionable. Gründel and Nützel (Reference Gründel and Nützel1998) considered Cylindrobullina and Actaeonina as synonymous, giving the priority to Actaeonina. Later, Gründel and Nützel (Reference Gründel and Nützel2012) stated out that the systematic placement of Actaeonina is unclear due to the unknown protoconch of the type species, A. carbonaria (de Koninck, 1843). The authors suggested that the teleoconch morphology of A. carbonaria resembles that of the Mesozoic species of ‘Cylindrites’, including a low spire, very narrow aperture, and a distinct ramp which is demarcated by an angulation from the whorls flank. These similarities suggest that Actaeonina could be indeed the earliest member of Cylindrobullina/Cylindrites group. Representatives of Cylindrobullina share a cylindrical shell shape, the whorls embrace just below the subsutural ramp, the aperture is high and narrow, the spire is low and distinct, whorls are smooth or weakly ornamented with strengthened growth lines, spiral striae or spiral threads on base, growth lines on the flanks weakly prosocyrt and on the ramp directed backward and opisthocyrt, aperture very high, and the protoconch (although unknown from the type species) is heterostrophic (see diagnosis of Gründel, Reference Gründel2010; Gründel and Nützel, Reference Gründel and Nützel2012). These characters are considered here to assign the specimens described below to Cylindrobullina.
Cylindrobullina ovata (Behrendsen, Reference Behrendsen1891) new combination
1891 Actaeonina ovata Reference BehrendsenBehrendsen, p. 383, pl. 22, fig. 7.
2015 Actaeonina ovata Behrendsen, Reference Behrendsen1891; Ferrari, p. 88, fig. 7 R−T.
Type material
One specimen originally described by Behrendsen (Reference Behrendsen1891) from the Early Jurassic of Paso del Portezuelo Ancho, Mendoza Province, Argentina (Behrendsen, Reference Behrendsen1891, p. 383, pl. 22, fig. 7). Behrendsen´s original specimen, which should be housed at Göttingen, Germany, could not be examined.
Emended diagnosis
Oval, egg-shaped shell; teleoconch whorls convex; last whorl greatly expanded; sub-sutural ramp horizontal; prosocyrt collabral ribs; regularly spaced spiral furrows on base.
Occurrence
Cerro Roth, Pliensbachian, Neuquén Province; Cerro Puchenque, Paso del Portezuelo Ancho and Arroyo Serrucho, Pliensbachian, Mendoza Province, Argentina.
Description
Dextral, oval, egg-shaped, medium-sized and low-spired shell. The protoconch is not preserved. The teleoconch is composed of five convex whorls with a mean height of 23.9 mm and a mean width of 13.8 mm. The axis forms an apical angle of 72°. Last teleoconch whorl is greatly more expanded than the spire. The spire is low. The sub-sutural ramp is narrowly horizontal and convex. The outer face of whorls is strongly convex. Suture is impressed in a weak spiral furrow. The shell is smooth, or has prosocyrt collabral ribs on last teleoconch whorl in some specimens. The base is convex with fine and regularly spaced spiral furrows. The aperture elongated and strongly oval, with the posterior end acute and the anterior end expanded. The columellar lip is thin.
Materials
MCF-PIPH 546, MLP 35511, 35512, 35514; four recrystallized specimens.
Dimensions (mm)
MLP 35514: height=17.2; width=8.7; apical angle=54 º. MCF-PIPH 546: height=22; width=13; apical angle=87º. MLP 35511: height=29.2; width=15.8; apical angle=75°. MLP 35512: height=27.2; width=17.8; apical angle=72°.
Remarks
The species was originally described by Behrendsen (Reference Behrendsen1891, p. 383, pl. 22, fig. 7) under the genus Actaeonina, from the Early Jurassic of Portezuelo Ancho (Mendoza Province, Argentina). Here, Behrendsen’s species is reassigned to Cylindobullina, following the updated characterizations of Gründel (Reference Gründel2010) and Gründel and Nützel (Reference Gründel and Nützel2012). Moreover, the new material here described show the typical prosocyrt growth lines on the shell surface and the spiral striae on base. The spiral ornament was not described in the original characterization of Behrendsen, and in the present paper this character is included in the emended the diagnosis of the species.
Cylindrobullina ovata resembles Actaeonina novozealandica Bandel et al. (Reference Bandel, Gründel and Maxwell2000, p. 101, pl. 10, figs. 8−10, 12), from the Early Jurassic of New Zealand; however, the species described by Bandel et al. (Reference Bandel, Gründel and Maxwell2000) is much smaller than C. ovata, has a higher spire shell, a teleoconch whorls with rounded angular corner, a developed spiral groove, and fine growth lines on the shell surface. Actaeonina submoorei Kuhn (Reference Kuhn1936) (in Gründel and Nützel, Reference Gründel and Nützel1998, p. 76, pl. 4, figs. 5−10), from the Early Jurassic (Pliensbachian) of Germany differs from C. ovata in having a more developed sub- sutural ramp, spiral grooves on the angular corner of whorls, and a more acute spire. Actaeonina domeria Gründel and Nützel (Reference Gründel and Nützel1998, p.77, pl. 5, figs. 2−4), Actaeonia obliquata Gründel and Nützel (Reference Gründel and Nützel1998, p.78, pl. 4, fig. 11−12, pl. 5, fig. 1) and Actaeonina rara Gründel and Nützel (Reference Gründel and Nützel1998, p.78, pl. 4, figs. 11−12, pl. 5, fig. 1), all from the Early Jurassic (early Pliensbachian) of Germany, have wider sub- sutural ramps in comparison to A. ovata. Cylindrobullina arduennensis (Piette, Reference Piette1856) (in Gründel and Nützel, Reference Gründel and Nützel2012, p. 36, fig. 2), from the Early Jurassic (Hettangian) of France, is very similar to C. ovata in shell shape, although the European form is much smaller.
