



Introduction
Biodiversity in oceanic ecosystems is poorly characterized and under threat from multiple human activities and fragmented global ocean governance (Jones et al., Reference Jones, Klein, Halpern, Venter, Grantham, Kuempel, Shumway, Friedlander, Possingham and Watson2018; Crespo et al., Reference Crespo, Dunn, Gianni, Gjerde, Wright and Halpin2019). Top predators, such as whales and dolphins (cetaceans) are no exception and may also act as valuable indicators of high oceanographic or biological diversity (Findlay et al., Reference Findlay, Best, Ross and Cockcroft1992; Azzellino et al., Reference Azzellino, Fossi, Gaspari, Lanfredi, Lauriano, Marsili, Panigada and Podestà2014; Di Tullio et al., Reference Di Tullio, Gandra, Zerbini and Secchi2016). The occurrence of cetaceans in the eastern South Atlantic Ocean has been poorly studied to date with most available information arising from 20th century whaling records and opportunistic records from the few islands in the area such as Gough and Tristan da Cunha (Best et al., Reference Best, Glass, Ryan and Dalebout2009; Elwen et al., Reference Elwen, Findlay, Kiszka and Weir2011). In the past 10 years, passive acoustic monitoring and satellite telemetry have begun to shed light on current patterns of seasonality and movement for some species, reflecting the rapid changes taking place in cetacean presence in the area as populations recover from whaling (Best et al., Reference Best, Glass, Ryan and Dalebout2009; Elwen et al., Reference Elwen, Findlay, Kiszka and Weir2011; Rosenbaum et al., Reference Rosenbaum, Maxwell, Kershaw and Mate2014; Shabangu et al., Reference Shabangu, Findlay, Yemane, Stafford, van den Berg, Blows and Andrew2019; Thomisch et al., Reference Thomisch, Boebel, Bachmann, Filun, Neumann, Spiesecke and Van Opzeeland2019).
The eastern South Atlantic Ocean is a geologically complex area which includes several islands, multiple seamounts and the Walvis Ridge, a 2000 km long underwater mountain range, all of which may act as aggregation sites or play a role in shaping whale migration routes (Best et al., Reference Best, Findlay, Sekiguchi, Peddemors, Rakotonirina, Rossouw and Gove1998; Rosenbaum et al., Reference Rosenbaum, Maxwell, Kershaw and Mate2014; Garrigue et al., Reference Garrigue, Clapham, Geyer, Kennedy and Zerbini2015). Seamounts are submarine mountains that rise to at least 1000 m above the seafloor but do not break the surface, providing shallow areas in the mid-ocean. They are often formed by volcanic action and have the typical steep conical shape of volcanic islands (White, Reference White2005; Bergstad et al., Reference Bergstad, Høines, Sarralde, Campanis, Gil, Ramil, Maletzky, Mostarda, Singh and António2019b). As essentially submerged islands, seamounts influence the surrounding waters through their effect on currents and the provision of shallow water habitat in otherwise oceanic areas. Seamounts in areas beyond national jurisdiction (offshore of the exclusive economic zone of coastal states) are not managed by any one country and may be heavily fished by multiple nations (Watling and Auster, Reference Watling and Auster2017). In such cases, regional sea conventions (the South East Atlantic Fisheries Organisation in this area) are responsible for managing potential human impacts such as fishing, although this is challenging in offshore areas where baseline data and monitoring are typically poor or absent. The Vema Seamount, at 31.6°S and 8.3°E, is in the eastern South Atlantic Ocean, 980 km west of Cape Town, approximately halfway between the continent and the southern end of the Walvis Ridge. The roughly conical seamount rises from the abyssal plain of more than 3000 m depth to within 20 m of the sea surface at its shallowest. The top is a plateau roughly 11 km × 8.5 km with an average depth of 90–100 m (Bergstad et al., Reference Bergstad, Høines, Sarralde, Campanis, Gil, Ramil, Maletzky, Mostarda, Singh and António2019b). This range of habitat (shallow vs deep) and topography (steep slopes, flatter summit) supports a high level of biological diversity in a small area, including many species native to the Indian Ocean. It hosts an intriguing mixture of shallow water taxa including kelp and corals on its upper areas as well as deeper-water taxa on its slopes such as crayfish, sponges and pelagic armour-head fish (Bergstad et al., Reference Bergstad, Gil, Høines, Sarralde, Maletzky, Mostarda, Singh, António, Ramil, Clerkin and Campanis2019a), which had been the target of significant fishing effort in the past. Vema Seamount is classified as a vulnerable marine ecosystem according to United Nations Food and Agriculture Organisation criteria and following heavy exploitation, has been closed to fisheries since 2007 (FAO, 2019).
The eastern South Atlantic Ocean marine ecosystem is dominated by the nutrient-rich waters of the Benguela upwelling ecosystem which runs for 2000 km along the south-western edge of the African continent. The cooler upwelling waters extend to roughly 300 km offshore or the shelf edge (Shannon and Nelson, Reference Shannon, Nelson and Wefer1996; Ansorge and Lutjeharms, Reference Ansorge, Lutjeharms and Best2007) while the broader limit of the current and its impacts can be extended to the eastern limit of the South Atlantic subtropical gyre (Veitch et al., Reference Veitch, Penven and Shillington2010), which overlaps the Vema Seamount. The Benguela upwelling is a highly productive ecosystem that provides an important mid-latitude feeding ground for at least two baleen whale species, the southern right (Eubalaena australis) and humpback whale (Megaptera novaeangliae) (Barendse et al., Reference Barendse, Best, Thornton, Pomilla, Carvalho and Rosenbaum2010; Mate et al., Reference Mate, Best, Lagerquist and Winsor2011) and is home to two endemic dolphins: the Heaviside's dolphin (Cephalorhynchus heavisidii) and the subspecies of African dusky dolphin (Lagenorhynchus obscurus obscurus) (Elwen and Gopal, Reference Elwen and Gopal2018; Alafaro-Shiguieto et al., Reference Alafaro-Shiguieto, Crespo, Elwen, Lundquist and Mangel2019). Additionally, large intrusions of warm water move into the South Atlantic Ocean from the Indian Ocean in the form of Agulhas ‘rings’ which break off from the Agulhas Current as it flows westwards around the southern tip of Africa (Ansorge and Lutjeharms, Reference Ansorge, Lutjeharms and Best2007). The combination of the strong westward flowing Agulhas Current and net transport of large amounts of tropical Indian Ocean water is thought to effectively form a one-way-barrier to the dispersion of marine organisms including cetaceans (the so-called South African species gate). These current patterns result in the tropical Atlantic Ocean being richer in cetacean species and other marine species than the adjacent Indian Ocean (Perrin, Reference Perrin and Best2007). For example, the tropical Atlantic Ocean hosts five species of the dolphin genus Stenella, while all other tropical oceans have only three (Perrin, Reference Perrin and Best2007).
