


Introduction
Marine sediments at Bentiaba, Angola, have produced one of the most diverse assemblages of marine amniotes known from the Cretaceous (Mateus et al., Reference Mateus, Polcyn, Jacobs, Araújo, Schulp, Marinheiro, Pereira and Vineyard2012; Fig. 1; Table 1), comparable to the type Maastricht fauna of the Netherlands and Belgium (Jagt, Reference Jagt2005) and the Maastrichtian beds of Morocco (Bardet, Reference Bardet2012). The Bentiaba assemblage is of particular importance because of the high richness of mosasaurs, a group of extinct marine lizards, and other top predators that have been interpreted as reflecting bottom-up selection pressure within a productive ocean ecosystem during the last 32 million years of Cretaceous time (Polcyn et al., Reference Polcyn, Jacobs, Araújo, Schulp and Mateus2014). Bentiaba and other vertebrate fossil localities along the southwest coast of Africa reflect the effects of the growing South Atlantic Ocean on Late Cretaceous marine ecosystems. The Bench 19 Fauna of southern Angola provides a time-constrained glimpse of a marine vertebrate assemblage during the latter phase of this history, placed within the broader context of South Atlantic rifting, sea level, paleoclimate and Late Cretaceous time. In this study we use δ13C values of mosasaur and elasmosaurid plesiosaur tooth enamel to investigate niche differentiation and feeding strategies of the Bench 19 Fauna, and we analyse the geological setting and paleoenvironment of the Bench 19 Bonebed to understand better the biological and physical factors that led to formation of the assemblage (Rogers & Kidwell, Reference Rogers, Kidwell, Rogers, Eberth and Fiorillo2007).

Fig. 1. Location map of Bentiaba. Inset, lower right, shows location of Angola in southwest Africa. Dashed line represents boundary between the Congo Craton and Mesozoic basins, labelled in capital letters. Modified from Strganac et al. (Reference Strganac, Salminen, Jacobs, Polcyn, Ferguson, Mateus, Schulp, Morais, Tavares and Gonçalves2014).
Table 1. Annotated vertebrate fauna list for Bench 19 Bonebed.

Differential carbonate cementation in the Mocuio Formation at Bentiaba creates an alternating pattern of more and less resistant beds. The prominent, resistant beds (or benches) were numbered consecutively beginning with the most basal bench in the Mocuio Formation. Overlying Bench 19 in the stratigraphic section are 10 m of fine sandstone, capped at about 138 m in the section by a series of thin lag deposits composed mainly of comminuted fish bone, as well as numerous shark and mosasaur teeth (Strganac et al., Reference Strganac, Salminen, Jacobs, Polcyn, Ferguson, Mateus, Schulp, Morais, Tavares and Gonçalves2014; Fig. 2). Although fossil remains of mosasaurs, plesiosaurs and marine turtles are distributed throughout the 10 m of the Mocuio Formation overlying Bench 19, vertebrate specimens are particularly concentrated in a 1–2 m thick zone directly overlying Bench 19, referred to here as the Bench 19 Bonebed and Fauna.

Fig. 2. Stratigraphic section at Bentiaba, with Mocuio Formation enlarged. The Ombe Formation is a basalt dated at 84.6 ± 1.5 Ma (Strganac et al., Reference Strganac, Salminen, Jacobs, Polcyn, Ferguson, Mateus, Schulp, Morais, Tavares and Gonçalves2014), within the Santonian. The overlying Baba and Mocuio formations are Campanian–Maastrichtian.
The chronological context for Bentiaba is provided by a δ13C chemostratigraphic curve and magnetic polarity stratigraphy constrained by an 84.6 ± 1.5 Ma 40Ar/39Ar whole-rock radiometric date on a basalt of the Ombe Formation and by ammonite biostratigraphy (Strganac et al., Reference Strganac, Salminen, Jacobs, Polcyn, Ferguson, Mateus, Schulp, Morais, Tavares and Gonçalves2014). The Bench 19 Bonebed is located near the top of the Bentiaba section, in chron C32n.1n between 71.4 and 71.64 Ma (Husson et al., Reference Husson, Galbrun, Laskar, Hinnov, Thibault, Gardin and Locklair2011). Water temperature of 18.5°C for the Bench 19 Fauna was determined from a δ18O chemostratigraphic curve derived from marine bivalves (Strganac et al., in press).
Abbreviations
ML – Museu da Lourinhã, Portugal; MGUAN-PA and PA – Museu de Geologia da Universidade Agostinho Neto, Luanda, Angola (PaleoAngola Collection); SMU – Southern Methodist University, Dallas, USA.
Methods
Locations on the ground of amniote fossils were marked by GPS, recorded in the field and marked on the stratigraphic section. Nearest-neighbour data were compiled from GPS data and the long-axis orientation of the specimens was noted for those that could be identified to taxon and were clearly in place. Remains of bony fish and sharks are found throughout the Mocuio Formation and were collected opportunistically. Precise specimen locality data are on file at SMU. Skeletal completeness and bone modification features were noted during specimen preparation at SMU and ML.
