



Introduction
Phorusrhacidae Ameghino, Reference Ameghino1889 is an extinct group of flightless, cursorial carnivorous birds that occupied one of the dominant, large land-predator niches in South America from the lower Eocene to the Pleistocene (Tonni and Tambussi, Reference Tonni and Tambussi1986; Tambussi et al., Reference Tambussi, Ubilla and Perea1999; Alvarenga et al., Reference Alvarenga, Jones and Rinderknecht2010; Degrange, Reference Degrange2017; Jones et al., Reference Jones, Rinderknecht, Alvarenga, Montenegro and Ubilla2018). They dispersed into North America during the Great American Biotic Interchange (~3 Ma; Marshall, Reference Marshall, Hecht, Goody and Hecht1977; Carr, Reference Carr1981; Baskin, Reference Baskin1995; Alvarenga and Höfling, Reference Alvarenga and Höfling2003; MacFadden et al., Reference MacFadden, Labs-Hochstein, Hulbert and Baskin2007). Some remains from Africa and Europe (Mourer-Chauviré et al., Reference Mourer-Chauviré, Tabuce, Mahboubi, Adaci and Bensalah2011; Angst et al., Reference Angst, Buffetaut, Lecuyer and Amiot2013) and the Paleocene of Brazil (Alvarenga, Reference Alvarenga1985) have been referred to this clade or identified as phylogenetically related to the extant South American seriemas (Cariamidae), but these assignments remain controversial (Agnolín, Reference Agnolín2009; Degrange et al., Reference Degrange, Tambussi, Taglioretti, Dondas and Scaglia2015; Mayr, Reference Mayr2017).
With an estimated body mass of more than 100 kg (Degrange, Reference Degrange2012; Degrange et al., Reference Degrange, Noriega, Areta, Vizcaíno, Kay and Bargo2012), Phorusrhacos longissimus Ameghino, Reference Ameghino1887 is one of the most iconic fossil birds from South America and part of the early history of paleontology in Argentina. Discovered by Carlos Ameghino in Santa Cruz province, it was described by Florentino Ameghino from an incomplete mandible originally assumed to be from a toothless mammal (Ameghino, Reference Ameghino1887; Alvarenga and Höfling, Reference Alvarenga and Höfling2003; Buffetaut, Reference Buffetaut2013a, Reference Buffetaut, Göhlich and Krohb). Unfortunately, no illustration accompanied the original description (see Ameghino, Reference Ameghino1887).
The avian nature of Phorusrhacos was first proposed in 1891 on the basis of comparisons with new remains of the related species Patagornis marshi Moreno and Mercerat, Reference Moreno and Mercerat1891 (= Tolmodus inflatus according to Ameghino, also originally described as a toothless mammal, Ameghino, Reference Ameghino1891a). Moreno and Mercerat (Reference Moreno and Mercerat1891) figured the Phorusrhacos holotype mandible for the first time (MLP 20-118, Fig. 1; see Acosta Hospitaleche et al., Reference Acosta Hospitaleche, Tambussi and Reguero2001) but erroneously proposed it to be the premaxilla. A subsequent discovery of a skull of P. longissimus (NHMUK A529) by Carlos Ameghino could only be described and illustrated (Fig. 2) while it remained in its original sediment matrix in the field, as the specimen was “in such a bad state of conservation that it [was] almost reduced to dust” (Ameghino, Reference Ameghino1895, p. 268). Fossil material recovered and deposited from this specimen represents only a small portion of the skull figured by Ameghino (Reference Ameghino1895), preserving only the rostral tip of the maxilla, fragments of the processus supraorbitalis of the os lacrimale, and part of the mandible (Alvarenga and Höfling, Reference Alvarenga and Höfling2003). The illustrations of this specimen of P. longissimus have been widely reproduced in textbooks and museums, although a lack of new skull material referable to the taxon has precluded the verification of the morphology illustrated.
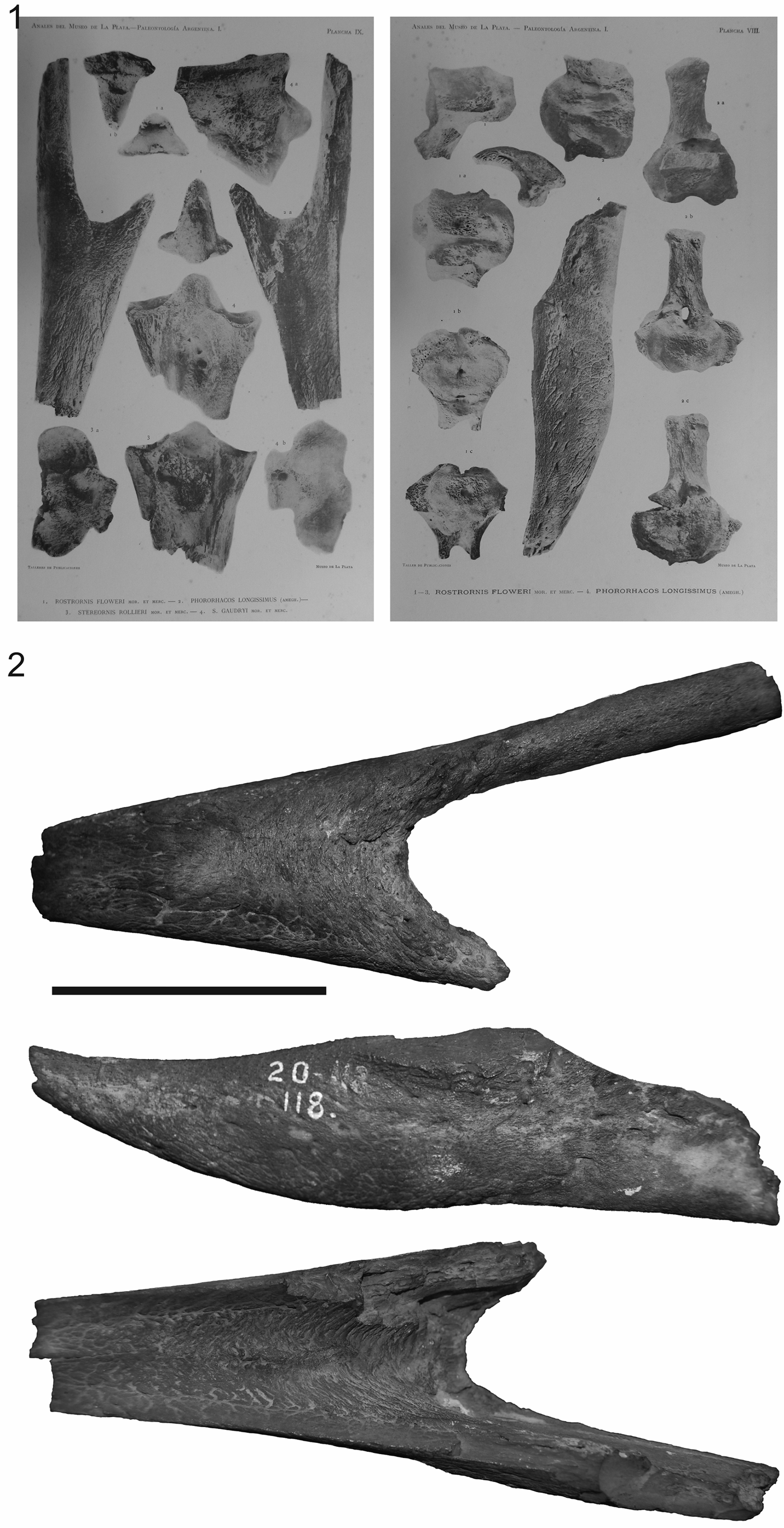
Figure 1. Phorusrhacos longissimus holotype MLP 20-122: (1) as figured by Moreno and Mercerat (Reference Moreno and Mercerat1891); (2) photographs in dorsal, ventral, and lateral views. Scale bar = 1 cm.

Figure 2. (1) The Phorusrhacos longissimus specimen discovered by Carlos Ameghino and drawn by Florentino Ameghino (Reference Ameghino1895) according to the descriptions, sketches, and measurements taken by his brother in the field. (2) Photographs of the jaw of that specimen, numbered NHMUK A529, in right lateral, ventral, and dorsal views. Other remains of the same specimen are a fragment of the rostral tip of the maxilla and processus supraorbitalis of the os lacrimale figured by Alvarenga and Höfling (Reference Alvarenga and Höfling2003, fig. 24). Scale bar = 5 cm.