A range of cetacean species are expected to occur in the region of the Vema Seamount or be seen en route from Cape Town. For most baleen whales – the timing of the survey from late October to early November overlaps with the southward migration from warmer water-breeding grounds to cooler water-feeding grounds. Spatially, the latitude of the seamount (31.6°S) lies along the migration path of most of the large whales including blue (Balaenoptera musculus), fin (Balaenoptera physalus), humpback and sei (Balaenoptera borealis) whales (Best, Reference Best2007). Vema Seamount also lies approximately at the southern end of the migration path of the West African ‘Offshore’ Bryde's whales (Balaenoptera brydei), which migrate up to equatorial West Africa during winter (Best, Reference Best2001), and near the northern distribution limit of pygmy right whales (Caperea marginata) (Best, Reference Best2007). Antarctic minke whales (Balaenoptera bonaerensis) are thought to occur in low numbers throughout the region (Best, Reference Best2007), with peaks in acoustic detections occurring off western South Africa in September–October (Shabangu et al., Reference Shabangu, Findlay, Yemane, Stafford, van den Berg, Blows and Andrew2019) and northern Namibia in June–August (Thomisch et al., Reference Thomisch, Boebel, Bachmann, Filun, Neumann, Spiesecke and Van Opzeeland2019) but with some presence year round.
Southern right whales, which winter along the southern African coast to the east of the survey area (Elwen and Best, Reference Elwen and Best2004), may migrate through this area to feeding grounds. Historically, catches by Yankee whalers operating during the 18th and 19th centuries were made in this part of the South Atlantic as reported on global-scale charts by Townsend (Reference Townsend1935), with these data now available through the OBIS SeaMap database (Halpin et al., Reference Halpin, Read, Fujioka, Best, Donnelly, Hazen, Kot, Urian, LaBrecque, Dimatteo, Cleary, Good, Crowder and Hyrenbach2011; Smith et al., Reference Smith, Reeves, Josephson and Lund2012). Although habitat modelling of these and other more recent data suggests the area remains suitable foraging habitat for right whales (Purdon et al., Reference Purdon, Shabangu, Yemane, Pienaar, Somers and Findlay2020) the spatial precision of the older whaling data is fairly broad and fine-scale association with the complex oceanographic and bathymetric features are not captured. More recent data since the 20th century suggest that southern right whales typically remain much farther south of the Vema Seamount; for example, southern right whales are reported occasionally around Tristan da Cunha (2000 km WSW of Vema Seamount) and acoustic detections of ‘gunshot’ sounds were made during October–November at a hydrophone placed approximately 700 km ESE of the Vema Seamount (Shabangu et al., Reference Shabangu, Andrew and Findlay2021). Several southern right whales satellite tagged by Mate et al. (Reference Mate, Best, Lagerquist and Winsor2011) did not pass Vema, but aggregated in an area roughly mid-way between Vema and Tristan da Cunha (37–40°S and 0–3°E, just south of the end of the Walvis Ridge), suggesting that Vema Seamount may lie north of the typical migration paths of right whales, although it is important to note that the species has undergone significant shifts observed in their feeding distribution in the last 20 years (van den Berg et al., Reference Van Den Berg, Vermeulen, Valenzuela, Bérubé, Ganswindt, Gröcke, Hall, Hulva, Neveceralova, Palsbøll and Carroll2020) which are likely not yet fully described.
Humpback whales seen near or en route to Vema Seamount are likely part of the population that breeds off West Africa (Rosenbaum et al., Reference Rosenbaum, Pomilla, Mendez, Leslie, Best, Findlay, Minton, Ersts, Collins, Engel, Bonatto, Kotze, Meÿer, Barendse, Thornton, Razafindrakoto, Ngouessono, Vely and Kiszka2009). The current consensus on humpback whale population structure (IWC, 2010) is that there are two separate but linked stocks of humpback whales off West Africa, a breeding and calving ground in the Gulf of Guinea (from Angola northwards) and a summer feeding ground in the Benguela ecosystem (where whales were seen aggregating in high numbers during the survey period, Ross-Marsh et al., Reference Ross-Marsh, Elwen, Fearey, Thompson, Maack and Gridley2022). Humpback whales satellite tracked from the Gulf of Guinea followed two migration paths, with some animals staying on the African continental shelf while others followed the Walvis Ridge southwards, essentially diverging either side of Vema Seamount.
A range of odontocete species may occur in the region of Vema Seamount or be seen en route but almost nothing is known about the distribution, numbers or behaviours of the smaller odontocete species in the high seas of the eastern South Atlantic Ocean other than which species are likely to occur. The combination of timing overlapping with seasonal migrations, and a location which straddles the range limits of several species, means that the cetacean diversity at Vema Seamount has the potential to be very high, but also difficult to predict. To add to the limited knowledge available for this area, here we present the results of a ship-based survey from Cape Town, South Africa to Vema Seamount during October and November of 2019, which included both visual survey and towed passive acoustic monitoring from the vessel, and passive acoustic monitoring from moored receivers at the seamount.
Methods
Data collection
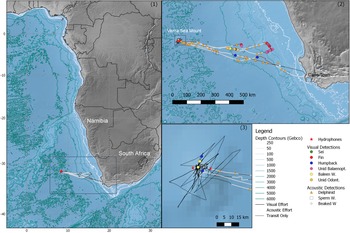
Visual observations of cetaceans were collected during a multi-objective, ship-based cruise from Cape Town, South Africa to Vema Seamount aboard the Greenpeace ship MY Arctic Sunrise, a 50 m converted seal hunting and supply vessel. The cruise departed Cape Town on 23rd October 2019, taking 2.5 days to transit to the seamount where it spent 2 days undertaking various tasks including deploying the two moored hydrophones. The ship then returned most of the way to Cape Town to undertake a crew-change between 27th and 30th October near the African shelf edge, before returning to the seamount for a further 4 days and finally departing on 4th November after retrieving the hydrophones, returning to Cape Town on 7th November (Figure 1).