A predicted paleolatitude for Bentiaba from the Ombe Formation dated at 84.6 Ma ± 1.5 Ma (Strganac et al., Reference Strganac, Salminen, Jacobs, Polcyn, Ferguson, Mateus, Schulp, Morais, Tavares and Gonçalves2014) was determined from oriented paleomagnetic samples collected within the measured section at Bentiaba. Results from three basalt samples yield a mean magnetisation direction of declination (D) = 346.9° and inclination (I) = –42.4°, with α95 = 7.8° (Fisher, Reference Fisher1953), which is different from the Earth’s present magnetic field direction at the sampling site (D = 353.2° and I = –56.3°). Paleolatitude (λ) can be calculated by the simple equation:
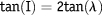 $${\rm{tan(I)}} = {\rm{2tan(\lambda )}}$$
$${\rm{tan(I)}} = {\rm{2tan(\lambda )}}$$
Obtained mean inclination from Ombe Formation basalts yields a paleolatitude of 24.5°S for the Bentiaba locality. The paleolatitude at 71 Ma was a few degrees lower because of the northward drift of Africa.
The paleolatitudinal trace of Bentiaba from the initial opening of the South Atlantic in the Early Cretaceous to its current latitude is presented in Jacobs et al. (Reference Jacobs, Strganac and Scotese2011), determined using PointTracker (Scotese, Reference Scotese2008). The width of the South Atlantic Ocean at 71 Ma and 20°S was determined from Earthworks BV (Reeves, Reference Reeves2014). The bathymetric image of the Angolan continental shelf discussed below was constructed from http://cmtt.tori.org.tw/data/App_map/maplist.htm.
Sediment samples from the Bench 19 Bonebed were taken from the field jacket containing a specimen of Prognathodon kianda (PA 183) at the base of the Bench 19 Bonebed and analysed for mineral composition, grain size and detrital zircon ages. A second sample was taken from the jacket of a Prognathodon sp. (PA 186) near the top of the unit. To determine the mineralogy of the Bench 19 Bonebed matrix, a small amount of material was powdered with a mortar and pestle and analysed at SMU using a Rigaku III Ultima X-ray diffractometer with a step size of 0.05° over a range of 2–60° 2θ. Grain size was determined by sonicating a matrix sample in deionised water to disaggregate it and sieving the dried matrix to separate the size fractions.
A single sample of sandstone (MJP 2006A) was analysed for detrital zircon geochronology to investigate sediment provenance. This sample comprises sand from the Mocuio Formation that was excavated from one of the plaster jackets containing a Prognathodon kianda (PA 183). Detrital zircons were analysed using laser ablation inductively coupled plasma mass spectrometry (LA-ICPMS) at the Arizona LaserChron Center (www.laserchron.org) using methods described by Gehrels et al. (Reference Gehrels, Valencia, Pullen, Loszewski and Huff2006, Reference Gehrels, Valencia and Ruiz2008).
Isolated teeth of mosasaurs and plesiosaurs collected from the Bench 19 Fauna were identified by comparison with specimens at the Shuler Museum of Paleontology at SMU and published literature (Bardet & Pereda Superbiola, Reference Bardet and Pereda Superbiola2002; Schulp et al., Reference Schulp, Polcyn, Mateus, Jacobs, Morais, Da Silva Tavares and Schulp2006, 2008; Polcyn et al., Reference Polcyn, Jacobs, Schulp and Mateus2010; Araújo et al., submitted A; submitted B). Tooth specimens were soaked in 10% acetic acid for at least 72 hours to remove diagenetic carbonate encrustation and rinsed with deionised water. The samples were then sonicated, treated with methanol, rinsed again in a deionised water bath and dried. Enamel was sampled using a carbide-tipped hand drill and the resulting enamel powders were placed in vacuum-sealed reaction vessels with 100% orthophosphoric acid at 25°C for a minimum of 4 hours. The evolved gas was cryogenically purified to isolated CO2, which was analysed with a Finnegan MAT 252 isotope ratio mass spectrometer to determine its δ13C and δ18O values, reported here in permil units relative to the Vienna Pee-Dee Belemnite standard (VPDB). Data obtained for individuals or groups of taxa were compared statistically using the nonparametric Mann–Whitney U test for two groups of data and Kruskal–Wallis ANOVA for three or more groups.
Results
Sedimentology and detrital zircons
The Mocuio Formation at Bentiaba is composed primarily of massive fine-grained sandstone with, near the top, occasional low-angle cross-bedding and some thin fossiliferous lag deposits. Scour marks are minor and rare, and there is no evidence of high-energy deposition or wave action, such as hummocky cross-stratification, indicating that depth was below storm wave base. The macroinvertebrate assemblage is relatively sparse and comprises fragments of the pteriomorphian bivalve Inoceramus, dissolution moulds of non-inoceramid bivalves and irregular vertical burrows.
Although vertebrate fossils occur sporadically throughout the upper Mocuio Formation, they are concentrated in a 1–2 m thick bonebed overlying Bench 19, although the Bench 19 Fauna (Table 1) occurs throughout the overlying 10 m. The base of Bench 19 is planar and the overlying fossil-bearing sediments grade upward rapidly into friable sand that forms steep walls, overlain by more frequent, albeit uncommon, cross-beds and fossiliferous lags. Terrestrial influence in Bench 19 and the overlying 10 m includes rare oxidised, unidentifiable plant remains and occasional isolated dinosaur and pterosaur bones (Mateus et al., Reference Mateus, Polcyn, Jacobs, Araújo, Schulp, Marinheiro, Pereira and Vineyard2012), indicating proximity to shore. No pollen or microfossils were recovered.
The basal Bench 19 Bonebed sediment (sample from PA183; Fig. 3A) comprises a pale yellow (2.5Y 8/2) to white (2.5Y 8/1) poorly sorted, subangular to subrounded, silty, fine-grained feldspathic arenite, weakly cemented by calcite. Clast size ranges from coarse sand to silt-size, with a positively skewed distribution. Larger clasts are lithics and quartz, and smaller clasts are predominantly microcline and anorthite. There is an indistinct and gradual facies change in the upper part of the Bench 19 unit (sample from PA 186). This part of the unit comprises a well-sorted, subrounded to rounded, very fine-grained yellow (2.5Y 7/6) to olive yellow (2.5Y 6/6) silty sand. XRD analysis indicates it is also a feldspathic arenite with a relatively high abundance of quartz (75–90%). This level has a greater textural and compositional maturity relative to the lower part of the Bench 19 Bonebed.