Haas (Reference Haas1953) described many Cylindrobullina species from the Late Triassic of the Pucará Group (Central Peru), which are very similar to C. ovata. Cylindrobullina (Cylindrobullina?) pyrulaeformis Haas (Reference Haas1953, p. 266, pl. 17, figs. 67−69, 72−72, 84, 85) and Cylindrobullina (C?) obesa Haas (Reference Haas1953, p. 264, pl. 17, figs. 54−57, 60−63, 65, 66, 70, 71, 75, 76, 79−82, 86) differs from C. ovata in having strongly convex whorls, and more pronounced sutural ramps. Particularly, Cylindrobullina (C.?) pyrulaeformis has a strongly corkscrew- shaped columella and Cylindrobullina (C.?) obesa has a slightly concave base; these characters are not visible in A. ovata.
Cylindrobullina brevispira new species
Holotype
MLP 35513; one recrystallized specimen.
Diagnosis
Egg-shaped; extremely low spire for the genus; sub-sutural ramp widely horizontal; last whorls greatly expanded; ornament lacking; aperture elongated and oval.
Occurrence
Arroyo Serrucho, Pliensbachian, Mendoza Province, Argentina.
Description
Dextral, oval, egg-shaped, medium-sized and very low-spired shell. The protoconch is not preserved. The teleoconch is composed of five convex whorls and has a height of 25.0 mm and a width of 17.4 mm. The axis forms an apical angle of 120°. Last teleoconch whorl is greatly more expanded than spire. The sub-sutural ramp is widely horizontal and forms a rounded edge as transition with last whorls; the outer face of whorls is strongly convex. Suture is impressed in a weak spiral furrow. The shell is smooth and the base convex. The aperture is elongated and strongly oval, with the posterior end acute.
Etymology
Referred to the extremely low-spired shell in comparison with other representatives of the genus.
Dimensions (mm)
MLP 35513, holotype: height=25; width=17.4; apical angle=120°.
Remarks
Cylindrobullina brevispira n. sp. shows a more obtuse apical angle and a more convex whorls outline than Cylindrobullina ovata. Cylindrobullina brevispira is also comparable to the type species Cylindrobullina fragilis (Dunker, Reference Dunker1846), from the Early Jurassic (Hettangian) of North Germany; however, the European form has a more elevated spire and clearly visible opisthocyrt collabral ribs on the outer face of last whorl. Cylindrobullina arduennensis (Piette, Reference Piette1856) (in Gründel and Nützel, Reference Gründel and Nützel2012, p. 36, fig. 2), from the Early Jurassic (Hettangian) of France, is similar to C. brevispira, although the European form is much smaller, has a less convex last whorl with opistocyrt collabral ribs, and a slightly more elevated spire.
Acknowledgments
I am deeply grateful to S. Damborenea (MLP) for providing most of the specimens described here, which were collected by her from 1975 onwards during field trips to the Neuquén Basin accompanied by M. Manceñido and A. Riccardi (both at MLP) to whom I also extend my gratitude. I thank A. Riccardi for allowing access to the MLP collection and for supplying an updated biostratigraphical framework for the Neuquén Basin. I also thank the Dirección General de Patrimonio Cultural, Secretaría de Estado de Cultura de la Provincia de Neuquén, and L. Zingoni, who allowed the access to outcrops in his land in southern Neuquén Province. I am also grateful to R. Coria (Museo Carmen Funes, Plaza Huincul, Neuquén, Argentina) for arranging the loan of the gastropod material collected by the author. I extend my gratitude to S. Bessone (CENPAT), L. Canessa (MEF), and N. Pfeiffer (MEF) for preparation of specimens; and to M. Manceñido and J. Echevarría (MLP) for their help during fieldwork in the Neuquén Basin in 2012. I thank S. Lanés for providing specimens from Arroyo El Pedrero and J. Groizard (ALUAR) for allowing access to the scanning electron microscope. Study of the MLP and MCF-PIPH gastropod material was made possible by a Sylvester-Bradley Award from the Palaeontological Association. The revision of the MB.Ga. gastropod material was made posible in part by a project financed by AGENCIA being PICT 2013-0356. I thank M. Aberhan (Museum für Naturkunde, Berlin) for allowing the access to the MB.Ga. invertebrate collection. This study is also part of a long-term project financed by CONICET grants, the last one being PIP 112-200801-01567. Finally, I extend my gratitude to R. Gatto (Dipartimento di Geoscienze, Università di Padova, Italy) and A. Beu (GNS, New Zealand) for their valuable comments, which helped to greatly improve the manuscript.