



Introduction
Marine biodiversity in temperate seas faces several threats, including climate change and invasive species (Halpern et al., Reference Halpern, Walbridge, Selkoe, Kappel, Micheli, D'Agrosa, Bruno, Casey, Ebert, Fox, Fujita, Heinemann, Lenihan, Madin, Perry, Selig, Spalding, Steneck and Watson2008). The increasing global temperatures associated with climate change have led to notable shifts in the distribution ranges of marine species, often pushing them towards higher latitudes (Gervais et al., Reference Gervais, Champion and Pecl2021). Additionally, anthropogenic changes in marine ecosystems can introduce invasive species, resulting in changes in species composition and thresholds in ecosystems such as intertidal rocky shores, temperate and tropical reefs, estuaries, and pelagic systems (Bulleri et al., Reference Bulleri, Batten, Connell, Benedetti-Cecchi, Gibbons, Nugues, Gribben, Hawkins, Allcock, Bates, Firth, Smith, Swearer, Evans, Todd, Russell and McQuaid2020). Such transformations in marine ecosystems can result in complex adverse interactions (Council of Europe, 2008).
Unlike terrestrial organisms, marine species tend to exhibit more pronounced geographic expansion in response to climate change. Planktonic larvae, in particular, can readily spread to new environments because of their transportability within turbulent boundary layers (Eckman, Reference Eckman1990; Pinsky et al., Reference Pinsky, Selden and Kitchel2020). Pinsky et al. (Reference Pinsky, Selden and Kitchel2020) reported two major trends in the geographical distribution of warm-water marine organisms in the Northern Hemisphere: (1) a northward expansion in their range and (2) a shift into deeper waters. Numerous warm-water organisms have extended their geographical habitats from lower to higher latitudes (Rubenstein et al., Reference Rubenstein, Weiskopf, Bertrand, Carter, Comte, Eaton, Johnson, Lenoir, Lynch, Miller, Morelli, Rodriguez, Terando and Thompson2023). For example, the Korean Peninsula has witnessed the northward movement of species such as perforated barnacles (Perforatus perforatus) and Korean top shells (Turbo sazae) due to the recent increase in seawater temperature along the East Sea (Kim et al., Reference Kim, Chan, Lee and Kim2020; Son et al., Reference Son, Lee, Park, Kim, Riedel, Hwang, Kim and Jung2020).
The precise prediction of ecological responses, including alterations in habitat range due to climate change, has become a pressing challenge (Bennett et al., Reference Bennett, Duarte, Marbà and Wernberg2019). Geographical information system (GIS) technology combined with species distribution modelling has shown that the rate of invasive species entering and spreading into new habitats has been accelerating globally (Maravelias and Reid, Reference Maravelias and Reid1997; Tittensor et al., Reference Tittensor, Baco, Brewin, Clark, Consalvey, Hall-Spencer, Rowden, Schlacher, Stocks and Rogers2009; Capinha et al., Reference Capinha, Anastacio and Tenedorio2012; Tyberghein et al., Reference Tyberghein, Verbruggen, Pauly, Troupin, Mineur and De Clerck2012; Assis et al., Reference Assis, Tyberghein, Bosch, Verbruggen, Serrão and De Clerck2018; Park et al., Reference Park, Lee, Kim and Shin2020). The 5th Assessment Report of the Intergovernmental Panel on Climate Change (IPCC, Reference Pachauri and Meyer2014) provided long-term climate change projections, termed representative concentration pathways (RCPs), defined by different carbon dioxide emission assumptions. In IPCC RCP4.5 scenario, technological developments enable substantial stabilisation of atmospheric carbon dioxide concentrations and a consequent deceleration of seawater warming. Conversely, the IPCC's RCP8.5 depicts unmitigated atmospheric carbon dioxide concentration increases and sharp temperature rises. Multiple studies have explored marine species distribution based on these IPCC RCP scenarios (e.g. Jones and Cheung, Reference Jones and Cheung2015; Allyn et al., Reference Allyn, Alexander, Franklin, Massiot-Granier, Pershing, Scott and Mills2020; Fabri-Ruiz et al., Reference Fabri-Ruiz, Danis, Navarro, Koubbi, Laffont and Saucède2020).