Fig. 3. Bench 19 sediments and zircon age analysis. A. SEM image of sediment samples from main bonebed layer above Bench 19 taken from PA183. Large grains are quartz and small grains are feldspathic. B. Detrital zircon age probability distribution (red) and histogram (blue) for sample MJP2006A from the Bonebed sandstone in the Mocuio Formation. Age of key probability peaks is shown on the figure. C. Concordia plot for detrital zicrons from MJP2006A showing error ellipses from resulting 206Pb/238U and 207Pb/235U values.
Uranium-lead ages were obtained from 76 zircons extracted from a single sample from the Bench 19 Bonebed sandstone with a total age distribution of 107 ± 2 Ma to 2571 ± 7 Ma. A standard age probability distribution (Fig. 3B) shows major grain populations at ∼1996 Ma and ∼1807 Ma, and a late Precambrian to Cambrian population with an age peak at ∼515 Ma. Uranium concentrations for all grains is < 600 ppm. As illustrated in Fig. 3C, some of the Paleoproterozoic grains depart from the concordia, suggesting potential lead loss during the Pan-African orogenic event.
Fossil spatial distribution and orientation
The Bench 19 Fauna was collected from a contiguous band of badland outcrops within an area approximately 500 m north–south by 200 m east–west, with additional specimens obtained from sporadic outcrops extending some 300 m east (Fig. 4A). The Bench 19 unit intersects the eroded surface along a trace approximately 1.5–2 km in length, estimated by connecting the fossil occurrences on a map. The dip of the strata approximates the surface of Bench 19 in some places and in others the intersection is along steep faces. Fragments of pterosaur bones were found throughout the interval, but more complete bones were discovered by their cross-sections in steep outcrop faces, which afforded them protection from deeper weathering and erosion. Bony fish and sharks, although not systematically collected, appear broadly distributed, but shark teeth, especially Squalicorax (Antunes & Cappetta, Reference Antunes and Cappetta2002) are closely intermingled with amniote skeletal remains. A histogram of amniote nearest-neighbour distances shows a peak between 2.5 and 5 m (n = 192, range = 0–35 m,
 $\bar x$
= 8.5, σ = 6.5; Fig. 4B). The closest associations of amniotes (0 m) are interpreted as gut contents because of the position of one individual within the gut area of a presumed consumer and the etched condition of bones and teeth in consumed individuals (see discussion below).
$\bar x$
= 8.5, σ = 6.5; Fig. 4B). The closest associations of amniotes (0 m) are interpreted as gut contents because of the position of one individual within the gut area of a presumed consumer and the etched condition of bones and teeth in consumed individuals (see discussion below).

Fig. 4. A. Satellite image of Bentiaba (map data: Google, DigitalGlobe). Red dots represent locations of specimens recovered from Bench 19 Bonebed. B. Histogram of nearest-neighbour distances among fossils at Bench 19. C. Rose diagram for orientations of elongate fossil elements at Bench 19.
There is no preferred orientation of elements and all specimens rest parallel to the bedding plane (Fig. 4C). The spatial distribution of amniote specimens and their separation by metres indicates a scattered distribution over the sea floor, lacking linear orientation as in a strand, or concentration by currents or other agents into a tight mass. The fossils are not concentrated in ravinements or on scoured surfaces. There is no evidence of turbulence in the sediments or of transport in the condition of the bone.
Bone modification
Amniote fossils occur as partially articulated skeletons, disarticulated but associated partial skeletons and isolated elements. All specimens within the bonebed, except those interpreted as gut contents, exhibit a similar preservation style. Postburial crushing, indicated by longitudinal cracks, is common, but fossils appear more completely permineralised with less apparent postburial crushing in the upper part of the Bench 19 Bonebed. Irrespective of position within the Bench 19 Bonebed, pitting and erosion of cortical bone is present only in juvenile plesiosaur limbs. No bioencrustation or microboring was observed. Evidence of transport (abrasion, rounding or polishing) is absent and bone surfaces are generally pristine (Fig. 5A).

Fig. 5. A. Bite marks attributed to the shark Squalicorax pristodontus on a mosasaur rib from the Bench 19 Bonebed, enlarged in B. Note short transverse marks along major groove from serrations on teeth. C. Specimen PA183, Prognathodon kianda, with preserved gut contents. Two specimens in the gut are shown as highlighted regions 1 and 2, and are enlarged in D and E, respectively. A third specimen was recovered between the blocks and is not shown.
Most specimens show evidence of scavenging by sharks. Bite marks and scratches most closely match the size and serration pattern of Squalicorax pristodontus teeth, displaying secondary lineations perpendicular to the main grooves (Schwimmer et al., Reference Schwimmer, Stewart and Williams1997; Shimada et al., Reference Shimada, Tsuihiji, Sato and Hasegawa2010). Marks from shark scavenging are common on limb elements and ribs but rare on vertebrae, irrespective of taxon (Fig. 5B).