The Phorusrhacinae subclade within Phorusrhacidae currently includes Phorusrhacos longissimus, Titanis walleri Brodkorb, Reference Brodkorb1963, Devincenzia pozzi Kraglievich, Reference Kraglievich1931, and Kelenken guillermoi Bertelli et al., Reference Bertelli, Chiappe and Tambussi2007. Although recent phylogenetic analyses have recovered different relationships among these species and with the rest of phorusrhacids (e.g., Agnolín, Reference Agnolín2009; Alvarenga et al., Reference Alvarenga, Chiappe, Bertelli, Dyke and Kaiser2011; Degrange et al., Reference Degrange, Tambussi, Taglioretti, Dondas and Scaglia2015), none of the analyses recovered Phorusrhacinae as a natural group. Phorusrhacinae represent fossil birds with gigantic proportions (e.g., body masses greater than 100 kg; Alvarenga and Höfling, Reference Alvarenga and Höfling2003; Bertelli et al., Reference Bertelli, Chiappe and Tambussi2007; Degrange, Reference Degrange2012), long and narrow mandibular symphyses more than twice as elongate as the width at their base (Alvarenga and Höfling, Reference Alvarenga and Höfling2003), and elongated tarsometatarsi (Angst et al., Reference Angst, Buffetaut, Lecuyer and Amiot2015; Degrange, Reference Degrange2017). The majority of these large-bodied phorusrhacid taxa are known from fragmentary skulls and postcrania, with the exception of a nearly complete skull of Kelenken guillermoi (Bertelli et al., Reference Bertelli, Chiappe and Tambussi2007). This contrasts sharply with the fossil record of smaller and medium-sized phorusrhacids, which is far more complete. An array of nearly intact crania as well as associated postcranial material have been referred to taxa within these clades (i.e., Mesembriornithinae, Patagornithinae, and Psilopterinae; Andrews, Reference Andrews1889; Sinclair and Farr, Reference Sinclair, Farr and Scott1932; Kraglievich, Reference Kraglievich1946; Alvarenga and Höfling, Reference Alvarenga and Höfling2003; Degrange et al., Reference Degrange, Tambussi, Taglioretti, Dondas and Scaglia2015). Reconstructions of larger phorusrhacine skulls have been based on these smaller-bodied phorusrhacid taxa (Bertelli et al., Reference Bertelli, Chiappe and Tambussi2007), while the paucity of cranial material continues to limit morphological comparisons within Phorusrhacinae with other phorusrhacids.
Here we present new cranial material referable to the Phorusrhacinae Phorusrhacos longissimus comprising a skull roof, rostrum, and partial mandible (MPEF-PV 11356). A thoracic vertebra (MPEF-PV 11355) is also described and tentatively referred to Phorusrhacinae. It is hypothesized to belong to the same taxon due to its size and morphological similarities when compared with other remains of Phorusrhacos. Two sites in southern Argentina produced the phorusrhacid material. Both localities (Fig. 3.1) are situated in Santa Cruz Province. The vertebra was discovered as an isolated element at the Puesto Estancia La Costa locality, in sediments typical of the Santa Cruz Formation. The associated cranium and mandible were discovered at the Cerro de los fósiles site, less than 50 km east of the Chilean border, approximately 20 km southeast of Lago Belgrano and slightly lower in section than the vertebra. Santacrucian units in this region are constrained to a late early Miocene age based on comparisons of faunal composition to other early Miocene formations in Argentina dated to the Santacrucian Land Mammal Age (~22.0–16.0 Ma; Marshall, Reference Marshall1976; Marshall et al., Reference Marshall, Drake, Curtis, Butler, Flanagan and Naeser1986; Fleagle et al., Reference Fleagle, Perkins, Heizler, Nash, Bown, Tauber, Dozo, Tejedor, Vizcaíno, Kay and Bargo2012; Perkins et al., Reference Perkins, Fleagle, Heizler, Nash, Bown, Tauber, Dozo, Vizcaíno, Kay and Bargo2012).
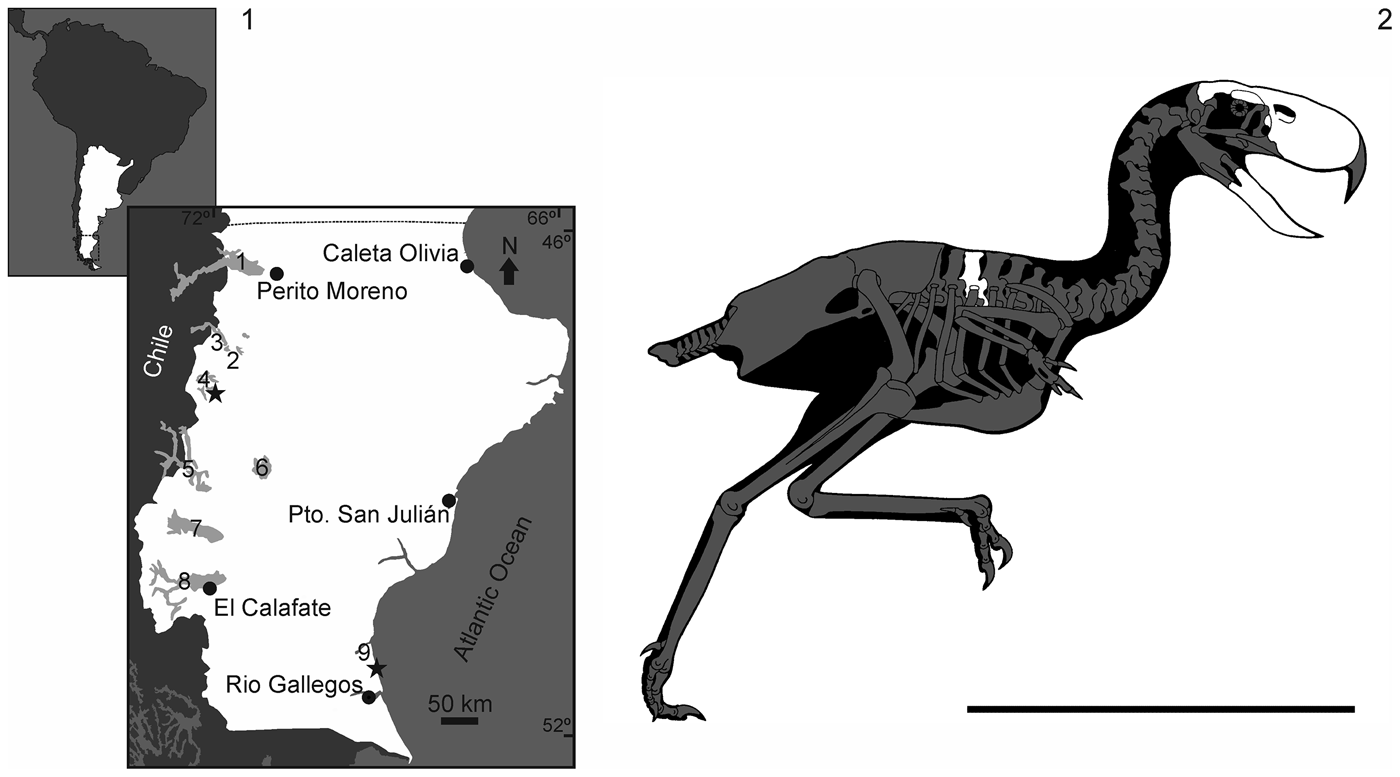
Figure 3. (1) Location map of the fossil locality (star), near Lago Belgrano, at southern Patagonia, Argentina. (2) Reconstruction of Phorusrhacos longissimus skeleton with the preserved bones in white. Scale bar = 1 m. 1 = Lago Buenos Aires; 2 = Lago Posadas; 3 = Lago Pueyrredón; 4 = Lago Belgrano; 5 = Lago San Martín; 6 = Lago Cardiel; 7 = Lago Viedma; 8 = Lago Argentino; 9 = Puesto Ea. La Costa. Reconstruction of Phorusrhacos is modified from Paul (Reference Paul2002).
Materials and methods
Unless indicated otherwise, the osteology terminology used in the anatomical description follows Baumel and Witmer (Reference Baumel, Witmer, Baumel, King, Breazile, Evans and Berge1993) or their English equivalent. Jaw muscles are discussed following Holliday and Witmer (Reference Holliday and Witmer2007).
Repositories and institutional abbreviations
BAR, Museo Asociación Paleontológica Bariloche, Río Negro, Argentina; DGM, Divisão de Geologia e Mineralogia do Departamento Nacional da Produção Mineral, Rio de Janeiro, Brazil; FM, Field Museum of Natural History, Chicago, United States of America; MLP, Museo de La Plata, La Plata, Argentina; MNHN, Muséum National d'Histoire Naturelle, Paris, France; MPEF-PV, Museo Egidio Feruglio, Trelew, Chubut, Argentina; MPM-PV, Museo Padre Molina, Río Gallegos, Santa Cruz, Argentina; NHMUK, The Natural History Museum, London, United Kingdom.