The red-throated ascidian Herdmania momus is an invasive, solitary sea squirt species (Class: Ascidiacea) that is globally distributed in tropical and subtropical marine habitats (Palomares and Pauly, Reference Palomares and Pauly2023). Recent GIS-based species distribution analysis has predicted range shifts for warm-water species along the Korean Peninsula (Park et al., Reference Park, Lee, Kim and Shin2020). The results showed that H. momus, which was first observed along the coast of Jeju Island in 1969 (Rho, Reference Rho1971), has spread throughout Jeju and reached parts of Busan. According to Park et al. (Reference Park, Lee, Kim and Shin2020) and the RCP8.5 scenario, the habitat range of H. momus is projected to expand from Busan to Ulsan by 2100. Due to its short lifespan (between 10 and 12 months) and rapid growth, H. momus can readily extend its habitat range in response to environmental changes (Yi and Kim, Reference Yi and Kim2016; Park et al., Reference Park, Lee, Kim and Shin2020). However, it may face spatial competition with perennial seaweeds (e.g. Sargassum spp. and Ecklonia cava) and/or corals (e.g. scleractinian corals and Alveopora japonica) in its introduced habitat.
The aim of this study was to examine the extent of northward expansion of the H. momus habitat in line with Park et al.'s (Reference Park, Lee, Kim and Shin2020) predictions based on IPCC RCP scenarios. H. momus habitat was assessed via qualitative scuba diving surveys conducted at nine stations in Busan, Ulsan, and Gyeongju between 2016 and 2022.
Materials and methods
Study area
The study area is located along the coastal region from Busan to Gyeongju, Korea (Figure 1), including the northernmost points where H. momus habitat has been confirmed or predicted previously by Park et al. (Reference Park, Lee, Kim and Shin2020). Observations were made at nine rocky subtidal stations within the geographically interconnected Busan–Ulsan–Gyeongju area, including three stations in Busan (BS-1, BS-2, and BS-3), four stations in Ulsan (US-1, US-2, US-3, and US-4), and two stations in Gyeongju (GJ-1 and GJ-2) (Figure 1). Three stations, BS-1, BS-3, and GJ-2, were among the National Marine Ecosystem