Mosasaur-on-mosasaur predation or scavenging is present in one specimen of Prognathodon kianda (PA 183), which comprises the skull, some proximal rib portions and the vertebral column complete to the anterior caudals, but is missing the limbs, girdles and most of the ribs. The skeleton shows bite marks attributed to sharks and numerous Squalicorax pristodontus teeth were found with the specimen. Partial skulls of ‘Platecarpus’ ptychodon, Mosasaurus sp. and an indeterminate mosasaurine were found in the gut region of PA 183 (Fig. 5C). In these gut-content specimens, the skull roof, snout and other areas thinly fleshed in life are etched, and tooth crowns are dissolved above the gum line.
A second mass of bones exhibiting similar modification to those seen in the gut area of PA 183 comprises the partial skeletons of three Halisaurus specimens (PA 179) and a juvenile P. kianda (PA 25). Individual bones show no modification by sharks. At the time of discovery and excavation, the mass was thought to be isolated and a consumer remains undiscovered, possibly still in the quarry. The size of the bone mass, if in fact it is gut contents, suggests the consumer was a large mosasaur.
Results of δ13C
The 49 enamel carbonate δ13C values derived from 37 tooth specimens of five mosasaur taxa and four plesiosaur teeth from Bench 19 Bonebed are presented in Table 2 and Fig. 6. PA 314 (Halisaurus) yielded a δ13C value of –7.2‰. Specimens PA 177 and PA 312 (‘P.’ ptychodon) produced an average δ13C value of –7.3‰. Twenty-one teeth assigned to Prognathodon kianda produced an average δ13C value of –10.1‰ and ranged from –14.1 to –5.2‰. Three teeth of Mosasaurus sp. yielded an average δ13C value of 11.6‰ and ranged from –12.9 to –10.2‰. Eleven Globidens phosphaticus teeth yielded an average δ13C value of 13.0‰ and ranged from –16.1 to –10.8‰. Four plesiosaur teeth produced an average δ13C of –13.1‰ and ranged from –14.0 to –12.2‰.
Table 2. δ13C values of tooth enamel carbonate in Bench 19 mosasaurs and plesiosaurs. Length estimates from Polcyn et al. (Reference Polcyn, Jacobs, Araújo, Schulp and Mateus2014).


Fig. 6. δ13C values of mosasaurs and plesiosaurs at Bench 19.
Halisaurus and ‘P.’ ptychodon are grouped together for analyses due to their small body size (<5 m in length; Polcyn et al., Reference Polcyn, Jacobs, Araújo, Schulp and Mateus2014) and their similar δ13C values. The standard deviation of Halisaurus + ‘P.’ ptychodon (–7.3 ± 0.4) does not overlap with those of Prognathodon (–10.3 ± 2.4), Globidens (–13.1 ± 1.6) or plesiosaurs (–13.1 ± 0.7) and is therefore statistically different.
Mann–Whitney U tests (Table 3) were used to compare δ13C values between two taxa or groups at significance levels of 0.05. Globidens values are significantly different from Prognathodon (p = 0.001), but not from plesiosaurs (p = 0.862) or Mosasaurus (p = 0.223). Mosasaurus is not significantly different from Prognathodon (p = 0.316) or plesiosaurs (p = 0.114). The δ13C values of Prognathodon are not significantly different from ‘P.’ ptychodon + Halisaurus (p = 0.046).
Table 3. Results for statistical analysis of variance for marine amniote δ13C values.

In comparing the δ13C values between more than two taxa, Kruskal–Wallis ANOVA tests indicate Globidens, Mosasaurus and plesiosaurs were not significantly different (p = 0.293). The group is significantly different when Prognathodon is included (p = 0.003). There is a significant difference in δ13C values derived from Mosasaurus, Prognathodon and plesiosaurs (p = 0.046), and Globidens, Prognathodon and plesiosaurs (p = 0.001).
Discussion
The modern marine ecosystem off the shore of southern Angola and Namibia is highly productive due to the Benguela Current and coastal upwelling system (Shannon & Nelson, Reference Shannon, Nelson, Wefer, Berger, Siedler and Webb1996). The Benguela upwelling zone provides nutrient-rich waters that support a diverse marine ecosystem containing ∼70% of the marine mammal species in the South Atlantic Ocean (Cury et al., Reference Cury, Bakun, Crawford, Jarre, Quiñones, Shannon and Verheye2000). Although the modern Benguela upwelling system is purported to have originated in the late Miocene (Diester-Haas et al., Reference Diester-Haas, Meyers and Vidal2002), the presence of organic-rich Cenomanian–Turonian deposits offshore in western Africa and the rich marine vertebrate fauna preserved in Turonian, Campanian and Maastrichtian coastal outcrops of Angola indicate that latitudinally controlled, high-productivity environments consistent with upwelling existed in the Late Cretaceous (Handoh et al., Reference Handoh, Bigg, Jones and Inoue1999; Jacobs et al., Reference Jacobs, Mateus, Polcyn, Schulp, Antunes, Morais and Da Silva Tavares2006, Reference Jacobs, Mateus, Polcyn, Schulp, Scotese, Goswami, Ferguson, Robbins, Vineyard and Buta Neto2009; Polcyn et al., Reference Polcyn, Jacobs, Araújo, Schulp and Mateus2014).