Figure. 1. Map of the Vema Seamount and southern Africa at three different spatial scales, showing the survey line followed by the Arctic Sunrise, cetacean sightings and acoustic detections, and the location of the hydrophones at the seamount.
Dedicated visual surveys for cetaceans were conducted from the ship's bridge, approximately 7 m above sea level. Visual surveys were led by a experienced observer (author JF), who was occasionally assisted by a crew member. Depending on weather, observer/s were situated outside on the bridge wing or from within the bridge. Surveys started 15 min after sunrise and continued throughout the day with the target of a 2 h on, 30 min rest period duty cycle but with changes due to weather or logistical constraints dependent on the priorities of the overall expedition. Effort logs were kept comprising data on survey times, weather condition (cloud cover in eighths, Beaufort Sea state, wind strength and direction, and angle and intensity of sun glare), the number of observers, presence of other vessels and any opportunistic notes pertinent to the survey. Although visual search effort was focused forward, all cetacean encounters were logged, along with associated time, and distance (estimated visually), behaviour and group size (minimum, best and maximum estimates). Individuals or animals within approximately 5 body lengths were treated as a group. Where possible, animals were photographed to assist in species confirmation using a Canon 6D with a 100–400 mm L series lens and built-in GPS. On occasion, the ship adjusted course to facilitate confirmation of species identity. Encounters were tracked as the ship passed groups to avoid duplication, and ad hoc sightings made by crew or during non-survey periods were also logged.
With respect to species identification, small-to-medium-sized balaenopterid whales (such as sei, Bryde's and minke whales) are challenging to identify at sea from a distance without extensive experience of identifying the animals in the survey area, as there are regional variations in colour patterns and behaviour. Ideally, one would need a close approach for observation and photographs of distinguishing characteristics, but it was not possible to change the ship's course. As such, we have taken a precautionary approach identifying animals to the finest taxonomic category we could following De Rock et al. (Reference De Rock, Elwen, Roux, Leeney, James, Visser, Martin and Gridley2019). Notably, animals which were clearly a baleen whale (too large to be a beaked whale but with an upright blow and therefore not a sperm whale) were identified as ‘unidentified baleen whale (UBW)’. If a falcate dorsal fin was observed, thereby excluding humpback and right whales, then this was refined to ‘unidentified balaenopterid (UBal)’. Where images were available, photographic frame numbers were noted for each cetacean encounter, and species or category identification was confirmed retrospectively through inspection of the photographs.
Passive acoustic monitoring: towed-hydrophone array
Passive acoustic data were continuously collected from the Arctic Sunrise using a towed-hydrophone array during all available hours while the ship was transiting. The linear four-element array was deployed in a 10 m oil-filled polyurethane tube from a 350 m cable secured to the stern of the ship. The four hydrophone elements were grouped into pairs, two mid-frequency (MF) Benthos QA4 elements and Magrec HP02 preamplifiers (nominal frequency response 50 Hz to 40 kHz) and two high-frequency (HF) Magrec HP03 hydrophone and preamplifier units (nominal frequency response 2–200 kHz), each with a sensitivity of −201 dB re 1 V/μPa. Analogue signals were sent to a SAIL DAQ sound acquisition card with all four channels being digitally sampled at 16 bits and 500 kHz. The MF channels had a 6 dB gain and a 10 Hz high-pass filter, and the HF channels had a 12 dB gain and a 2000 Hz high-pass filter. This system was used to continuously record acoustic data using PAMGuard (Gillespie, Reference Gillespie, Gordon, Uk, West, Tay and Redmond2008) through a dedicated computer onboard the Arctic Sunrise for offline post processing.
Acoustic analysis of towed-hydrophone data
Acoustic wav files were processed in PAMGuard offline using a click detection threshold of 16 dB. Various thresholds were tested in 3 dB steps from 10 to 19 dB to ensure maximum noise removal while keeping clicks from marine mammals. The vessel's propulsion system (propeller and engine noise) was still the source of many false detections, and so a 20° angle veto was implemented to minimize the amount of noise from the vessel triggering the click detector. The click train detector module (Macaulay, Reference Macaulay2020) within PAMGuard was used to group clicks into trains. Settings for the detector were provided by Macaulay (Reference Macaulay2020). Three spectral template classifiers for sperm whales, delphinids and beaked whales were also added using the defaults within PAMGuard. In order to assess the suitability of the provided settings, a subset of the entire data set (3.1%) were manually annotated for odontocete detections. This subset contained 52.3 h of acoustic data from a larger 1696.3 h data set which includes the data collected between Cape Town and the Vema Seamount reported here. A comparison between times of click train detections and the manual audit of the 3.1% subset gave a precision rate of 0.69, and a recall rate of 0.99, with only one manually identified sperm whale train missed, and no manually identified delphinid trains being missed. These settings were then applied across the entire data set.
PAMGuard's ‘Whistle and Moan Detector’ (Gillespie et al., Reference Gillespie, Caillat, Gordon and White2013) was used to detect odontocete whistles up to 24 kHz on data decimated to 48 kHz from the ‘medium frequency’ hydrophone pair, using the settings provided in Gillespie et al. (Reference Gillespie, Caillat, Gordon and White2013). The Whistle and Moan Detector identifies tonal sounds within acoustic data using a multi-stage process that removes noise, applies an amplitude threshold and joins detected tonal sounds that are close in time and frequency.
Manual verification of recordings was then conducted on sections identified by the detectors as containing potential odontocete presence. Click trains and whistles were manually marked up in PAMGuard and spectrograms were checked externally in Raven 1.6 (K. Lisa Yang Center for Conservation Bioacoustics, 2023). Each audio file was scanned at the specific time indicated by the PAMGuard data. The original, undecimated files were used (FFT: 27,900, contrast = 55, brightness = 55, colour = jet black, Hann window default). Files were viewed from 0 to 22 kHz for files with only whistles, or 0 to 42 kHz for files with echolocation clicks. Delphinid encounters were defined as periods of whistles and/or echolocation clicks separated by at least 20 min of silence, while individual sperm whale and beaked whale click trains were treated as separate events. If any overlap between clicks and whistles occurred, they were included in the same event, except if the clicks originated from sperm whales.