Monitoring Program (rocky ecosystem) Project (NMMP) study sites. The remaining six stations were selected arbitrarily while considering the requirement for maintaining a consistent distance among study stations for this research (Table 1). The water depths of each study site were between 5.0 and 39.0 m, with US-2 and US-3 being shallower compared to the other stations, while GJ-1 and GJ-2 were deeper (Table 1).
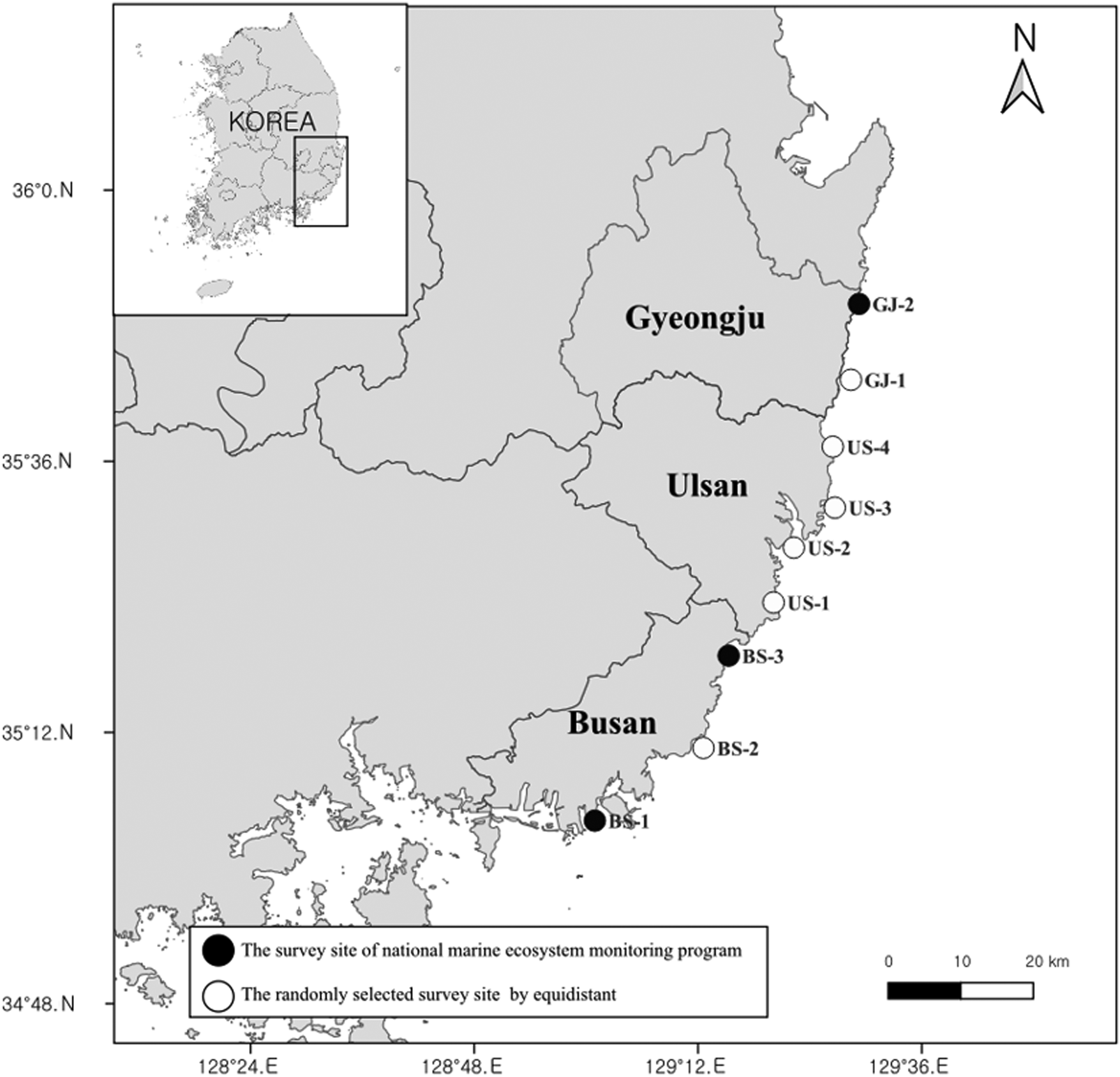
Figure 1. Locations of study sites (nine stations) for observing the presence of solitary ascidians via scuba observations from 2016 to 2022. Black symbols indicate three NMMP stations and open symbols are additionally selected stations.
Table 1. Characteristics of the study sites

Field survey
To document the distribution of H. momus and co-occurring ascidians, a field survey was conducted by underwater scuba diving at the nine study stations in rocky subtidal zones to water depths of 1‒20 m. In accordance with the NMMP framework, quantitative samples were collected according to the Protocol of National Survey on Marine Ecosystems (MOF and KOEM, 2022). Photographs were taken to identify solitary ascidians during the survey. As the present study focused on the latitudinal trends of solitary ascidians, the presence or absence of H. momus was identified by analysing high-resolution underwater images in a laboratory. At all stations, two professional divers moved approximately 20 m in a zigzag pattern while remaining at a distance of 2–5 m from each other. They photographed a number of the individual images encountered during their movement, which lasted approximately 30 min overall.
Field surveys were conducted between 2016 and 2022, except in 2019 when the sites were not surveyed. The three NMMP stations were examined twice a year (May and August), whereas observations at the other six stations were conducted once a year (between May and June).
We collected surface water temperature data from Marine Environment Monitoring stations operated by the Korea Marine Environment Management Corporation to understand the factors affecting the spatial distributions of ascidians (https://www.meis.go.kr/portal/; accessed 10 February 2024).