The paleolatitude of Bentiaba, at the time of the accumulation of the Bench 19 Fauna (C32n.1n; 71.40–71.64, using the calibration of Husson et al., Reference Husson, Galbrun, Laskar, Hinnov, Thibault, Gardin and Locklair2011; see Strganac et al., Reference Strganac, Salminen, Jacobs, Polcyn, Ferguson, Mateus, Schulp, Morais, Tavares and Gonçalves2014) was not more than the –24° as calculated for the Ombe Formation basalt (84.6 ±1.5 Ma 40Ar/39Ar whole-rock radiometric date; Strganac et al., Reference Strganac, Salminen, Jacobs, Polcyn, Ferguson, Mateus, Schulp, Morais, Tavares and Gonçalves2014) and as reported by Jacobs et al. (Reference Jacobs, Strganac and Scotese2011). The width of the South Atlantic at paleolatitude 20°S and 71 Ma was approximately 2700 km calculated from Reeves (Reference Reeves2014), roughly half the current width. Thus, the Bench 19 Fauna was emplaced at a paleolatitude analogous to the position of Walvis Bay in the present-day latitudinal position of Africa, although in a wide but narrower than present-day ocean. This latitude is well within the area of the high-pressure, descending limb of the southern Hadley cell, driving rich upwelling along the coastal desert. Oceanic upwelling cells provide nutrients that enhance primary productivity, which has been suggested as a driver of mosasaur evolution generally (Jacobs et al., Reference Jacobs, Mateus, Polcyn, Schulp, Scotese, Goswami, Ferguson, Robbins, Vineyard and Buta Neto2009; Polcyn et al., Reference Polcyn, Jacobs, Araújo, Schulp and Mateus2014). The presence of abundant marine amniote fossils in coastal Angola, specifically at Bentiaba, is consistent with that hypothesis.
The continental shelf at Bentiaba is much narrower than in northern Namibia (100–160 km in Walvis Bay, as opposed to <10 km at Bentiaba; Fig. 7). The extremely narrow width of the continental shelf and the orientation of the continental margin in the Bentiaba region are controlled by the tectonics of the Benguela and Lucapa transform faults, relating to the opening of the South Atlantic (as discussed by Guiraud et al., Reference Guiraud, Buta-Neto and Quesne2010; see also Moulin et al., Reference Moulin, Aslanian, Rabineau, Patriat, Matias, Mohriak, Danforth, Post, Brown, Tari, Nemcok and Sinha2012). The depth of the shelf near Bentiaba was shallow throughout the Late Cretaceous (Strganac et al., Reference Strganac, Salminen, Jacobs, Polcyn, Ferguson, Mateus, Schulp, Morais, Tavares and Gonçalves2014). The 84.6 Ma Ombe Formation basalt flowed across the coastline, but based on paleomagnetic and δ13C chemostratigraphic correlations (Strganac et al., Reference Strganac, Salminen, Jacobs, Polcyn, Ferguson, Mateus, Schulp, Morais, Tavares and Gonçalves2014), the Bentiaba coast was emergent as indicated by the missing section shown by 5 my of time not represented by strata subjacent to the Ombe Formation and 6 my of missing strata superjacent to a thin veneer of sediment atop the Ombe Formation. Examination of sea-level curves presented by Müller et al. (Reference Müller, Sdrolias, Gaina, Steinberger and Heine2008) and Miller et al. (Reference Miller, Mountain, Wright and Browning2011) does not suggest an apparent link to eustatic sea-level fall as the cause for these hiatuses, but tectonically induced stratigraphic hiatuses have been documented in the Benguela basin to the north (Giraud et al., Reference Guiraud, Buta-Neto and Quesne2010). At Bentiaba, the two Late Cretaceous episodes of relative sea-level fall were caused by local to regional uplift. The intervening Ombe Formation defines sea level at the time by crossing the Santonian shoreline. The fossiliferous upper Campanian to lower Maastrichtian Baba Formation, which overlies the unconformity, and the Mocuio Formation, which contains the Bench 19 Fauna, represent shelf flooding resulting from relaxation and subsidence. The narrow shallow shelf may have served to concentrate prey items and attract predators in a relatively small area compared to the shelf to the north and south. Cenozoic faulting and eustatic sea level change culminated in a Pliocene or younger terrace that truncates Cretaceous deposits at Bentiaba, perhaps comparable to terraces seen farther south in the Namibe basin (Sessa et al., Reference Sessa, Callapez, Dinis and Hendy2013).

Fig. 7. Bathymetric map of southwest Africa. Arrow points to Bentiaba. Dashed lines indicate fracture zones (FZ), which can be traced to transform faults at the Mid-Atlantic Ridge. South of the Walvis Ridge is Walvis Bay. Note the width of the continental shelf at Bentiaba compared to Walvis Bay. In the early Maastrichtian (Late Cretaceous), Bentiaba lay approximately 10° further south at a latitude analogous to that of modern Walvis Bay.
The bonebed sandstone is compositionally immature and texturally submature. Its poor sorting, incomplete rounding and feldspathic composition indicate close proximity to a granitic source. The suite of ages exhibited by detrital zircons (Fig. 3B) show significant populations at 1996 Ma and 1807 Ma with lesser populations at 515 and ∼2500 Ma. The observed age probability distribution is consistent with a nearby source terrain for the Mocuio Formation sandstones. Within close proximity to the present-day outcrops, Angolan basement yields radiometric ages of 1795–2243 Ma (Hanson, Reference Hanson, Yoshida, Windley and Dasgupta2003). Angolan basement is dominated by Paleoproterozoic granitic and gneissic lithologies with inliers of Archean crust consistent with the dominance of Paleoproterozoic detrital zircon ages (62 of the 76 grains studied) and occasional Archean grains. Granitoid rocks of the Pan-African Kaoko Belt, south of Bentiaba, yield radiometric ages of 505–656 Ma (Hanson, Reference Hanson, Yoshida, Windley and Dasgupta2003; Goscombe et al., Reference Goscombe, Gray, Armstrong, Foster and Vogl2005). The youngest detrital zircon grains at 107 Ma and 132 Ma are significantly older than the depositional age of the Mocuio Formation. They may represent minor sediment derivation from Early Cretaceous kimberlites and carbonatites emplaced along the Lucapa fault zone (Giuraud et al., Reference Guiraud, Buta-Neto and Quesne2010). While not providing a unique fingerprint of sediment provenance, detrital zircon ages from the bonebed sandstone are consistent with derivation from nearby basement outcrops and do not require long distance sediment transport. The detrital zircon data show that the Precambrian basement was exhumed by the Cretaceous and that Bench 19 sedimentary rocks could have been derived from a nearby source area. Thus, mineralogy, physical characteristics of grains, detrital zircon ages and proximity of the granitic shield to the coast at Bentiaba all indicate sediment transport by short intermittent or seasonal rivers draining the shield and flowing directly to the sea. This is consistent with the sedimentation in the arid environment seen today in northern Namibia (Calvert & Price, Reference Calvert, Price, Suess and Thiede1983). However, in northern Namibia, a 70-km band of organic-rich diatom ooze is deposited at depths shallower than 140 m (Calvert & Price, Reference Calvert, Price, Suess and Thiede1983) and along the shelf where terrigenous input from the coastal arid region is insufficient to dilute planktonic and skeletal organic matter. At Bentiaba, there is no evidence of diatom ooze.