Passive acoustic monitoring: static monitoring at Vema Seamount
Two autonomous hydrophones (Ocean Instruments SoundTrap 300HF) were deployed at two sites 12 km apart located on the eastern (31.6371°S, 8.40481°E, water depth: 70 m, 25th October 2019) and western (31.6338°S, 8.27811°E, water depth: 55 m, 27th October 2019) slopes of the seamount with the intended goal of maximizing the area monitored acoustically. To minimize the risk of instrument loss and keep the moorings upright in the currents, moorings were fitted with both acoustic release mechanisms and surface buoys. Hydrophones were mounted approximately mid-water column at each site to minimize background noise from reefs and the sea surface. Instruments were deployed mid-afternoon, with recordings scheduled to start in the evening (18:00 UTC + 2, which was the time zone of the departure location at Cape Town), allowing time for the ship to move away. Instruments were recovered at 11:30 (east) and 15:39 (west) on 4th November 2019. Acoustic data were recorded continuously, with a 96,000 Hz sample rate (flat frequency response 20 Hz–48 kHz ± 3 dB) and file length was limited to 5 min to facilitate data handling.
Acoustic analysis of static hydrophones
Long-term spectral averages (LTSAs) summarize large acoustic data series and are especially useful to identify the broad characteristics of the acoustic data sets (including aspects of the soundscape such as snapping shrimp, fish chorus, boat noise, recording interference). Vocal signals which are clearly defined and prominent, such as humpback whale song, balaenopterid call series, odontocete echolocation, diurnal fish choruses and boat echo sounders are easily differentiated from the background soundscape. In such cases, prominent energy peaks within signature frequency bands (i.e. group or species-specific) can be observed quickly and then further investigated to confirm sound sources (see e.g. Ross-Marsh et al., Reference Ross-Marsh, Elwen, Prinsloo, James and Gridley2020). LTSAs were generated from the acoustic data collected at each monitoring site using the MATLAB package Triton (Wiggins et al., Reference Wiggins, Roch and Hildebrand2010) with the settings: 5 s time resolution and 50 Hz frequency. The final 45 min of recording time were removed prior to analysis as ship noise dominated the recordings.
In the second stage of analysis, a fine-scale visual inspection was undertaken. The entire data set was carefully inspected by an experienced analyst (author TG) in Raven Pro 1.6 to investigate overall sound types and patterns of production. For logging the presence of sounds, we used a point sampling approach in which the first 1 min from each 5 min of recording was systematically logged. Recordings were scanned at two different frequency resolutions: 0–1000 Hz for low-frequency baleen whale calls (data set decimated to 2000 Hz sample rate, spectrogram settings: FFT 512, Hann window), and 0–48,000 Hz for mid-frequency cetacean calls, including odontocete echolocation clicks and whistles (original sample rate, spectrogram settings: FFT 512, Hann window). The entire recordings were again manually inspected at the lower frequency for the presence of humpback whale song and non-song calls and analysed by Ross-Marsh et al. (Reference Ross-Marsh, Elwen, Fearey, Thompson, Maack and Gridley2022), so we are confident that all whale call types present were detected. For this paper, diel and weekly temporal patterns over the deployment were described as the sum of detection positive minutes (dpm) per day, which were subsampled at 1 min every 5 min (100% presence would be represented by 288 dpm per day).
As boat noise can mask detections, characteristic boat noise was also logged during this process. The characteristics of each sound type were defined where possible, by measuring the time and frequency parameters (low frequency, high frequency, frequency range, peak frequency, peak power and duration) of clear and prominent call examples (identified through visual inspection – those measured had a strong signal-to-noise ratio (SNR) and were not masked, as per Gridley et al., Reference Gridley, Cockcroft and Janik2012). Spectral parameters were measured from the spectrogram display in Raven Pro 1.6 software. As calls were simultaneously detected at both sites, data measurements were conducted from one deployment site only (the site with the best examples) to avoid pseudo-replication. Results on temporal occurrence of calls are presented graphically, together with spectrographic examples of the main calls detected which can aid comparisons with other geographic areas and species.
Results
Visual survey effort
Visual surveys took place over 14 days between 23rd October 2019 and 7th November 2019 for a total of 52 h. In total, there were 39 cetacean encounters made during visual surveys by the observer and/or crew (Figure 1, Table S1). Most of the visual survey effort occurred during the ship transit to and from the seamount, with observation time at the seamount limited due to conflicting cruise objectives. Consequently, the majority of sightings (n = 31 or 79%) were made during transit days, with a high proportion (n = 21 or 53% of total sightings) seen on a single day (29th October 2019) while passing over the African continental shelf edge. UBal was the most frequently seen taxonomic group (n = 18 encounters), followed by seven groups of humpback whales, six of fin whales, four unidentified odontocetes, four UBW and one sei whale. Encounters classified as ‘unidentified odontocetes’ were all sightings of one or two individuals with very small blows. The five cetacean sightings at Vema Seamount were all baleen whales. Of the seven encounters with humpback whales, four encounters were approximately midway from Cape Town to the seamount and three were at the seamount. An encounter on 4th November consisted of two individuals on the southwestern side of the seamount, diving repeatedly in the same location for approximately an hour and a half. The observed behaviour; long dives in the same location (est. 10 min dive duration), interspersed with periods of surface activity (e.g. 5–7 min of surface activity including pectoral slapping), indicated possible feeding behaviour.
Passive acoustic monitoring: towed-hydrophone array from the vessel
The towed-hydrophone array was deployed for a total of 78.7 h during transit legs to and from the seamount, recording during both day and night. In total 82 detections of odontocete cetaceans were made using the PAMGuard Whistle and Moan Detector. Ten were removed following manual verification as having a likely non-biological origin (electrical noise) and the 26 sperm whale clicks were grouped into five encounters (three single detections and two clear clusters, Figure 1). A single-beaked whale was identified acoustically using the PAMGuard-beaked whale detector and based on the characteristics of its clicks identified as likely Mesoplodon densirostris or Ziphius cavirostris. The remaining 45 encounters were all identified as likely delphinids (two by clicks only, 28 by whistles only and 15 by both clicks and whistles). Spatially the non-sperm whale odontocete detections were spread widely across the survey tracks with no apparent preference for any depth or habitat strata covered. None of the acoustic detections could be matched with visual observations for species confirmation, nor are enough confirmed calls from known species available for this area for confident species classification based on only acoustic characteristics.