Species identification and historical distribution
Solitary ascidians photographed from field surveys were identified to the species level based on various taxonomic references (e.g. Bae et al., Reference Bae, Lee, Kim and Yi2022; Shenkar et al., Reference Shenkar, Gittenberger, Lambert, Rius, Moreira da Rocha, Swalla and Turon2023). Species identifications for each ascidian species were conducted by two or more researchers and then double-checked from expert ascidian taxonomists in Korea. The occurrence of each ascidian species as well as their geographical location and time were also recorded. The species identified by image analysis were initially annotated according to their Korean names based on the National List of Marine Species II: Marine Invertebrates (MABIK, 2022; MOF and KOEM, 2022), and their scientific names were confirmed in accordance with the World Register of Marine Species (WoRMS, 2023).
For H. momus, the current observations recorded on the southeastern coast of Korea were compared with previous global records from the biological databases of the Global Biodiversity Information Facility (GBIF, 2023).
Results
Sea surface water temperatures (SSTs) gradually decreased from southern to northern stations, but SSTs tended to increase over time across the entire study area (Figure 2). In May, SSTs remained below 16°C at all stations except for the southernmost station BS-1, but there was a notable increase in SSTs at middle latitude station BS-3 beginning in 2018, followed by a widespread increase in SSTs at most of the stations between 2021 and 2022. While in August, SSTs remained consistently above 23°C at three stations located in Busan coast (BS-1, BS-2, and BS-3); significant increases in SSTs were observed at all stations in Ulsan and Gyeongju during 2017, 2018, and 2021.