The Bench 19 unit was deposited relatively near the shore based on the paleogeography of the Bentiaba coast, as shown by the shoreline observed in the Ombe Formation, the narrow shelf, and the composition and texture of the sediment. On the other hand, it is interpreted to have been deposited below storm wave base (Peters & Loss, Reference Peters and Loss2012) because sedimentary structures such as hummocky cross-stratification and significant scour surfaces are absent. Furthermore, there is no evidence of postmortem, pre-burial, subaerial weathering on the bones of the marine species (Behrensmeyer, Reference Behrensmeyer1978), indicating that the life, death and burial of these animals was completely below the surface of the ocean. While the Bench 19 Bonebed preserves a range of completeness of specimens, none appears to have bloated and floated from gases released during decomposition. The presence of relatively complete carcasses in the bonebed suggests that the volume of gas generated and captured in the carcasses was insufficient to overcome hydrostatic pressure, leaving the carcasses on the bottom. Reisdorf et al. (Reference Reisdorf, Bux, Wyler, Benecke, Klug, Maisch, Fornaro and Wetzel2012), in their study of ichthyosaur taphonomy and bathymetry, concluded that intact articulated carcasses remaining on the sea bed due to hydrostatic pressure at certain localities were indicative of water depths >50 m because putrifying carcasses sometimes rise from depths of 50 m but do not rise from depths of 100 m or more. Variability in the tendency to rise, especially at shallower depths, is temperature dependent due to the solubility of gases. The water temperature at Bentiaba immediately below Bench 19 as determined from δ18O of Inoceramus shells was 18.5°C (Strganac et al., in press). Assuming a maximum shelf depth of 150 m, a reasonable range of depths for the formation the Bench 19 Bonebed is below wave base, between 50 and 150 m.
The amniote fauna of the Bench 19 Bonebed was reviewed by Mateus et al. (Reference Mateus, Polcyn, Jacobs, Araújo, Schulp, Marinheiro, Pereira and Vineyard2012) and an updated, annotated faunal list is provided in Table 1. Schulp et al. (Reference Schulp, Vonhof, van der Lubbe, Janssen and van Baal2013) documented the presence of a large Prognathodon sp. and Carinodens sp., neither of which is known from the Bench 19 Fauna, but rather from higher stratigraphic levels. Lower in the section, in the Baba Formation, below the Bench 19 unit, a small russellosaurine mosasaur (sensu Polcyn & Bell, Reference Polcyn and Bell2005) is found. Russellosaurina is the clade that includes the subfamilies Plioplatecarpinae and Tylosaurinae, representatives of which are found in the Bench 19 Fauna, but the Baba specimen is distinct from those found higher in the section. Thus, the amniote fauna from the Bench 19 Bonebed is taxonomically consistent but distinct taxonomically from levels above and below Bench 19. In addition, above the Bench 19, the bivalve Inoceramus, common in the bonebed, has its last appearance in the section, nurse sharks (Ginglymostomatidae) have their first appearance and Squalicorax teeth obtain a larger maximum size co-occurring with smaller forms. The taxonomic internal consistency of the Bench 19 Fauna and the short time bin into which it fits, compared to the fauna above and below, suggests accumulation of the Bench 19 Bonebed on an ecological rather than a geological timescale.
The fossiliferous fine-grained sandstone Bench 19 Bonebed in the upper Mocuio Formation preserves an attritional fossil assemblage, as indicated by the stratigraphic distribution of specimens and a faunal list that includes pterosaurs and dinosaurs as rare and isolated elements among an overwhelmingly marine fauna. Attritional accumulations forming bonebeds have an excess of bones relative to sediment input, and hence exhibit some amount of time averaging. There is no evidence of past exhumation and reburial of individual fossils within the Bench 19 Bonebed, consequently the stratigraphic position of fossils within the bonebed is a time-ordered sequence and the fauna must be considered time-averaged in the strictest sense, of a relatively short duration. Temporal limits on the duration of the Bench 19 Bonebed are set by the length of chron C32n.1n, which is the 240 ky bin between 71.4 and 71.64 Ma (Strganac et al., Reference Strganac, Salminen, Jacobs, Polcyn, Ferguson, Mateus, Schulp, Morais, Tavares and Gonçalves2014).