Static acoustic monitoring: soundscape and cetacean detections at the seamount
Continuous passive acoustic monitoring for cetaceans was conducted from two mooring locations on the east and west sides of Vema Seamount for 10 (233 h) and 8 (189 h) days, respectively. Initial investigation of the LTSAs indicated that the ambient soundscape at the Vema Seamount was dominated by constant biophony (sounds from animals) in the form of generalized reef noise including snapping shrimp (frequency range: 5–20 kHz, with a peak energy at 10 kHz, Figure S1). Clear diel patterns in reef noise were apparent, which was more prominent from 18:00 to 06:00 than from 06:00 to 18:00 (Figure S1, apparent as banding within the LTSAs). However, no obvious odontocete echolocation or whale song were identified from the LTSA scans.
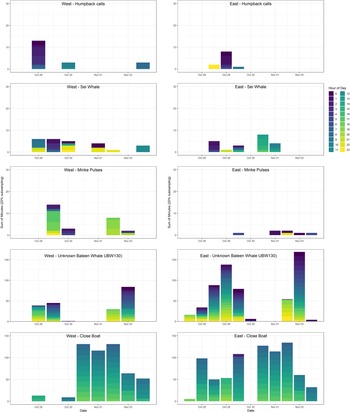
A detailed sub-sampling approach was employed to interrogate the data further. There was an absence of odontocete vocalizations in the subsampled data set. However, calls from several baleen whale species were positively identified (humpback whales, sei whales and minke whales) as well as calls which could not confidently be assigned to species, described below. The variation in acoustic detections between days (Figure 2) indicates a diversity of cetacean species around Vema Seamount, none of which had a consistent diel pattern in calling behaviour (Figure S2).

Figure. 2. Daily occurrence of the dominant whale calls and close boats detected over the duration of the deployment of two SoundTrap hydrophones on the western (left panels) and eastern (right panels) sides of Vema Seamount during October and November 2019. Data presented as sum of detection positive minutes (dpm) per day, which were subsampled at 1 min every 5 min (100% presence represented at 288 dpm per day).
Humpback whale
The presence of humpback whales was confirmed through the occurrence of non-song calls on 5 days over the study period, usually only at one site, except for 28th October when calls were detected on both hydrophones. Positive detections were slightly higher at night than during daylight hours on the west, while at the eastern site, detections were mostly overnight (Figure S2), suggesting that only a few animals were vocalizing in the area for a short period. The detected calls included those identified from other Southern Hemisphere humpback whale populations (Dunlop et al., Reference Dunlop, Noad, Cato and Stokes2007; Rekdahl et al., Reference Rekdahl, Tisch, Cerchio and Rosenbaum2017; Ross-Marsh et al., Reference Ross-Marsh, Elwen, Fearey, Thompson, Maack and Gridley2022), including ‘whups’ and ‘grumbles’. Faint detection of likely humpback whale song (~30 s) was possible on one day, but as the SNR was low and song structure could not be characterized, this was not analysed further. The non-song data are presented in greater detail in Ross-Marsh et al. (Reference Ross-Marsh, Elwen, Fearey, Thompson, Maack and Gridley2022).
Sei whale
Frequency-modulated down-sweep calls, characteristic of sei whales, were detected periodically at both sites but with no predictable diel pattern (Figures 3a and S2). Calls were identified as sei whales based on their frequency, duration (Table 1) and regular repetition (Figure 3a), matching those described from the North Atlantic Ocean (Nieukirk et al., Reference Nieukirk, Mellinger, Dziak, Matsumoto and Klinck2020; Romagosa et al., Reference Romagosa, Baumgartner, Cascão, Lammers, Marques, Santos and Silva2020). They occurred in doublet and triplet bouts (visually identified through shorter inter-call intervals within than between calls) with up to five calls per bout, which are also characteristic of the species. Calls were detected with a greater SNR at the western site.
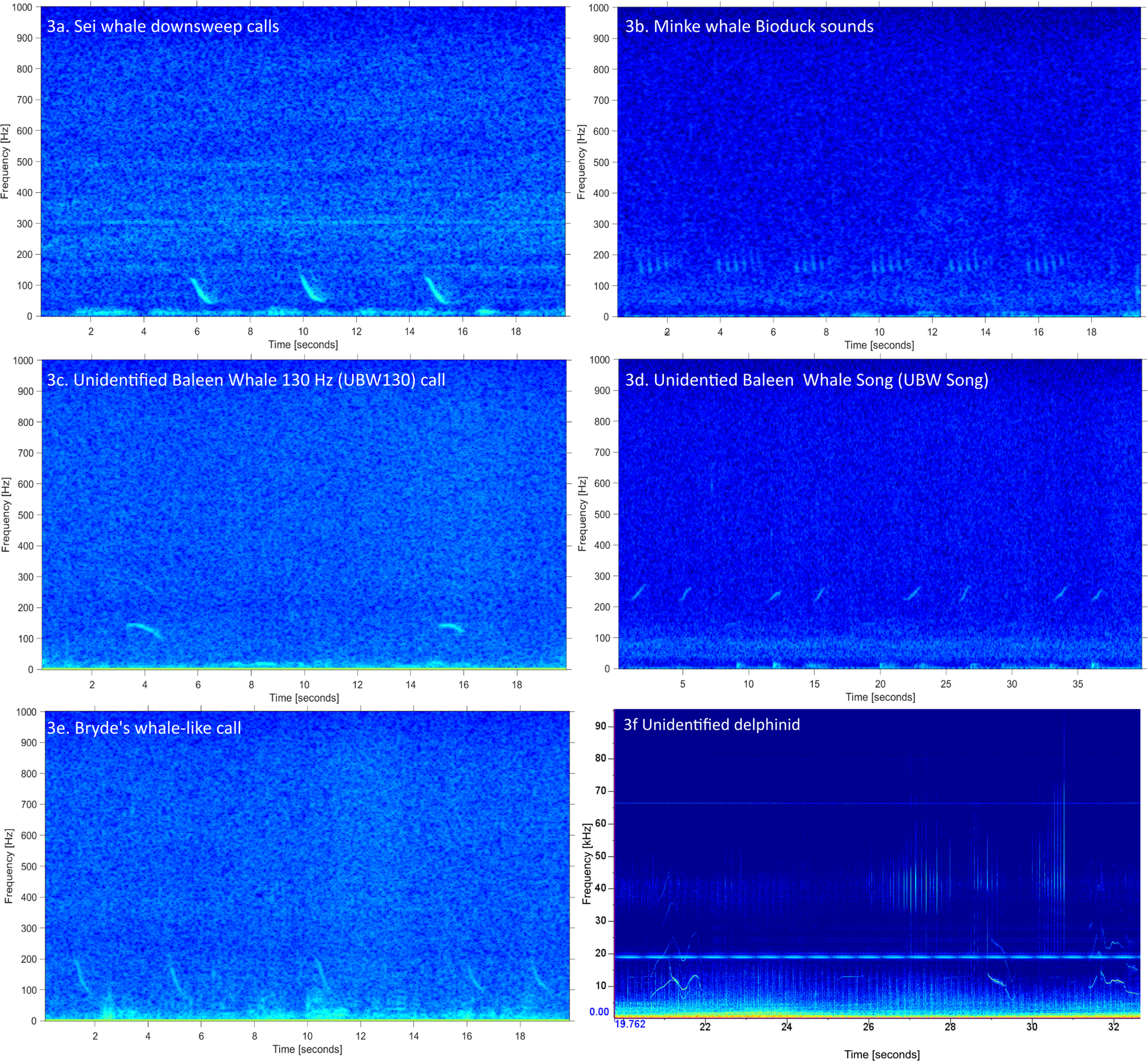
Figure. 3. Spectrograms of whale calls identified from moored acoustic recorders at Vema Seamount during October–November 2019. Frequency parameters described in Table 1 and timing of detections in Figures 2 and S2. Note that Figures 3a–3e are recorded from the hydrophones moored on Vema Seamount, the dolphin recording in Figure 3f is displayed on difference axes and is from the towed-hydrophone array recorded en route.