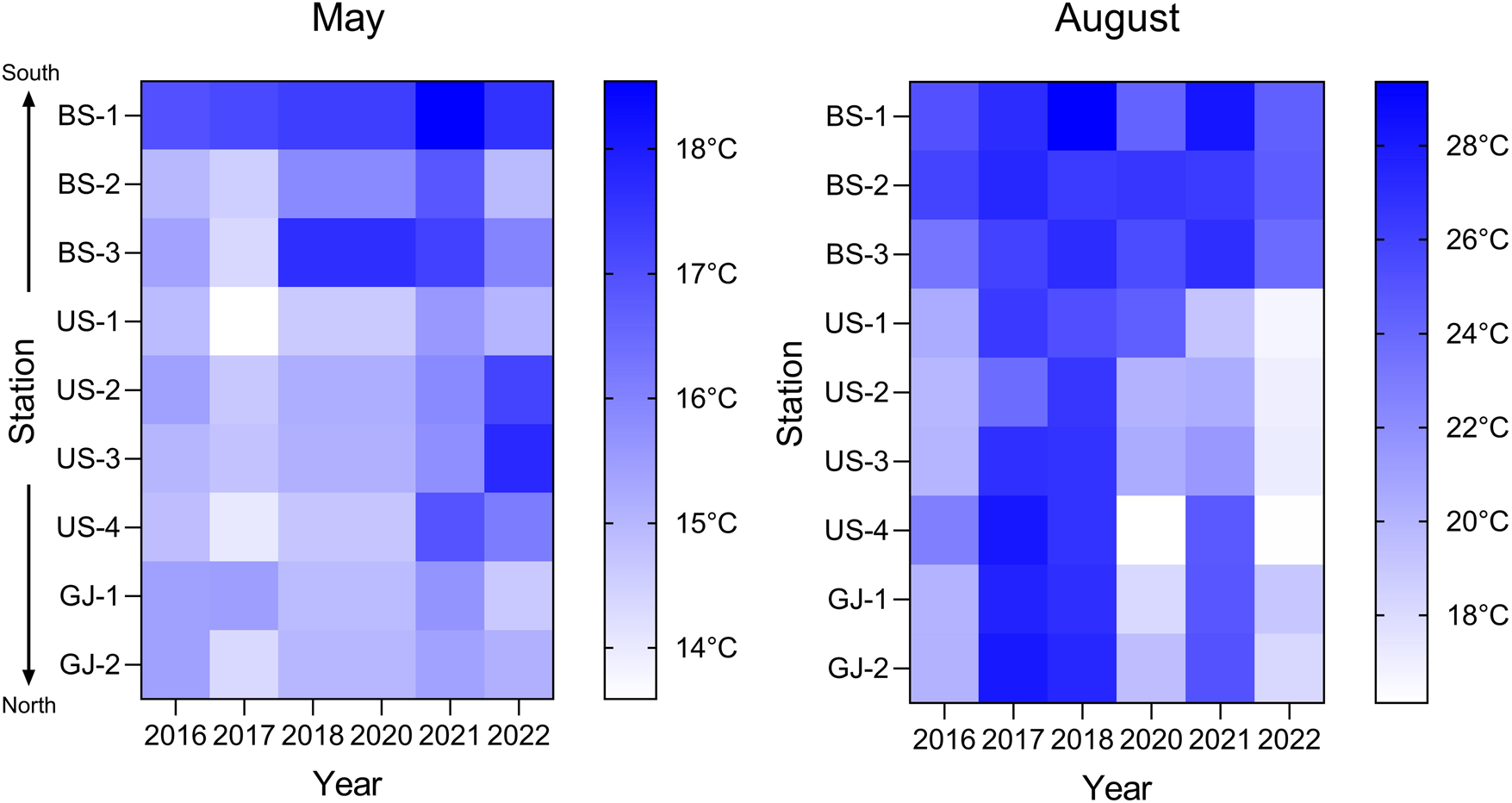
Figure 2. Heat map showing SSTs at nine study stations over 6 years between 2016 and 2022 on the southeastern coast of the Korean Peninsula.
Seven solitary-type ascidian species were identified across all nine stations, with Styela clava being the most frequently observed and Pyura sacciformis, the least common. The stations with the highest diversity of solitary ascidians were BS-3 and US-4, where six species were observed. The remaining stations showed the presence of between four and five ascidian species. In 2020, five of the seven species, with the exception of Boltenia echinata and P. sacciformis, were consistently observed at the southernmost station (i.e. BS-1) since 2016. The most common S. clava occurred at Busan and Ulsan stations during the study period, but not at the Gyeongju station.
The presence of H. momus was confirmed at five stations (BS-1, BS-3, US-3, US-4, and GJ-2). Notably, H. momus was observed only at the southernmost station, BS-1, from 2016 to 2020. However, their presence was observed for the first time at more northern stations, US-3 and US-4, in 2021, and their habitats expanded to GJ-2, the northernmost station within the study area by the last year of the survey (Table 2).
Table 2. Summary of Ascidiacea observation data from 2016 to 2022 by station
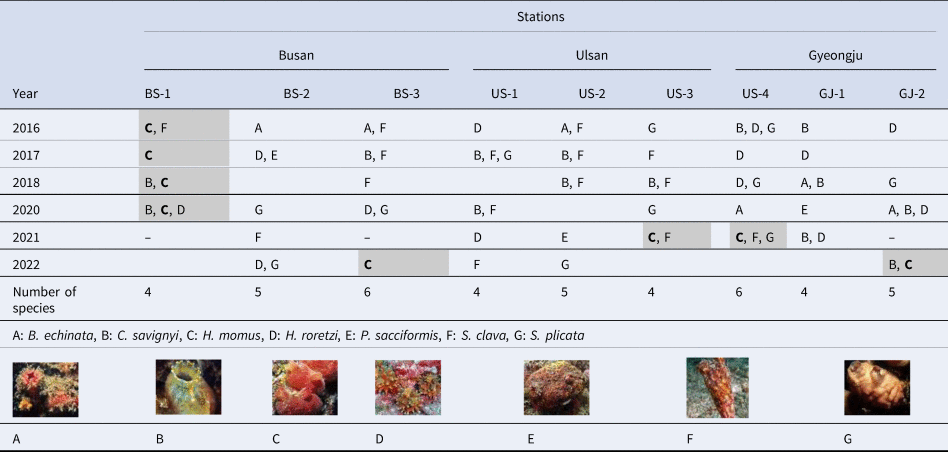
Grey shadings indicate the presence of H. momus.