Systematic paleontology
Class Aves Linnaeus, Reference Linnaeus1758
Infraclass Neognathae Pycraft, Reference Pycraft1900
Order Cariamiformes Verheyen, Reference Verheyen1957
Family Phorusrhacidae Ameghino, Reference Ameghino1889
Subfamily Phorusrhacinae Ameghino, Reference Ameghino1889
Genus Phorusrhacos Ameghino, Reference Ameghino1887
Type species
Phorusrhacos longissimus Ameghino, Reference Ameghino1887 from Santa Cruz Formation (late early Miocene), Santa Cruz Province, Argentina.
Phorusrhacos longissimus Ameghino, Reference Ameghino1887
Figures 3–7
- Reference Ameghino1889
Phororhacos longissimus Ameghino, p. 24.
- Reference Moreno and Mercerat1891
Stereornis rollieri Moreno and Mercerat, p. 45, pl. 9, fig. 3, pl. 10, figs. 1, 2, pl. 11, fig. 1.
- Reference Moreno and Mercerat1891
Stereornis gaundryi Moreno and Mercerat, p. 47, pl. 9, fig. 4, pl. 10, fig. 3.
- Reference Moreno and Mercerat1891
Mesembriornis studeri Moreno and Mercerat, p. 48, pl. 4, figs. 2, 3, pl. 7, fig. 4, pl. 10, fig. 4, pl. 11, figs. 2–4, pl. 12, figs. 1–6, lam. 2, fig. B.
- Reference Moreno and Mercerat1891
Mesembriornis quatrefragesi Moreno and Mercerat, p. 50, pl. 18, figs. 1–6, pl. 16, fig. 3.
- Reference Moreno and Mercerat1891
Darwinornis copei Moreno and Mercerat, p. 60, pl. 17, figs. 1, 2.
- Reference Moreno and Mercerat1891
Darwinornis zittelli Moreno and Mercerat, p. 63, pl. 17, figs. 3, 4.
- Reference Moreno and Mercerat1891
Darwinornis socialis Moreno and Mercerat, p. 63, pl. 17, fig. 5.
- Reference Moreno and Mercerat1891
Owenornis affinis Moreno and Mercerat, p. 64, pl. 17, fig. 6, pl. 18, fig. 1.
- Reference Moreno and Mercerat1891
Owenornis lydekkeri Moreno and Mercerat, p. 64, pl. 18, figs. 2–5.
- Reference Ameghino1891a
Phororhacos shenensis Ameghino, p. 258.
- Reference Ameghino1891b
Phororhacos sehuensis Ameghino, p. 451.
- Reference Ameghino1891b
Phororhacos platygnathus Ameghino, p. 452.
- Reference Mercerat1893
Titanornis mirabilis Mercerat, p. 5.
- Reference Ameghino1895
Callornis giganteus Ameghino, p. 574, figs. 28, 29.
- Reference Ameghino1901
Eucallornis giganteus Ameghino, p. 78.
- Reference Dolgopol de Saez1927
Liornis minor Dolgopol de Saez, p. 158, pl. 4.
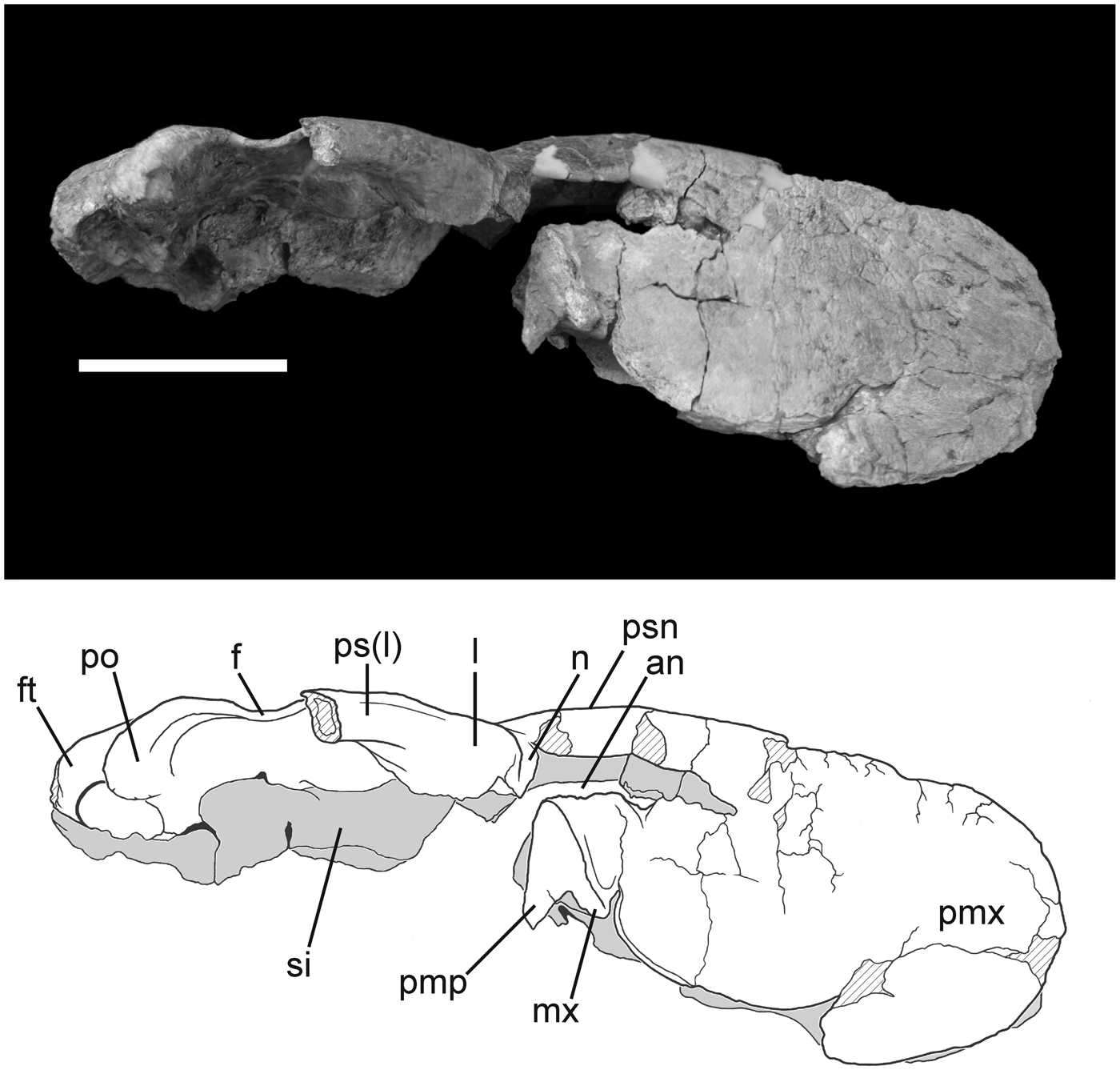
Figure 4. Photograph and interpretive drawing of the skull of Phorusrhacos longissimus MPEF-PV 11356 in right lateral view. an = aperture nasi ossea; f = os frontale; ft = fossa temporalis; l = os lacrimale; mx = os maxillare; n = os nasale; pmp = processus maxillopalatinus; pmx = os premaxillare; po = processus postorbitalis; ps(l) = processus supraorbitalis of the os lacrimale; psn = pila supranasalis; si = septum interorbitale. Scale bar = 10 cm.