The Bench 19 Fauna does not appear to be a catastrophic death assemblage such as reported in marine mammal strandings caused by harmful algal blooms (Brongersma-Sanders, Reference Brongersma-Sanders1948, Reference Brongersma-Sanders and Hedgepeth1957; Brongersma-Sanders et al., Reference Brongersma-Sanders, Stephan, Kwee and DeBruin1980; Pyenson et al., Reference Pyenson, Gutstein, Parham, Le Roux, Carreño Chavarría, Little, Metallo, Rossi, Valenzuela-Toro, Velez-Juarbe, Santelli, Rogers, Cozzuol and Suárez2014) because at Bentiaba individuals do not align with strand, the bones are not limited to a single plane and there is definitive evidence of active predation or scavenging. Pyenson et al. (Reference Pyenson, Irmis, Lipps, Barnes, Mitchell and McLeod2009) studied the formation of the Miocene Shark Tooth Hill Bonebed of California and concluded that this rich accumulation, which is both larger and denser than the Bench 19 Bonebed, was a winnowed, attritional accumulation formed under sediment-starved conditions and terminated when sediment input increased. The time averaged in the Shark Tooth Hill Bonebed is as much as 700 ky. At Bentiaba there is no evidence of significant winnowing and mixing by bottom currents, large specimens are partially articulated and scavenged, and biological interactions related to feeding are clear.
Bench 19 does not fit the model of preservation in a subaqueous gravity flow as presented by Adams (Reference Adams2009) because the shelf setting of Bentiaba is inconsistent with the geological setting and depositional environment reflected in sedimentary textures and structures expected of active margins. However, the Pisco Formation along the coast of western South America in latitudes comparable to the paleolatitude of Bentiaba is also on an active margin, yet preserves a diverse cetacean fauna, including articulated skeletons with baleen. Sediments are both diatomaceous and tuffaceous, and most fossil occurrences appear to be rapidly buried falls (Esperante et al., Reference Esperante, Brand, Chadwick, Poma, De Renzi, Alonzo, Belinchon, Penalver, Montoya and Marquez-Aliaga2002, Reference Esperante, Brand, Nick, Poma and Urbina2008; Brand et al., Reference Brand, Esperante, Chadwick, Porras and Alomía2004, Reference Brand, Urbina, Chadwick, DeVries and Esperante2011) or strandings (Pyenson et al., Reference Pyenson, Gutstein, Parham, Le Roux, Carreño Chavarría, Little, Metallo, Rossi, Valenzuela-Toro, Velez-Juarbe, Santelli, Rogers, Cozzuol and Suárez2014) in embayments. Along passive margins, Calvert Cliffs, Maryland, preserves a rich cetacean fauna, but the accumulation is attritional over millions of years (Vogt & Eshelman, Reference Vogt and Eshelman1987; Gottfried et al., Reference Gottfried, Bohaska, Whitmore, Berta and Deméré1994). In Africa, the Cretaceous upwelling area along Morocco is the antitropical equivalent of that which occurred at the same time along the Bentiaba coast and it has produced a rich and varied marine fauna (Bardet et al., Reference Bardet, Pereda Suberbiola, Jouve, Bourdon, Vincent, Houssaye, Rage, Jalil, Bouya and Amaghzaz2010; Bardet, Reference Bardet2012). However, upwelling in Morocco produced phosphatic sediments and specimens are not concentrated as in the Bench 19 Fauna.
In the case of the Bench 19 Bonebed, it is clear that a large number of carcasses accumulated individually soon after death, presenting a food source that attracted large numbers of mobile scavengers, as evidenced by numerous associated shark teeth. Decomposition was truncated by burial at varying stages, but none of the carcasses remained on the seafloor long enough to transition beyond the mobile scavenger phase (sensu Smith & Baco, Reference Smith and Baco2003).
Stable carbon isotopes
Whole-body δ13C values of consumer species largely reflect the carbon isotope values of primary producers on which they ultimately feed, albeit with an ∼0–1‰ enrichment in 13C per trophic step (Koch, Reference Koch, Michener and Lajtha2007). The δ13C of tooth enamel carbonate, as opposed to whole-body values, is typically ∼9‰ more positive relative to diet, but differences in the δ13C of tooth enamel carbonate among marine species still largely reflects the δ13C values of primary producers (Koch, Reference Koch, Michener and Lajtha2007). In marine settings the δ13C values of primary producers and particulate organic matter become depleted with increasing distance from shore and depth, possibly due to a more diverse group of primary producers found in highly productive nearshore environments, including 13C-enriched kelp beds and benthic macrophytes, relative to offshore where 13C depleted marine phytoplankton are the dominant primary producers (Clementz & Koch, Reference Clementz and Koch2001; Michener & Kaufman, Reference Michener, Kaufman, Michener and Lajtha2007). Clementz & Koch (Reference Clementz and Koch2001) used these relationships to examine the δ13C values derived from tooth enamel belonging to modern and recent marine mammals. They demonstrated that a variety of niches were used in the California upwelling zone and reflected the trend of depleted δ13C values in primary producers located farther from shore (Fig. 8A).

Fig. 8. Comparison of δ13C values of (A) modern primary producers and particulate organic matter (POM), (B) modern marine mammals (modified from Clementz & Koch, Reference Clementz and Koch2001), and (C) tooth enamel carbonate of Bench 19 mosasaurs and plesiosaurs.
Robbins et al. (Reference Robbins, Ferguson, Polcyn and Jacobs2008) examined stable carbon isotopes from a diverse sample of mosasaur teeth from numerous Upper Cretaceous localities and interpreted the δ13C values as reflecting feeding distance from shore and diving duration, showing that the δ13C values generally correlated with body size. Schulp et al. (Reference Schulp, Vonhof, van der Lubbe, Janssen and van Baal2013) examined five mosasaur taxa from the type Maastrichtian section and found a strong correlation between larger body size and lower δ13C values.