Table 1. Summary statistics of baleen whale tonal calls identified from both hydrophones moored at Vema Seamount during October and November 2019. SNR is a subjective measure of signal-to-noise ratio for the calls in each section, following Gridley et al. (Reference Gridley, Cockcroft and Janik2012) (1 - faint, 2 - clear, 3 - prominent)

Minke whale
Minke whale ‘bio-duck’ calls are characterized by repeated bouts of 4–8 pulses produced in packets (Figure 3b) (Risch et al., Reference Risch, Gales, Gedamke, Kindermann, Nowacek, Read, Siebert, Van Opzeeland, Van Parijs and Friedlaender2014). Bio-duck calls were confirmed more often on the western hydrophone (mostly in the evening hours) than at the eastern site (mostly in the early morning hours; Figures 2 and S2).
UBW130 call
The most common call detected at Vema Seamount was a simple frequency-modulated call with peak power around 130 Hz (Table 1, Figure 3c). We refer to this call as ‘UBW130’ in reference to its dominant frequency and the assertion that it was produced by an unidentified baleen whale (UBW) based on the frequency and temporal production properties of the calls. It was often detected with a relatively strong SNR. The observed call structure was either down-swept, constant or modulated with a ‘u’ shape. Calls in bouts were produced with a regular repetition and amplitude, indicating the presence of one caller throughout recordings. No overlapping calls were observed, and although possible, there was no clear evidence for call exchange between two or more individuals. Inter-call intervals averaged between 20 and 30 s, which differs from the bout patterns common in sei whale call production. Call-by-call matching confirmed simultaneous detection of the same calls at the two sites on several occasions. This call type was detected on most days on both recorders but more commonly at the eastern recorder. There were two peaks in detection on 27–29 October and 2–3 November, discernible at both locations. There was no clear diel pattern in calling. Despite an extensive literature search and consulting external expert opinion, we cannot confidently match these calls to any specific whale species at this point.
UBWSong
A repeated call-type arranged into a single ‘song’ was detected only once over 11 min on 3rd November 2019 (Figure 3d, Table 1). The constituent call unit was simple in structure and the song did not share the characteristic hierarchical pattern familiar in humpback whale song (units, phrases, themes; Payne and McVay, Reference Payne and McVay1971). The SNR was fair and the sound production characteristics, a regularly produced upsweep, together with the frequency, indicate production by a baleen whale. Song units at the start were slightly higher in frequency than other baleen whale calls identified (units of 287–444 Hz, range: 157 Hz) but dropped over the duration of the detection in minimum and maximum frequency and frequency range (units of 207–281 Hz, range: 74 Hz). The received level was initially low and increased and then decreased during detection, indicative of a biological and possibly moving sound source. Despite an extensive literature search and consultation with external experts, this song could not be attributed to any one species and requires further investigation.
Bryde's-like
No studies of Bryde's whales call repertoires are available from southern Africa but we identified a call similar to published Bryde's whale calls (Wang et al. Reference Wang, Duan, Chen, Mei, Sun, Nong, Liu, Akamatsu, Wang and Wang2022) from other regions and thus use the conservative term Bryde's-like. The call was down-swept and significantly higher in frequency than the sei calls identified. Bryde's-like calls ranged from 87 to 158 Hz (Figure 3e, Table 1) and were detected as a few bouts between 22:40 and 23:15 on 27th October 2019 on the eastern hydrophone. Antarctic minke whales also produce downsweep calls but those are typically lower in frequency and shorter in duration than the calls described here, nor did they overlap in time with detected minke whale bioduck calls.
Fish and other biological sounds
We detected pulse repetitions and cracks which were likely produced by the Vema Seamount fish community. In general, such sounds are not well characterized, particularly in the eastern South Atlantic Ocean and in the interest of brevity we have not discussed these further but keep the focus of this paper on cetaceans.
Anthropogenic
Distant ship noise was ubiquitous throughout recordings (low frequency, peak energy < 1 kHz), with ship noise from closer vessels detected at both mooring sites. Transient ship noise (peak energy < 1 kHz but with broadband energy when passing close to the hydrophone) was detected on most days (Figures 2 and S2). This was mostly generated by the Arctic Sunrise itself, although close-boat noise attributed to likely fishing vessels or container vessels which were seen operating in the area was also detected on the days when the Arctic Sunrise was absent (28–30 October 2019).