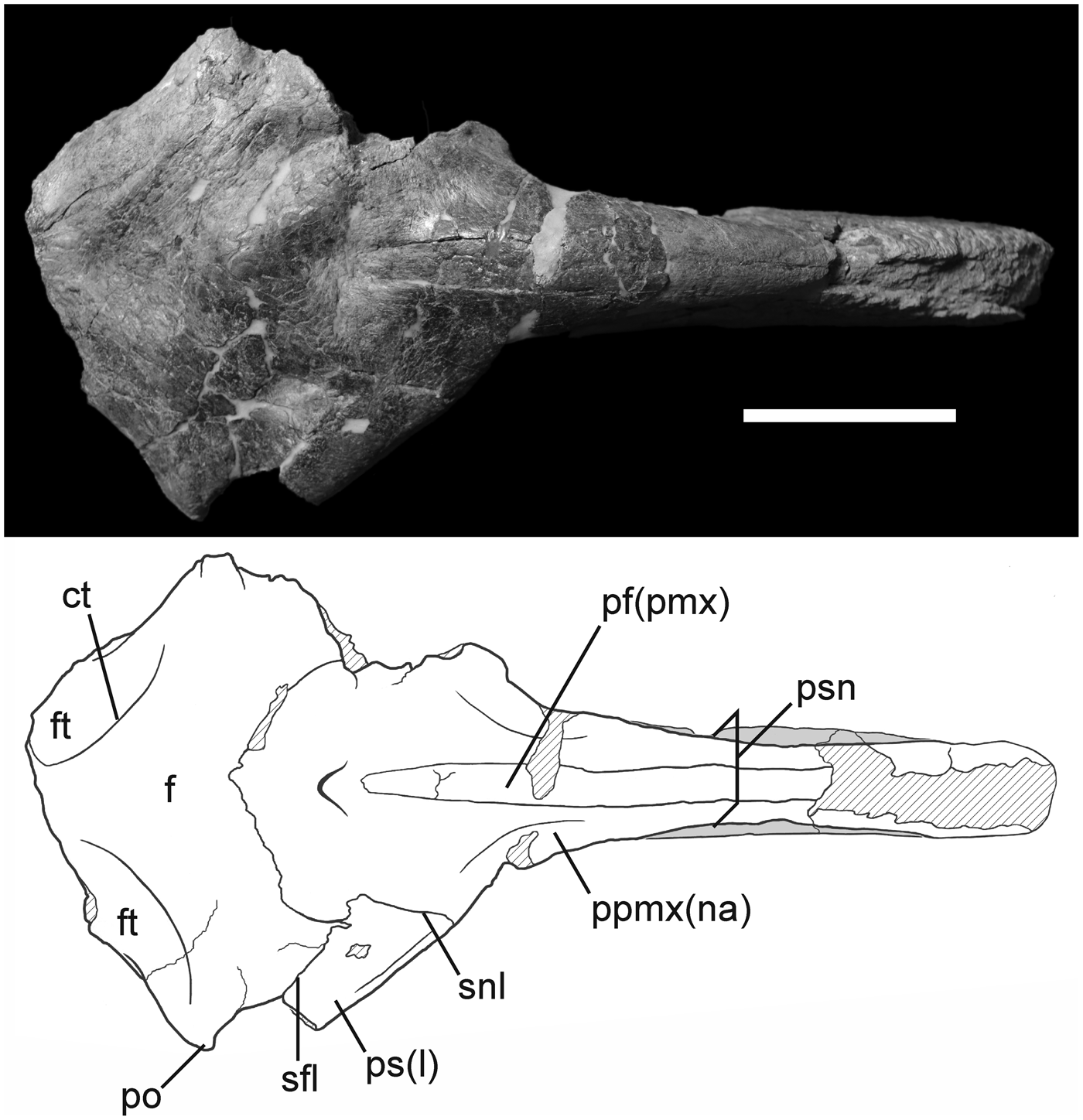
Figure 5. Photograph and interpretive drawing of the skull of Phorusrhacos longissimus MPEF-PV 11356 in dorsal view. ct = crista temporalis; f = os frontale; ft = fossa temporalis; pf(pmx) = processus frontalis of the os premaxillare; po = processus postorbitalis; ppmx(na) = processus premaxillaris of the os nasale; ps(l) = processus supraorbitalis of the os lacrimale; psn = pila supranasalis; sfl = sutura frontolacrimalis; snl = sutura nasolacrimalis. Scale bar = 10 cm.
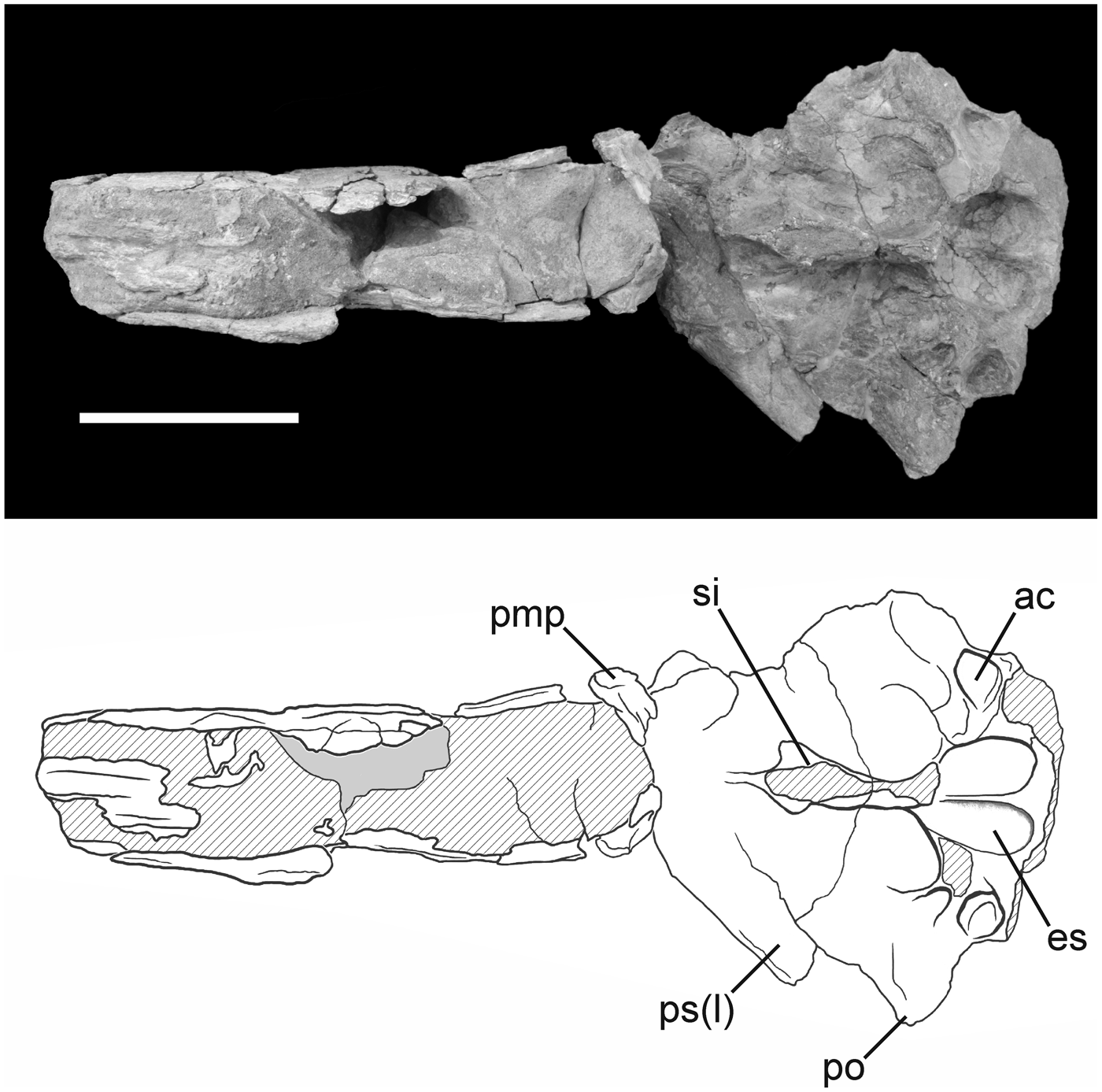
Figure 6. Photograph and interpretive drawing of the skull of Phorusrhacos longissimus MPEF-PV 11356 in ventral view. ac = accessory cotyla for m. adductor mandibulae externus profundus attachment; es = eminentia sagittalis; pmp = processus maxillopalatinus; po = processus postorbitalis; ps(l) = processus supraorbitalis of the os lacrimale; si = septum interorbitale. Scale bar = 10 cm.