The δ13C values derived from tooth enamel (Fig. 6, Table 2) from the Bench 19 Fauna show a general trend of decreasing average δ13C values with increasing body size, the pattern previously documented in mosasaurs (Robbins et al., Reference Robbins, Ferguson, Polcyn and Jacobs2008; Schulp et al., Reference Schulp, Vonhof, van der Lubbe, Janssen and van Baal2013). Halisaurus and ‘P.’ ptychodon exhibit enriched δ13C values similar to modern sea otters and reflect a shallow marine feeding habitat at the coastline. δ13C values of Mosasaurus and Prognathodon are similar in value (∼–12 to –8‰) to cetaceans and pinnipeds that feed in nearshore California waters (Clementz & Koch, Reference Clementz and Koch2001). Prognathodon teeth from Bentiaba have a much broader range of δ13C values (–14.8 to –5.2‰) than those reported from the Netherlands (–13.6 to –12.3‰; Schulp et al., Reference Schulp, Vonhof, van der Lubbe, Janssen and van Baal2013). This greater range may be a product of the larger sample size in this study, or it may indicate that Bentiaba mosasaurs used a greater variety of habitats or had broader diets. Average Globidens δ13C values are more depleted than expected for their body size, and are statistically indistinguishable from those of plesiosaurs. The δ13C values of plesiosaurs are consistent with foraging far from shore (Cicimurri & Everhart, Reference Cicimurri and Everhart2001), but this does not explain the values seen in Globidens. The relatively depleted δ13C values in Globidens are similar to northern fur seals and elephant seals, whereas the values of the latter species are probably affected by routine dives >300 m in depth (Clementz & Koch, Reference Clementz and Koch2001). Thus, Globidens δ13C values can be explained by foraging behaviour that involved increased dive time (Biasatti, Reference Biasatti2004) required by a diet of mollusks consistent with reported stomach contents (Martin, Reference Martin, Martin and Parris2007; Martin & Fox, Reference Martin, Fox, Martin and Parris2007) and with durophagus tooth morphology.
The δ13C values of Bench 19 mosasaurs indicate that they occupied a variety of niches as do modern marine mammals in the upwelling waters off California. At least six mosasaur taxa are represented in the Bench 19 Fauna, five of which were analysed for δ13C (Table 2). δ13C values of tooth enamel, body size and tooth-form disparity suggest niche differentiation in preferred foraging zones ranging from nearshore to open ocean. Although differentiation of foraging strategies would be expected in a rich upwelling zone, the five Bench 19 mosasaur taxa are associated in a short time horizon and a small area, and are not spatially segregated as the δ13C values predict.
The presence of Halisaurus and ‘P.’ ptychodon, both characterised by relatively small body size and positive δ13C values, is consistent with the near-shore depositional environment of the Bench 19 Bonebed. Likewise, the presence of the durophagous mosasaur Globidens can be explained by the abundance of the bivalve Inoceramus in the strata in which it is found; the predator–prey relationship between these taxa is established by Globidens stomach contents (Martin & Fox, Reference Martin, Fox, Martin and Parris2007). The stomach contents of Prognathodon kianda demonstrate the association of three of the five mosasaur species in the Bench 19 Fauna as a function of predation or scavenging.
Prognathodon kianda shows a wide range of δ13C values and it had remains of three mosasaur individuals in its gut area. The biological association of scavenging sharks with a diversity of mosasaurs and plesiosaurs, coupled with the association of three species of mosasaurs within the gut of a fourth, implies that all Bench 19 mosasaurs were contemporary inhabitants of this highly productive shallow, narrow shelf environment.
The association of mosasaur taxa with each other and with the sharks that scavenged their flesh is indicative of the use of this area as a foraging zone by individual taxa either as a relatively permanent home range or as an opportunistic feeding ground.
Conclusions
The Bench 19 Bonebed at Bentiaba, Angola, formed in a shallow, narrow, highly productive shelf environment characterised by relatively low sedimentation rates compared to bone input. The Bench 19 Bonebed represents a residual accumulation of instantaneous biological interactions that introduced carcasses into the sedimentary system where the animals lived, at the time they died, and in that sense, it is attritional. Although we cannot constrain the amount of time represented to less than 240,000 years, we interpret the Bench 19 Fauna as an accurate representation of the Late Cretaceous marine amniote ecosystem in offshore Angola. The Bench 19 Bonebed contains the richest, temporally constrained and intimately associated assemblage of mosasaurs yet known from the fossil record, and as such it provides an unparalleled window into the ecological relationships of Late Cretaceous marine ecosystems on the southwest African coast of the widening South Atlantic Ocean.
Acknowledgements
This publication results from Projecto PaleoAngola, an international cooperative research effort among the contributing authors and their institutions, funded by the National Geographic Society, the Petroleum Research Fund of the American Chemical Society, Sonangol E.P., Esso Angola, Fundação Vida of Angola, LS Films, Maersk, Damco, Safmarine, ISEM at SMU, the Royal Dutch Embassy in Luanda, TAP Airlines, Royal Dutch Airlines and the Saurus Institute. Detrital zircons were analysed courtesy of the ExxonMobil Upstream Research Company. We thank Margarida Ventura and André Buta Neto for providing our team with help in the field. Tako and Henriette Koning provided valuable support and friendship in Angola. We are grateful to Diana Vineyard for her facilitation of this study. Roy Beavers generously aided with SEM and mineral analysis.












{kind=link}