Discussion
In this study we use visual sightings and passive acoustic monitoring to describe the presence and acoustic behaviour of cetaceans in a poorly studied region of the eastern South Atlantic Ocean, including Vema Seamount. Most toothed whale and dolphin species in the South Atlantic Ocean are thought to be wide-ranging residents or have relatively small seasonal shifts in distribution while the majority of baleen whales are migratory (Best, Reference Best2007). Three species of baleen whale, humpback, fin and sei, were confirmed visually during the expedition, although only humpback whales were visually confirmed at the seamount. Most sightings could not be confirmed to the species level due to the distance, the brevity of observation while the ship was underway and the challenge of identifying balaenopterid whales to the species level at sea without a close approach. Only four visual sightings of groups of potential odontocetes occurred, none of which were at Vema Seamount. All were very small groups with short low blows, and unfortunately none could be confidently assigned to the species level due to distance from the ship and poor weather conditions. Note that no visual survey effort took place over the continental shelf where there are known high densities of dusky and Heaviside's dolphins, as that part of the transit was undertaken at night. Odontocetes were only acoustically detected during the transit including both sperm whales (n = 5) and echolocating and whistling delphinids (n = 45), and one beaked whale detection, all of which were fairly evenly spread along the track line with no clear spatial clustering. Without visual confirmation of acoustically recorded delphinid species, or the availability of acoustic classifiers trained on regionally confirmed species (e.g. Erbs et al., Reference Erbs, Elwen and Gridley2017 for coastal delphinids around southern Africa), we are hesitant to assign finer taxonomic classification to the acoustically detected delphinids at this stage and rather leave these questions to future analyses and concentrate our discussion on baleen whales from here on.
Acoustic recordings from the static hydrophones moored on the seamount confirmed the detection of humpback, sei and minke whales. Additionally, detection of a short period of downsweep calls from potentially Bryde's whale, and two other call types, could not be confidently confirmed to the species level despite extensive literature searches and external consultation. Despite many years of cetacean research in the southern African subregion (Elwen et al., Reference Elwen, Findlay, Kiszka and Weir2011) and an increasing use of passive acoustic monitoring in the South Atlantic Ocean (Shabangu et al., Reference Shabangu, Yemane, Stafford, Ensor and Findlay2017; Hawkey et al., Reference Hawkey, Elwen, James, Prinsloo and Gridley2020; Ross-Marsh et al., Reference Ross-Marsh, Elwen, Prinsloo, James and Gridley2020; Shabangu et al., Reference Shabangu, Andrew, Yemane and Findlay2020a), the calls of several baleen whale species remain poorly described or unconfirmed (e.g. Thomisch et al., Reference Thomisch, Boebel, Bachmann, Filun, Neumann, Spiesecke and Van Opzeeland2019). Very few studies visually confirmed the species of whale recorded (McDonald et al., Reference McDonald, Hildebrand, Wiggins, Thiele, Glasgow and Moore2005; Risch et al., Reference Risch, Gales, Gedamke, Kindermann, Nowacek, Read, Siebert, Van Opzeeland, Van Parijs and Friedlaender2014; Wang et al. Reference Wang, Duan, Chen, Mei, Sun, Nong, Liu, Akamatsu, Wang and Wang2022), reflecting the challenging nature of studying these wide-ranging animals that have calls which can be detected tens to hundreds of kilometres away. Species which likely occur in the study area but with poorly known vocalizations include pygmy right whales (with only a single paper published globally; Dawbin and Cato, Reference Dawbin and Cato1992), Bryde's and dwarf minke whales. However, even among those species that have fairly well described calls, such as blue, sei and humpback whales, there can be geographic and temporal variation in production making confirmation to species level challenging (Darling and Sousa-Lima, Reference Darling and Sousa-Lima2005; McDonald et al., Reference McDonald, Hildebrand and Mesnick2009).
The detection of Antarctic minke whale calls during October and November in this study supports the findings of Shabangu et al. (Reference Shabangu, Findlay and Stafford2020b), who detected regular vocalizations at their hydrophone moored roughly 700 km ESE and closer to Cape Town, and by Thomisch et al. (Reference Thomisch, Boebel, Bachmann, Filun, Neumann, Spiesecke and Van Opzeeland2019), who detected calls off the Walvis Ridge to the north of Vema Seamount during July and August. These recent acoustic studies support older data primarily from the 20th-century whaling era, that Antarctic minke whales have a more protracted migration pattern than the larger baleen whales with some animals remaining in tropical and temperate waters year-round (Best, Reference Best2007).
Humpback whales are likely the most abundant baleen whale in the South Atlantic Ocean (Wedekin et al., Reference Wedekin, Engel, Andriolo, Prado, Zerbini, Marcondes and Kinas2017). Given the overlap in timing of the cruise with the southern migration of West African humpback whales, and indications from other studies that seamounts may act as navigation beacons for migrating humpback whales (Garrigue et al., Reference Garrigue, Clapham, Geyer, Kennedy and Zerbini2015), the low number of humpback whale detections both visually and acoustically was unexpected. The low numbers and inconsistent acoustic detections, combined with the shift in detections from the western to eastern hydrophone from 28th October to 4th November 2019, suggest that only a few vocalizing humpback whales were present, which is consistent with the low number of visual sightings. No clear humpback whale song was detected, despite extensive recording of song at this time of year off the coast of South Africa where animals are migrating south and lingering to feed in the Benguela ecosystem (Gridley et al., Reference Gridley, Silva, Wilkinson, Seakamela and Elwen2018; Ross-Marsh et al., Reference Ross-Marsh, Elwen, Prinsloo, James and Gridley2020). All usable acoustic detections at Vema Seamount were of non-song vocalizations which typically occur during mating, socializing and feeding (Dunlop et al., Reference Dunlop, Noad, Cato and Stokes2007; Rekdahl et al., Reference Rekdahl, Dunlop, Noad and Goldizen2013, Reference Rekdahl, Dunlop, Goldizen, Garland, Biassoni, Miller and Noad2015). Humpback whales feeding in the super-groups off western South Africa are very vocal and multiple call types have been detected (Silva, Reference Silva2017; author's unpublished data), although to date none are linked specifically to feeding. The majority of acoustic detections of humpback whale calls were at night. This nocturnal peak in vocalizations is common in both calling and singing humpback whales (Kowarski et al., Reference Kowarski, Evers, Moors-Murphy, Martin and Denes2018; Ross-Marsh et al., Reference Ross-Marsh, Elwen, Prinsloo, James and Gridley2020, Reference Ross-Marsh, Elwen, Fearey, Thompson, Maack and Gridley2022) and is not yet fully understood, but has implications for the detection of humpback whales during short deployments or using duty-cycled recordings. The visual observation of potential feeding and detection of non-song calls does indicate the use of the seamount as a potential stop over for at least some migrating humpback whales. During the study period in 2019, high numbers of humpback whales were observed along the South African west coast closer to shore, with many animals engaged in active feeding, with some groups observed well into early 2020 (author's unpublished data).