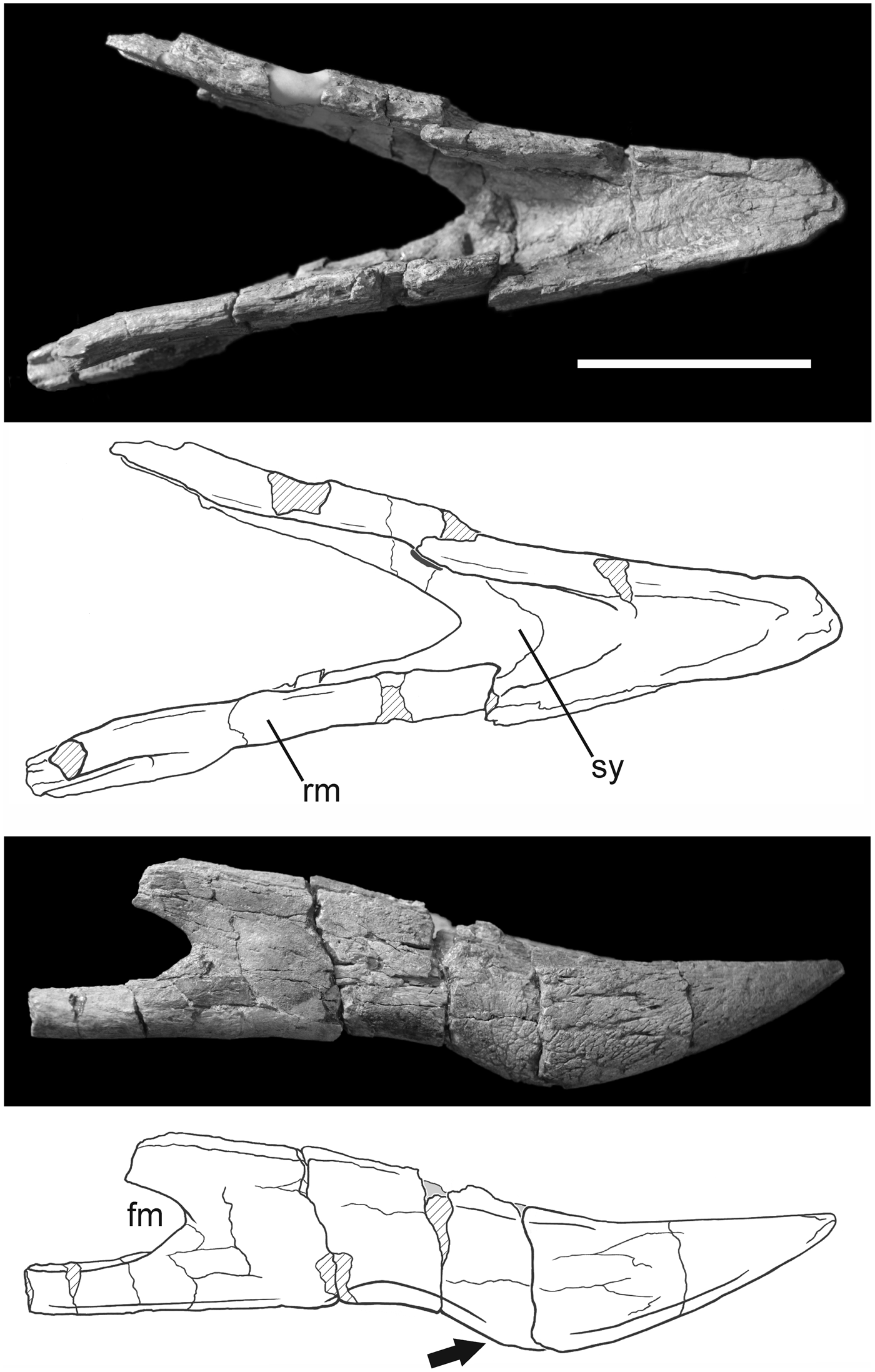
Figure 7. Photograph and interpretive drawing of the jaw of Phorusrhacos longissimus MPEF-PV 11356 in dorsal and right lateral views. fm = fenestramandibulae; rm = ramus mandibulae; sy = symphysis mandibulae. The arrow points to the ventral extension of the caudal portion of the symphysis. Scale bar = 10 cm.
Type specimen
Lower jaw (MLP 20-122) from Santa Cruz Formation (late early Miocene), Santa Cruz Province, Argentina (Moreno and Mercerat, Reference Moreno and Mercerat1891, pl. 8, fig. 4, pl. 9, fig. 2).
Occurrence
Cerro de los fósiles locality, NW of Santa Cruz Province, Argentinean Patagonia, santacrucian SALMA (late early Miocene) of Santa Cruz Formation.
Description
Cranial material of MPEF-PV 11356 (Figs. 4–6) is represented by a large part of the rostrum (including the ossa premaxillaris, ossa nasalis, and a small portion of the right os maxillare) and part of the skull roof and orbit (right os lacrimale, ossa frontalia, and ossa parietalia). Although most of the braincase (e.g., occipital region) and palatal elements are not preserved in MPEF-PV 11356, the internal surface of the dorsal cranium is in good condition, preserving the outline of the rostrodorsal portion of the brain cavity (Fig. 6). The dorsal surface of the rostrum is abraded along most of its length (Fig. 5), and damage continues along the curved rostral surface of the premaxilla. The shape of the bill tip has not been preserved, but it can be inferred from preserved remnants of the ventral margin that the typical downturned hook was present as in other Phorusrhacos specimens (e.g., NHMUK A535).
The rostrum of MPEF-PV 11356 is dorsoventrally tall (14.1 cm at its preserved midpoint, with an estimated maximum height at midlength of the nares of 17.3 cm) and narrow (~6 cm width) with the thick, heavy premaxillary construction typical of larger phorusrhacids. It is proportionally shorter (estimated length of 52 cm) relative to skull length than illustrations of the skull referred to Phorusrhacos longissimus by Ameghino (Reference Ameghino1895; 65 cm) and proportionally shorter than that of the phorusrhacine Kelenken guillermoi (71.6 cm according to Bertelli et al., Reference Bertelli, Chiappe and Tambussi2007). The shape of the rostrum of MPEF-PV 11356 most closely resembles those of the smaller patagornithines Patagornis marshi Moreno and Mercerat, Reference Moreno and Mercerat1891 (NHMUK A516) and Andalgalornis steulleti (Kraglievich, Reference Kraglievich1931) (FM-P14357). The lateral surfaces of the rostrum are characterized by an extensive meshwork of grooves and pits (i.e., neurovascular foramina). These small channels served as exits for the ophthalmic and nasopalatine nerves (Bertelli et al., Reference Bertelli, Chiappe and Tambussi2007), and their presence is widespread throughout Phorusrhacidae (e.g., Degrange and Tambussi, Reference Degrange and Tambussi2011; Degrange et al., Reference Degrange, Tambussi, Taglioretti, Dondas and Scaglia2015).
The tomium of the premaxilla is mostly broken, and a small portion of the os maxillare is visible as an irregularly shaped, paired element surrounded by matrix (Fig. 4). Rostrally, the os maxillare follows the preserved outline of the rostrum and curves dorsally, where it would have contacted the os premaxillare. MPEF-PV 11356 is interpreted to have a slightly upturned ventral margin of the rostrum immediately caudal to its beak tip, which contrasts with that of Kelenken in which the tomium is almost straight (although this could be the result of the dorsoventral crushing present in the holotype). This feature, characterized by a falcated tomia, is typical of the ‘Terror Bird’ skull type (Degrange, Reference Degrange2012) and is present in Patagornithinae and most Phorusrhacinae and contrary to the Psilopterinae type, in which the tomial margin is nearly straight. Upturning of the ventral premaxillary margin is exaggerated in the reconstructed Mesembriornithinae Mesembriornis milneedwardsi Moreno, Reference Moreno1889 (see Kraglievich, Reference Kraglievich1946) and in the illustration of the skull of the phorusrhacine Phorusrhacos longissimus (Ameghino, Reference Ameghino1895).
A sutural contact between the processus premaxillaris of the os nasale and the processus frontalis of the os premaxillare is visible in dorsal view and extends caudal to the opening of the apertura nasi ossea (Fig. 5). The nasopremaxillary contact becomes difficult to trace rostrally due to breakage and the fusion between both bones typical of phorusrhacids (Degrange, Reference Degrange2012); however, holorhiny can be confirmed in Phorusrhacos (Ameghino, Reference Ameghino1895). The pila supranasalis delimits a bow shape when viewed laterally that is not as marked as in Llallawavis or Patagornis. This condition is not observed in Kelenken, although absence could be a consequence of the deformation in the latter. The morphology of the external naris is somewhat obscured by breaks surrounding the opening on both sides of the skull. Only the dorsal margin of the external naris is well defined in MPEF-PV 11356. The general shape of the external naris is inferred to be rostrocaudally elongated, similar to that observed in Kelenken guillermoi (Bertelli et al., Reference Bertelli, Chiappe and Tambussi2007), although the dorsal surface of the nares seems straight in Kelenken and curved in MPEF-PV 11356. This suggests that the preserved shape of the naris in Kelenken guillermoi (BAR 3877-11) may not be an artifact of dorsoventral crushing of the skull, although it appears to be shorter in Phorusrhacos. The external naris illustrated for Phorusrhacos longissimus (Ameghino, Reference Ameghino1895) is more squared in appearance, as seen in the patagornithines Andalgalornis steulleti and Patagornis marshi.