Note: The labels A‒G correspond to arbitrarily assigned alphabetical designations used by the authors for species classification purposes. A blank space indicates that there no observed occurrences, while ‘–’ indicates that no examination of the site.
Discussion
In the present study, seven solitary-type ascidian species were observed in nine study locations from 2016 and 2022. Most of the ascidians were subtropical or tropical species except for B. echinata and Halocynthia roretzi (Palomares and Pauly, Reference Palomares and Pauly2023). Their lifespans were largely different from 1 to 4 years depending on the species, showing shorter lifespans in tropical species including Ciona savignyi, H. momus, and Styela plicata (Svane and Lundälv, Reference Svane and Lundälv1982; Nomaguchi et al., Reference Nomaguchi, Nishijima, Minowa, Hashimoto, Haraguchi, Amemiya and Fujisawa1997; Lambert and Lambert, Reference Lambert and Lambert1998; de Barros et al., Reference de Barros, da Rocha and Pie2009; Yi and Kim, Reference Yi and Kim2016). Generally, short-lived species take advantage of new opportunities to settle and colonise quickly in new marine environments by maximising population growth through high fecundity, rapid development, and a large investment in reproduction (Heppell et al., Reference Heppell, Heppell, Read, Crowder, Norse and Crowder2005). Among ascidians recorded in this study, tropical species, including H. momus, have relatively shorter longevity, and thus, they may have an advantageous edge for inhabiting newly settled temperate reef habitats (Figley, Reference Figley2003).
The main finding of the present study was the poleward extension of H. momus' habitat from the southernmost (35°03′41.5″N) to the northernmost (35°48′43.1″N) stations within our study area between 2016 and 2022. This finding indicates that by the end of the study period, the entire Busan–Ulsan–Gyeongju region had become part of the habitat range of H. momus. The rising seawater temperatures caused by global climate change have had a direct effect on the recruitment and habitat expansion of invasive species along the Korean Peninsula (Raitsos et al., Reference Raitsos, Beaugrand, Georgopoulos, Zenetos, Pancucci-Papadopoulou, Theocharis and Papathanassiou2010; Son et al., Reference Son, Lee, Park, Kim, Riedel, Hwang, Kim and Jung2020; Hong et al., Reference Hong, Jang, Kim, Kim, Park and Lee2022). These invasive species are expanding into other habitats through various routes, and their recruitment is progressing faster than predicted (Lee et al., Reference Lee, Reusser, Olden, Smith, Graham, Burkett, Dukes, Piorkowski and McPhedran2008; Capinha et al., Reference Capinha, Anastacio and Tenedorio2012).
H. momus is a typical tropical and subtropical species found primarily in the Indo-West Pacific Ocean, Atlantic Ocean, and Mediterranean Sea (Palomares and Pauly, Reference Palomares and Pauly2023). According to the GBIF database (Figure 3), the northernmost recorded distribution of H. momus in the West Pacific was at 35°03′30″N in 2015, along the Busan coast of Korea (GBIF, 2023).