The low number of humpback whale detections either visually or acoustically during the survey at Vema Seamount does support the observation of widely separated migration routes for humpback whales at this latitude, though we would need survey effort across multiple years to confirm this. Satellite tagging of humpback whales off equatorial Gabon (Rosenbaum et al., Reference Rosenbaum, Maxwell, Kershaw and Mate2014) revealed that humpback whales migrating southwards from this breeding ground follow two different migration routes: (1) along the coast/continental shelf edge (and through the Benguela feeding ground), or (2) following the Walvis Ridge, taking whales west of the Vema Seamount. There is some genetic differentiation between humpback whales from western South Africa and Gabon, but with some known interchange of biopsied and photographically identified individuals between these sub-stocks (Rosenbaum et al., Reference Rosenbaum, Pomilla, Mendez, Leslie, Best, Findlay, Minton, Ersts, Collins, Engel, Bonatto, Kotze, Meÿer, Barendse, Thornton, Razafindrakoto, Ngouessono, Vely and Kiszka2009; Barendse et al., Reference Barendse, Best, Thornton, Elwen, Rosenbaum, Carvalho, Pomilla, Collins, Meÿer and Leeney2011). It is therefore possible that these whales use different migration routes to the feeding grounds, and that song is exchanged while on the high-latitude feeding grounds (Best and Allison, Reference Best and Allison2010; Barendse et al., Reference Barendse, Best, Thornton, Elwen, Rosenbaum, Carvalho, Pomilla, Collins, Meÿer and Leeney2011; Schall et al. Reference Schall, Djokic, Ross-Marsh, Oña, Denkinger, Ernesto Baumgarten, Rodrigues Padovese, Rossi-Santos, Carvalho Gonçalves, Sousa-Lima, Hucke-Gaete, Elwen, Buchan, Gridley and Van Opzeeland2022, Ramos et al., Reference Ramos, Cheeseman, Cesar, Olio, Vogel, Elwen, Facchola, Cipolotti, Southerland, Findlay, Seyboth, Mccue, Kotze and Seakamela2023).
Multiple sightings of balaenopterid whales were made during the cruise. Six were confirmed as fin whales and one as a probable sei whale, most of which took place near the edge of the continental shelf during the crew-change transit. The area (32.15°S, 15.60°E) lies just to the northwest of the location where the bulk of commercial sei whale catches were made by vessels operating from the Saldanha Bay whaling station (33–34°S, 16–17°E; Best and Lockyer, Reference Best and Lockyer2002). The bulk of those sei whale catches were reported to be inshore of the main migration stream (Best and Lockyer, Reference Best and Lockyer2002). Catches off western South Africa were only made between November and April and the bulk of whales were thought to be present between 25°S and 28°S (Best, Reference Best2001). Almost everything we know about balaenopterid whales in this region is based on whaling catches in the 20th century, and at that time Bryde's whales and sei whales were not recognized as separate species so catch histories for these two species are not well separated. It is possible that some of the reported sei whale catches (and unidentified whales seen on this cruise) were actually Bryde's whales from the offshore population (Best, Reference Best2001; Best and Lockyer, Reference Best and Lockyer2002). Fin whales were one of the species hardest hit by commercial whaling as they were targeted both in the Southern Ocean, where as many as 28,761 were reportedly taken in a single season during 1960/1961, as well as from shore-based stations around southern Africa (Findlay et al., Reference Findlay, Elwen, Meÿer, Oosthuizen and Plön2016). The timing of whale catches and recent passive acoustic monitoring efforts (Shabangu et al., Reference Shabangu, Findlay, Yemane, Stafford, van den Berg, Blows and Andrew2019; Thomisch et al., Reference Thomisch, Boebel, Bachmann, Filun, Neumann, Spiesecke and Van Opzeeland2019) show a single peak in fin whale numbers near the Walvis Ridge off northern Namibia, during June–August, and similarly off the south west coast of South Africa with a single peak in June–July but with some detections as late as November at both sites. Feeding fin whales were seen over several summer months (April–May) in 2 years in southern Namibia (author's unpublished data) during 2014/2015, and sporadic sightings are made in the southern Benguela. Combined, these observations suggest that the African coast is near the northern limit of the migration route of this species (hence a single peak in numbers) and that the Benguela ecosystem may be an important foraging area for this species, as it is for both humpback and southern right whales.
In summary, the cetacean observations and detections made during this expedition provide a valuable insight into an area for which little recent data are available. Remarkably little is known about the cetaceans of the eastern South Atlantic Ocean beyond the continental shelf of Africa since the end of commercial whaling (e.g. Findlay et al., Reference Findlay, Best, Ross and Cockcroft1992 ; Best, Reference Best2007; De Rock et al., Reference De Rock, Elwen, Roux, Leeney, James, Visser, Martin and Gridley2019; Purdon et al., Reference Purdon, Shabangu, Yemane, Pienaar, Somers and Findlay2020). As such, there is scarce information on distribution, abundance or even bycatch rates from which to base assumptions or comparisons. We report valuable information on the presence and timing of several baleen whales in the eastern South Atlantic Ocean including humpback, fin, sei and minke whales, and the role of Vema Seamount and the continental shelf edge of southern Africa as a feeding area for baleen whales. Vema Seamount is the only shallow water habitat for a minimum of 1000 km in any direction and has an abundance of life and prey availability. Although not reported in detail here, the results of passive acoustic monitoring at the seamount highlight the utility of this method for detecting vessel traffic and potentially illegal fishing effort in remote locations such as these. Such baseline data can feed forward into management of the high-sea regions of the eastern South Atlantic Ocean and emphasizes that further investment in research and a global framework for legally binding protection is essential. The discovery of unidentified baleen whale call types is exciting and highlights how much there is to learn about biodiversity in offshore areas of the ocean.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0025315423000255
Acknowledgements
We thank the crew of the Greenpeace vessel MY Arctic Sunrise for practical help during the research. We also thank the many logistics staff from Greenpeace International Ships Unit and Greenpeace Research Laboratories. In particular, we thank Grant Oakes, David Santillo, Paul Johnston and Sandra Schoettner for helping to make this research possible. We also thank Jonathan Gordon for equipment and Douglas Gillespie for his advice and guidance on PAMGuard analysis.
Financial support
This research was funded by the Umweltstiftung Greenpeace Foundation, Hamburg, and supported by the Greenpeace Research Laboratories, Biosciences, University of Exeter, UK.




 Open access
Open access