Caudoventral to the external naris, the os premaxillare flares laterally to its contact with the processus maxillaris of the os nasale, which is not clearly distinguishable in MPEF-PV 11356. Rostral to this contact point, the premaxilla exhibits a shallow, bowl-shaped depression that opens to accommodate an accessory fossa of the external naris. It is unclear from Ameghino's (Reference Ameghino1895) illustrations whether early specimens of Phorusrhacos longissimus preserved this depression, although it is present (although shallower) in Kelenken guillermoi and the patagornithines Andalgalornis steulleti and Patagornis marshi. The smaller Psilopterinae also preserves a large accessory fossa to the external naris (Sinclair and Farr, Reference Sinclair, Farr and Scott1932; Agnolín, Reference Agnolín2009; Degrange and Tambussi, Reference Degrange and Tambussi2011). The depression angles rostroventrally and is proportionally smaller in MPEF-PV 11356 than in Psilopterinae. Although most of the ventral margin of the external naris is broken in MPEF-PV 11356, the narial floor is preserved and slopes caudoventrally. At the base of the curved surface of the narial floor, two small, broken processes slope ventromedially, preserving either the rostroventral portion of the maxillary process of the os nasale or part of the processus maxillopalatinus.
The skull roof of MPEF-PV 11356 is dorsoventrally flattened, wide (estimated width of 24 cm, calculated by doubling the distance between the right postorbital process and the sagittal plane of the skull), and would have been roughly triangular in shape when intact (Fig. 5). A shallow depression marks the caudal extent of the nasopremaxillary contact and is bordered by a small, raised knob directly caudal that is crossed by the nasofrontal suture. The wide nasofrontal contact curves rostrolaterally to meet the os lacrimale, at which point the suture divides into nasolacrimal contact rostrolaterally and frontal-lacrimal contact caudolaterally (suturae lacrimofrontalis et lacrimonasalis). The nasolacrimal contact meets the lateral margin of the orbit, and breakage along the ventral margin of the os lacrimale demarcates the missing processus orbitalis of the os lacrimale. In lateral view, the processus supraorbitalis of the os lacrimale forms a thick orbital brow that comprises the lateral borders of the skull roof, although this feature is broken caudally on the right side of the skull of MPEF-PV 11356. Phorusrhacos longissimus shares this prominent processus supraorbitalis with the patagornithines Patagornis marshi and Andalgalornis steulleti, meanwhile this feature is comparatively reduced in the os lacrimale of Psilopterinae. Nevertheless, the tip of this process seems separated from the orbital rim in MPEF-PV 11356, as in Devincenzia pozzi (Cabrera, Reference Cabrera1939). This condition is not seen in Patagornitinae, in which the processus supraorbitalis is in contact with the orbital rim through its entire length. According to Bertelli et al. (Reference Bertelli, Chiappe and Tambussi2007), in the phorusrhacinae Kelenken the contact between the processus supraorbitalis of the os lacrimale and the orbital rim of the os frontale is not clear. However, they also state that “The supraorbital ossification of Kelenken fits within a socket formed by the portion of the frontal that forms the postorbital process” (Bertelli et al., Reference Bertelli, Chiappe and Tambussi2007, p. 402). So it seems likely that in Kelenken the processus supraorbitalis of the os lacrimale is fused with the os frontale, forming the orbital rim. This condition represents an exclusive feature of Kelenken, different from any other Phorusrhacidae. The processus supraorbitalis runs approximately parallel to the roof of the cranium in MPEF-PV 11356 (Figs. 4, 5). An illustration of the skull of Phorusrhacos longissimus, NHMUK A529 (Ameghino, Reference Ameghino1895), shows this process slightly caudoventrally deflected relative to the plane of the skull roof. It is unclear whether this morphology is due to distortion. It is not present in MPEF-PV 11356.
The processus postorbitalis forms a point that constitutes the greatest width of the preserved skull of MPEF-PV 11356 in dorsal view. However, the missing parietal region was likely wider, as in other phorusrhacines. Although the left processus postorbitalis is broken, the right is deflected only slightly ventrally. This is not the condition in Devincenzia and Kelenken, in which the processus is more deflected ventrally or in patagornithines and psilopterines, in which a strong ventral deflection is present. The lateralmost point of the postorbital serves as the origin for a crescent-shaped crista temporalis that extends caudomedially to near the midline (Fig. 5), as in Kelenken (not preserved in Devincenzia). Caudolateral to this ridge, the skull roof slopes ventrally and displays a distinct depression, the fossa temporalis, which corresponds to the origin for the m. adductor mandibulae externus profundus. The crista nuchalis sagittalis (that separates both fossae temporalis) is hourglass shaped. It is much wider between the fossae temporalis than in Devincenzia pozzi, Kelenken guillermoi, Andalgalornis steulleti, and the Psilopterinae and somewhat wider than that in the NHMUK A529 specimen of Phorusrhacos longissimus. Only a small portion of the caudal margin of the temporal fossa is visible to the left of the midline.
The septum interorbitale is quite robust and is broken at approximately midheight of the skull (Figs. 4, 6). It is lateromedially compressed, expanding caudally to the cranial border of the braincase. Its rostral margin is intact, forming a sharp ridge. The lateral wall of the septum is steep and shows a narrow, longitudinal sulcus olfactorii preserved on the right side of the skull. Caudally, a heart-shaped fossa cranii rostralis indicates the space occupied by the telencephalon, and more specifically by well-developed eminentia sagittalis, a feature present and developed in all phorusrhacids (Degrange et al., Reference Degrange, Tambussi, Ridgely and Witmer2016a, Reference Degrange, Tambussi, Ridgely and Witmerb). Small crenulations of the brain are visible on the roof of the braincase (Fig. 6) and may represent impressions of the vena cerebralis dorsorostralis and vena cerebralis dorsocaudalis. This region of the braincase is divided medially by a low, rounded crista frontalis interna, which widens caudally near its contact with the missing rostral ridge for the fossa cerebelli. Lateral to the fossa cranii rostralis, an accessory fossa, interpreted here as an accessory attachment site for the m. adductor mandibulae externus profundus, opens ventrolaterally and slightly caudally (Fig. 6). This feature is absent in Psilopterinae, Mesembriornithinae, and other Phorusrhacinae such as Kelenken and Devincenzia, but is present, although shallower, in Patagornis.
Although the mandible of MPEF-PV 11356 is incompletely preserved, it includes most of the rostrum mandibulae and pars symphysialis and both left and right rostral portions of rami mandibulae (pars intermedia) (Fig. 7). The right ramus preserves the rostral margin of the fenestra mandibulae. The mandibular symphysis is robust (Table 1), and crushing has shifted the rostral portion to the right in dorsal view. The rostral tip of the symphysis is broken but preserves several small openings for the foveae corpusculorum nervosorum that probably housed sensory corpuscles within the rhamphotheca (Baumel and Witmer, Reference Baumel, Witmer, Baumel, King, Breazile, Evans and Berge1993). The margo dorsalis of the symphysis are textured with scars similar to those observed in the rostrum. In dorsal view (Fig. 7), the mandibular symphysis is bordered by lateral ridges that caudally become more elevated with respect to the symphysis floor. The floor of the mandibular symphysis slopes caudoventrally and terminates in a V shape along its caudal margin, which is dissimilar to what it is observed in other Phorusrhacos longissimus specimens (e.g., NHMUK PV A529, A530, MLP 20-118, MPM-PV4241). The preserved rostral portion of the fenestra mandibulae exhibits a wide rounded edge, indicating the presence of an oval-shaped fenestra. The position of the rostral limit of the fenestra relative to the caudal limit of the symphysis exhibits variation among Phorusrhacos specimens. In NMHUK A529, the distance between the rostral margin of the fenestra and the caudal limit of the symphysis is substantial, whereas in MPM-PV4241 these features are positioned much closer to one another. MPEF-PV 11356 shares similarities with the latter specimen. An obvious feature of MPEF-PV 11356 is the extension of the ventral margin of the symphysis caudally, which accentuates the sigmoid shape of the jaw. This feature is quite similar to that of MNHUK A530 but is even more pronounced in MPEF-PV 11356. A rostrocaudally elongated furrow excavates the ventral and cranial margin of the fenestrae mandibulae (Fig. 7), penetrating them rostrally. This opening represents the rostral opening of the Meckelian canal.
Table 1. Measurements of MPEF-PV 11356 jaw symphysis and specimens referred to Phorusrhacos longissimus compared with other phorusrhacids. Parentheses indicate estimates based on incomplete elements. H = height; L = length; W = width.