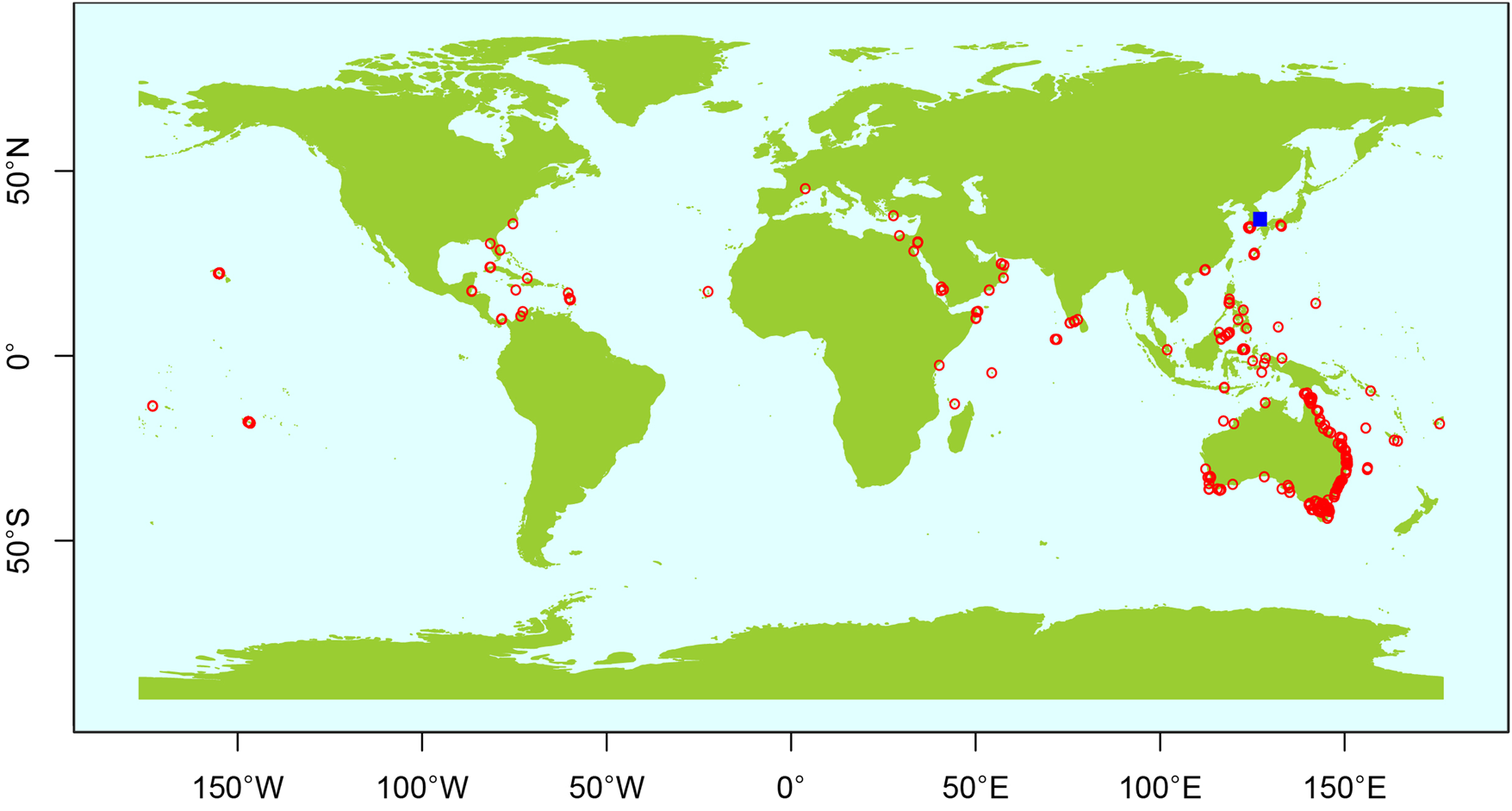
Figure 3. Distributional records of H. momus, showing historical records (red circles) from the Global Biodiversity Information Facility (GBIF, 2023) and the new record (blue square) off southeastern coast, South Korea.
Except for this record, the northern distribution limit of the species is approximately 35°N or lower latitudes in the Northern Hemisphere, whereas the southern distribution limit reaches approximately 43°S (GBIF, 2023). Therefore, our study presents a new northernmost record of H. momus, implying ca. 50′ poleward expansion in species distribution since the mid-2010s.
The use of GIS analysis coupled with species distribution modelling has significantly improved our ability to observe and predict the climate change-associated expansion of various taxa (Guinotte et al., Reference Guinotte, Bartley, Iqbal, Fautin and Buddemeier2006; Januario et al., Reference Januario, Estay, Labra and Lima2015; Assis et al., Reference Assis, Tyberghein, Bosch, Verbruggen, Serrão and De Clerck2018). Our present findings build upon previous observations of the northward expansion of the H. momus habitat range (Park et al., Reference Park, Lee, Kim and Shin2020). According to the IPCC (Reference Pachauri and Meyer2014) RCP4.5 scenario, which involves some mitigation of carbon dioxide concentrations, the habitat range of H. momus is projected to expand from the already established coastal habitats on Jeju Island to certain areas of the southern Korean Sea by 2100. In contrast, the worst-case RCP8.5 scenario predicts that the distributional expansion of H. momus will encompass the eastern part of the southeastern coast of the Korean Peninsula, including our study area. Park et al. (Reference Park, Lee, Kim and Shin2020) deduced that the most influential environmental factor affecting H. momus expansion was the maximum seawater temperature, which accounted for 77.1% of the expansion observed.
Since 2016, the habitat range of H. momus observed by Park et al. (Reference Park, Lee, Kim and Shin2020) has primarily included Jeju Island and some areas on the Busan coast. The observations we present in this study extend far beyond the habitat range of the RCP4.5 scenario, which predicted the habitat range of H. momus to remain relatively stable throughout 2100 (Park et al., Reference Park, Lee, Kim and Shin2020). Moreover, our observations of H. momus in the waters off Ulsan in 2021 indicate that the species has spread more rapidly than was predicted in the RCP8.5 scenario of unmitigated climate change, wherein it was anticipated to expand from Busan to Ulsan by 2100 (Park et al., Reference Park, Lee, Kim and Shin2020). This accelerated northward expansion of H. momus occurrences, beyond previous predictions, is likely due to the recent sharp increase in water temperatures in the East Sea (Lee and Park, Reference Lee and Park2019). Compared with global trends, sea surface temperatures around the Korean Peninsula, as well as the eastern coast of North America, are rising relatively quickly (Fasullo and Gent, Reference Fasullo and Gent2017; Widlansky et al., Reference Widlansky, Long and Schloesser2020). Therefore, continuous monitoring is required to better predict the habitat expansion of warm-water species and assess the impact of subtropical marine species on temperate marine ecosystems (Batten et al., Reference Batten, Hyrenbach, Sydeman, Morgan, Henry, Yen and Welch2006).
In conclusion, our study's findings suggest that the poleward habitat expansion of subtropical marine species entering the Korean Peninsula due to climate change outpaces previous predictions, at least in the case of H. momus. Although the IPCC's RCP scenarios are broadly used to predict future changes in marine ecosystems, it is difficult to apply them to specific regions experiencing rapid localised temperature increases, such as the Korean Peninsula. These scenarios are based on only four representative pathways and consider global socioeconomic, political, and policy factors, which limits their accuracy in predicting local marine ecosystem changes. Effectively addressing marine ecosystem changes in the Korean Peninsula will require the development of governmental policies that incorporate proactive measures and strategies to mitigate eventualities that exceed even the most recent predictions.
Data availability statement
Data availability is not applicable to this article as no new data were created or analysed in this study.
Acknowledgements
The authors thank all the individuals who contributed to the progress of this research.
Author's contribution
Choul-Hee Hwang: conceptualisation, data curation, methodology, formal analysis, and writing of the original draft. Su Jin Lee, Hyeong Ju Seok, and Hyun-Jung Kim: methodology and formal analysis. InSeo Hwang: conceptualisation, data curation, and supervision. Min Gu Kang: methodology, visualisation. Joo Myun Park: methodology, software, visualisation, writing – review, and editing.
Financial support
This research was part of the ‘National Marine Ecosystem Monitoring Program’ project, funded by the Ministry of Oceans and Fisheries of Korea, and managed by the Korea Marine Environment Management Corporation. This study was also supported by the Korea Institute of Ocean Science and Technology (grant number: PEA0205).
Competing interests
None.
Ethical standards
Because this study conducted a field survey using non-disturbing scuba observations, an ethical review by the Statement Animal Experiment Committee was not required.