Materials
Partial skull (MPEF-PV 11356; Fig. 4) including a large part of the rostrum, skull roof, and orbit, as well as the distal portion of the mandible. The roof of the skull is incomplete, with the occiput and large part of the fossae temporales missing. The mandibular symphysis is preserved as well as partial rami mandibulae; however, the mandibulae are broken caudal to the rostral margin of the external mandibular fenestra.
Remarks
Several features preclude MPEF-PV 11356 from assignment to other taxa for which skull material has been reported within Phorusrhacinae. In MPEF-PV 11356, the separation between the fossae temporales is less than that of Devincenzia but greater than that of Kelenken. The pila supranasalis has a bow shape when viewed laterally, a condition absent in Kelenken. The processus supraorbitalis of the os lacrimale is not completely fused with the orbital rim of the os frontale as in Kelenken. The processus postorbitalis is less developed (Fig. 4) than in either Devincenzia or Kelenken, and MPEF-PV 11356 presents an accessory fossa for the insertion of the m. adductor mandibulae extrenus profundus that is absent in Devincenzia and Kelenken. Supporting this assessment, the material proceeds from a Santacrucian locality from which only a single species of Phorusrhacos has been described, Phorusrhacos longissimus, and the size of MPEF-PV 11356 is consistent with the known size range of P. longissimus. Fragmentary cranial remains housed at AMNH and assigned to P. longissimus (AMNH 9158) are strikingly similar in size and morphology to the material presented here.
Phorusrhacinae indeterminate
Occurrence
Puesto Estancia La Costa locality, SE of Santa Cruz Province, Argentinean Patagonia, santacrucian SALMA (late early Miocene) of Santa Cruz Formation.
Description
The vertebra is strongly mediolaterally compressed and shows pronounced muscular scars on the cranial and caudal face of the processus spinosus as well as other morphological similarities with described thoracic vertebrae of the patagornithine Andalgalornis steulletis (Patterson and Kraglievich, Reference Patterson and Kraglievich1960; Tambussi et al., Reference Tambussi, de Mendoza, Degrange and Picasso2012) and those of the phorusrhacine Titanis walleri (see Gould and Quitmyer, Reference Gould and Quitmyer2005). It is narrower and taller than thoracic vertebrae of the Mesembriornithinae Mesembriornis milneedwardsi (Kraglievich, Reference Kraglievich1940), and the processus spinosus is relatively longer. Direct comparison with the nearly complete vertebral series from the patagornithine Andalgalornis steulleti suggests the vertebra of MPEF-PV 11355 may correspond to the second or third thoracic vertebra. The corpus vertebra is short (Fig. 8). In cranial view, the facies articularis cranialis of MPEF-PV 11355 is wide and saddle-shaped with a low ridge on its lateral margin. This facies is continuous at its base with a tall processus ventralis corporis that displays an asymmetrical, laterally expanded kink. The processus spinosus is very high and notably expanded dorsally. The foramen vertebrale is oval in shape. In lateral view, the processus ventralis corporis is rounded at its distal end; proximally, it spans nearly the length of the centrum. Placed on the ventrolateral surface of the stout processus transversi, the fovea costalis for the articulation with the tuberculum costae can be observed; meanwhile laterocaudally to the facies articularis cranialis, the fovea costalis for the articulation with the capitulum costae are present as a dorsoventrally oriented, oval shallow depression. Ventral to this fovea, a large pneumatic foramen penetrates the centrum. The opening for the foramen is of similar size and shape to the fovea costalis but is craniocaudally oriented. Dorsal to this large foramen, two smaller accessory pneumatic foramina penetrate the ventral surface of the processus transversi where it attaches to the centrum. The facies articularis caudalis is narrower and taller than the facies articularis cranialis.

Figure 8. Photograph and interpretive drawing of the thoracic vertebra of Phorusrhacinae MPEF-PV 11355 in caudal, right lateral, and cranial views. fac = facies articularis caudalis; facr = facies articualris cranialis; fc1 = fovea costalis for articulation with tuberculum; fc2 = fovea costalis for articulation with capitulum; fp = foramen pneumaticum; ps = processus spinosus; pt = processus transversus; pvc = processus ventralis corporis; zc = zygapophysis caudalis; zcr = zygapophysis cranialis. Scale bar = 1 cm.
Materials
A nearly complete thoracic vertebra (MPEF-PV 11355; Fig. 8).
Remarks
The identification of the vertebra MPEF-PV 11355 past the level of Phorusrhacinae is less certain, owing to a paucity of described or figured comparable material. It cannot definitively be referred to the species Phorusrhacos longissimus barring detailed postcranial diagnoses of species within phorusrhacine. Thus, it is here considered as Phorusrachinae indet. At 17.4 cm in height, it is smaller than the only undescribed thoracic vertebra for Titanis walleri but larger than complete vertebrae known from any other clades of phorusrhacids (e.g., Mesembriornithinae, Patagornithinae, and Psilopterinae).
Discussion
The cranial material (MPEF-PV 11356) is consistent with the mediolaterally compressed and dorsoventrally high terror bird skull type of Degrange (Reference Degrange2012), characterized by triangular ossa frontalia, processus supraorbitalis of the os lacrimale partially fused with the orbital rim, robust premaxillae, nares relatively small compared with the beak size, absence of the premaxillary fossa, and a dorsoventrally sigmoidal mandible. Although vertebra remains from other Phorusrhacinae have been reported, they have not been described in detail. Such is the case for a partially restored thoracic vertebra referred to Titanis walleri (see Gould and Quitmyer, Reference Gould and Quitmyer2005) and a fragmentary undescribed thoracic vertebra assigned to Devincenzia only on the basis of its size (Noriega and Agnolín, Reference Noriega and Agnolín2008). However, the lack of postcranial characters identified to diagnose distinct Phorusrhacinae species limits our referral of the recovered vertebra. We conclude its morphologies and size are consistent with referral to Phorusrhacos longissimus or a similarly sized phorusrhacine species.
The skull is referred to Phorusrhacinae because of its large skull size and the morphology of the mandible, specifically, the ratio of mandibular symphysis length to width at the base (~2.11), although a small amount of material missing from the tip of the mandible of MPEF-PV 11356 artificially decreases this ratio. These ratios are 2.38 in the holotype and 2.67 in the referred specimen NHMUK A529. Mandibular symphyses of bulkier phorusrhacids such as Physornithinae are shorter and wider at the base, with a ratio of 1.5 for Physornis fortis Ameghino, Reference Ameghino1895 (FM-P13340). However, it is worth noting that comparative mandibular material is not known for several other named species of Phorusrhacinae (i.e., Kelenken guillermoi, Titanis walleri, and Devincenzia pozzi). Smaller phorusrhacids such as Llallawavis scagliai Degrange et al., Reference Degrange, Tambussi, Taglioretti, Dondas and Scaglia2015 have slightly bigger ratios (2.74).
Within Phorusrhacinae, skull remains are generally limited, which complicates comparisons; those so far known are deformed (Kelenken), deformed and heavily artificially reconstructed (Devincenzia), or limited to fragmentary remains (Titanis). However, the nares of MPEF-PV 11356 appear to be less elongated than those present in Kelenken. The width of the skull roof between the fossae temporalis is greater in Phorusrhacos and Devincenzia than in Kelenken. In addition, in Phorusrhacos the processus postorbitalis is weakly projected ventrally when compared with the rest of Phorusrhacinae. Finally, the suturae frontolacrimalis and nasolacrimalis delimit an ‘L’ shape when viewed dorsally in the new specimen, while in Kelenken those suturae are not marked due to fusion of the bones (see the following). In this character, the condition observed in Phorusrhacos is more similar to that of the much smaller ‘Psilopterine’ skull type of Degrange (Reference Degrange2012) (e.g., Psilopterus and Llallawavis).
Several characters described in the preceding support the placement of MPEF-PV 11356 within Phorusrhacinae to the exclusion of other phorusrhacid groups and will likely benefit future analyses upon the recovery of more complete specimens. Although there is a comparative lack of cranial material among Phorusrhacinae specimens (Fig. 8), we propose the specimen MPEF-PV 11356 presented here is referable to Phorusrhacos longissimus.
The illustration of a skull of Phorusrhacos longissimus (Fig. 2, NHMUK A529; Ameghino, Reference Ameghino1895) differs substantially from that of MPEF-PV 11356 in the following ways: (1) the rostrum of MPEF-PV 11356 is dorsoventrally shorter and more elongated; (2) the external nares are elongated in MPEF-PV 11356 and are instead shown to be more quadrangular in the illustrated Phorusrhacos longissimus; (3) the ventral margin of the rostrum shows a strongly sigmoidal shape in the illustrated Phorusrhacos longissimus, while it is shallow and not as prominent in MPEF-PV 11356; and (4) the processus supraorbitalis of the os lacrimale is parallel to the plane of the skull roof in MPEF-PV 11356, whereas the caudal edge of this processus is ventrally deflected in the illustrated Phorusrhacos longissimus. It is important to consider here that the material depicted by Florentino Ameghino represents a field illustration of a skull that was highly fragmented. Commendable in its attempt, this illustration nevertheless was based on proportions of the jaw and beak hook, field measurements of length and width, in-person descriptions provided by his brother Carlos Ameghino, and references to the skull of Patagornis marshi (= Tolmodus inflatus; see specimen NHMUK A516). The problematic nature of this reconstruction is made clear by examination of the nares, as the nares illustrated for Phorusrhacos appear to be scaled-up versions of those seen in Patagornis marshi NHMUK A516 (see Ameghino, Reference Ameghino1895, fig. 1).
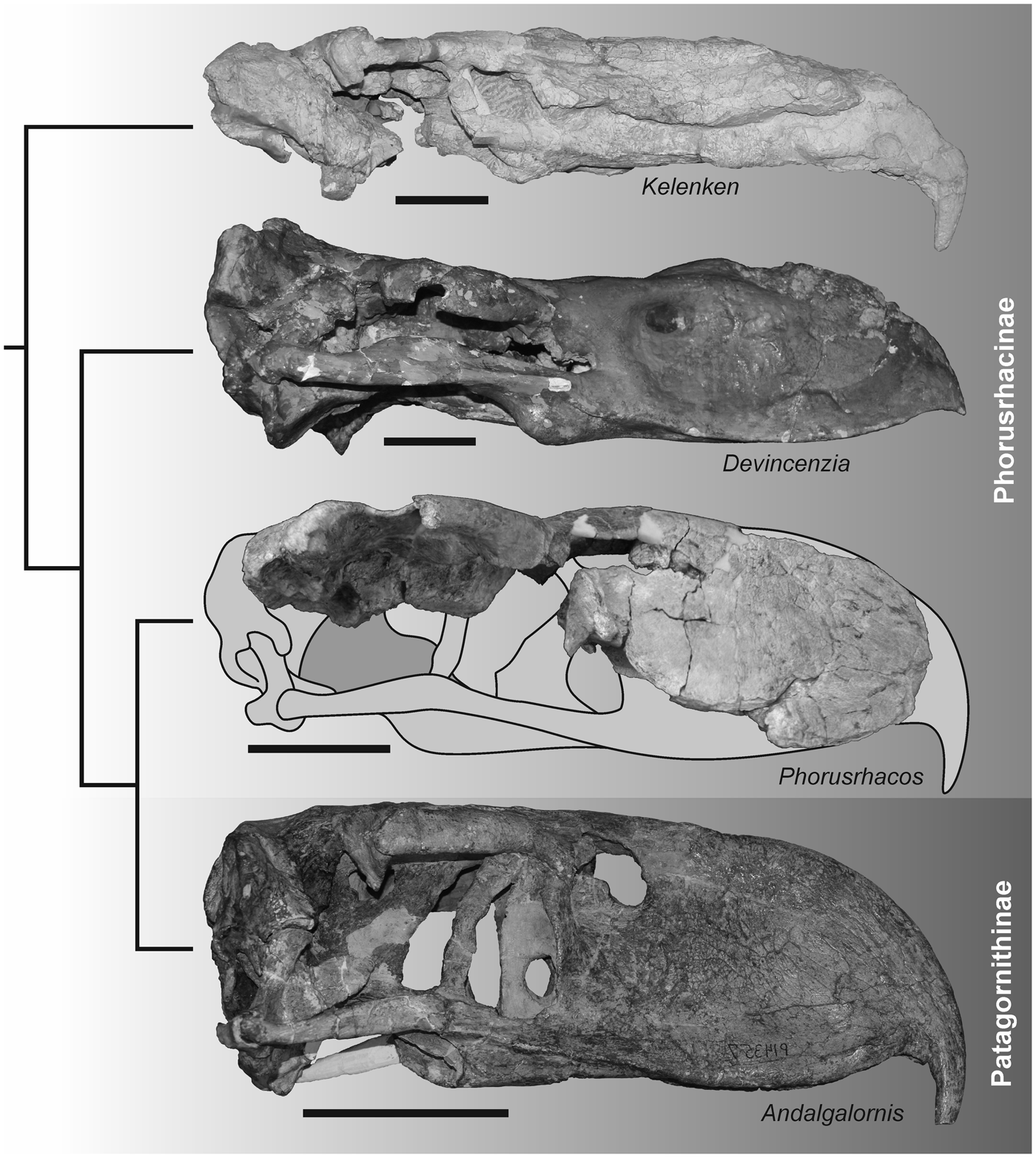
Figure 9. Lateral views of the skulls of the Phorusrhacinae Kelenken guillermoi BAR 3877-11, Devincenzia pozzi MLP 37-III-7-83, and Phorusrhacos longissimus MPEF-PV 11356 and the Patagornithinae Andalgalornis steulleti FM-P14357 displayed in a phylogenetic framework simplified from Degrange et al. (Reference Degrange, Tambussi, Taglioretti, Dondas and Scaglia2015). Skulls of Kelenken, Devincenzia, and Andalgalornis are mirrored for comparison. Scale bar = 10 cm.
The associated jaw presented here is also quite similar in morphology to that of P. longissimus. The only mandibles confidently referred to a taxon within Phorusrhacinae are from Phorusrhacos longissimus, although this element is well represented in other members of Phorusrhacidae, for example, Paraphysornis brasiliensis (Alvarenga, Reference Alvarenga1982), Andrewsornis abbotti Patterson Reference Patterson1941, Patagornis marshi, Andalgalornis steulleti, Llallawavis scagliai, and various psilopterines. Potential sexual dimorphism or ontogenetic variation notwithstanding, the mandible of MPEF-PV 11356 is similar in both size and shape to specimens of Phorusrhacos longissimus (including the holotype specimen) and specifically shares with that taxon a mandibular symphysis more than twice as long as its width at the symphysis base. This feature is the only character of the skull reported as diagnostic for Phorusrhacinae by Alvarenga and Höfling (Reference Alvarenga and Höfling2003) and distinguishes MPEF-PV 11356 from specimens referred to taxa within Physornithinae that possess a shorter, wider symphysis.
Conclusions
The new cranial material presented here is assignable to Phorusrhacos longissimus, one of the largest terror birds from Patagonia. The new vertebra, referred to Phorusrhacinae indet., calls attention to the paucity of described postcranial diagnoses for species in this clade. In light of the new cranial material, it is clear that some features previously described or mentioned that were based on the field drawings of Ameghino (Reference Ameghino1895) must now be revised. The new material allows us to state confidently that the skull of Phorusrhacos longissimus has the following previously unknown features: (1) narial openings are elongated; (2) the pila supranasalis forms a bow shape in lateral view; (3) the processus supraorbitalis of the os lacrimale is parallel to the orbital rim and caudally detached from the orbital margin; (4) the suturae frontolacrimalis and nasolacrimalis delimit an ‘L’ shape when viewed dorsally; (5) the processus postorbitalis is stout, although less developed than the rest of phorusrhacids; and (6) an accessory cotyla for the attachment of the m. adductor mandibulae externus profundus is present. The new specimens provide data critical to filling existing gaps in the knowledge of the evolutionary history of the most iconic fossil giant predatory birds. This description provides further key cranial comparisons among closely related phorusrhacine taxa and calls attention to the need for the identification of new cranial and postcranial morphological characters needed to better understand the phylogenetic relationships among phorusrhacines, and phorusrachids more generally.
Acknowledgments
We thank E. Ruigomez from the Museo Egidio Feruglio (Trelew) for granting access to the material. Our thanks to the following for allowing and facilitating the consultation of the collections under their management: W. Simpson, J. Holstein, and K. Lawson from the Field Museum of Natural History (Chicago); C. Norris, D. Brinkman, and M. Fox from Yale Peabody Museum (New Haven); C. Mehling from the American Museum of Natural History (New York); M. Reguero and A. Scarano from the Museo de La Plata (La Plata); and M. Taglioretti and F. Scaglia from the Museo Tradicional Lorenzo Scaglia (Mar del Plata). Phoebe Wang is thanked for her skillful drawings of the vertebra. Thanks to the editor D. Ksepka and three anonymous reviewers for their helpful comments. This research was supported by Jackson School of Geosciences to DE and JC.











 Open access
Open access