



Introduction
Lusitanian-type marine mollusc faunas containing Bittium reticulatum s.l., Acanthocardia paucicostata and Polititapes (formerly Venerupis) senescens are often found in Eemian (Marine Isotope Stage MIS5e) sediment records in the southern North Sea Basin and are also known from the Eemian of the western Baltic region (Funder et al., Reference Funder, Demidov and Yelovicheva2002). They are commonly used as a regional marker assemblage for the Eemian period (see Spaink, Reference Spaink1958; Van Leeuwen et al., Reference Van Leeuwen, Beets, Bosch, Burger, Cleveringa, van Harten, Herngreen, Kruk, Langereis, Meijer, Pouwer and de Wolf2000; Cleveringa et al., Reference Cleveringa, Meijer, van Leeuwen, de Wolf, Pouwer, Lissenberg and Burger2000; Wesselingh et al., Reference Wesselingh, Rijken, van Nieulande and Pouwer2012; Meijer et al., Reference Meijer, Pouwer, Cleveringa, de Wolf, Busschers and Wesselingh2021). Here, we report the occurrence of very similar faunas found at 20–30 m below MIS6 related glacial till in the northern Netherlands. Our finds have implications for the use of Bittium-dominated faunas as a unique regional biostratigraphic datum and point to hitherto unknown very warm marine conditions in Middle Pleistocene interglacial times in the North Sea Basin.
Material and methods
Borehole B11D2148 (RDX 195.178; RDY 556.799) is located near the village of Luxwoude, Friesland, northern Netherlands (Fig. 1). The surface level is at 1.30 m below Dutch Ordinance Datum and the termination depth of the borehole is 285 m below surface (b.s.). The drilling was performed in April 2018 by De Ruiter Boringen en Bemalingen B.V. using the air-lift method. This method retrieves bulk sediment samples that represent 1-m intervals (see Fig. 2). The samples were described and analysed at the TNO-core lab by P.C. van der Klugt following the SBB5.3 sample description procedure (expanded from Bosch, Reference Bosch2000; borehole data available at www.dinoloket.nl). Attribution of the sampled sequences into lithostratigraphic units followed criteria as outlined in TNO-GSN (2021). For mollusc analysis, approximately 300 g of dried (sandy) sediment was sieved over a 1 mm sieve. Species were identified and preservation characteristics were noted in order to distinguish poorly preserved (indicative of reworking) and well-preserved (possibly indicative of in-situ or proximal reworking) shells and fragments, following the approach described in Meijer et al. (Reference Meijer, Pouwer, Cleveringa, de Wolf, Busschers and Wesselingh2021).
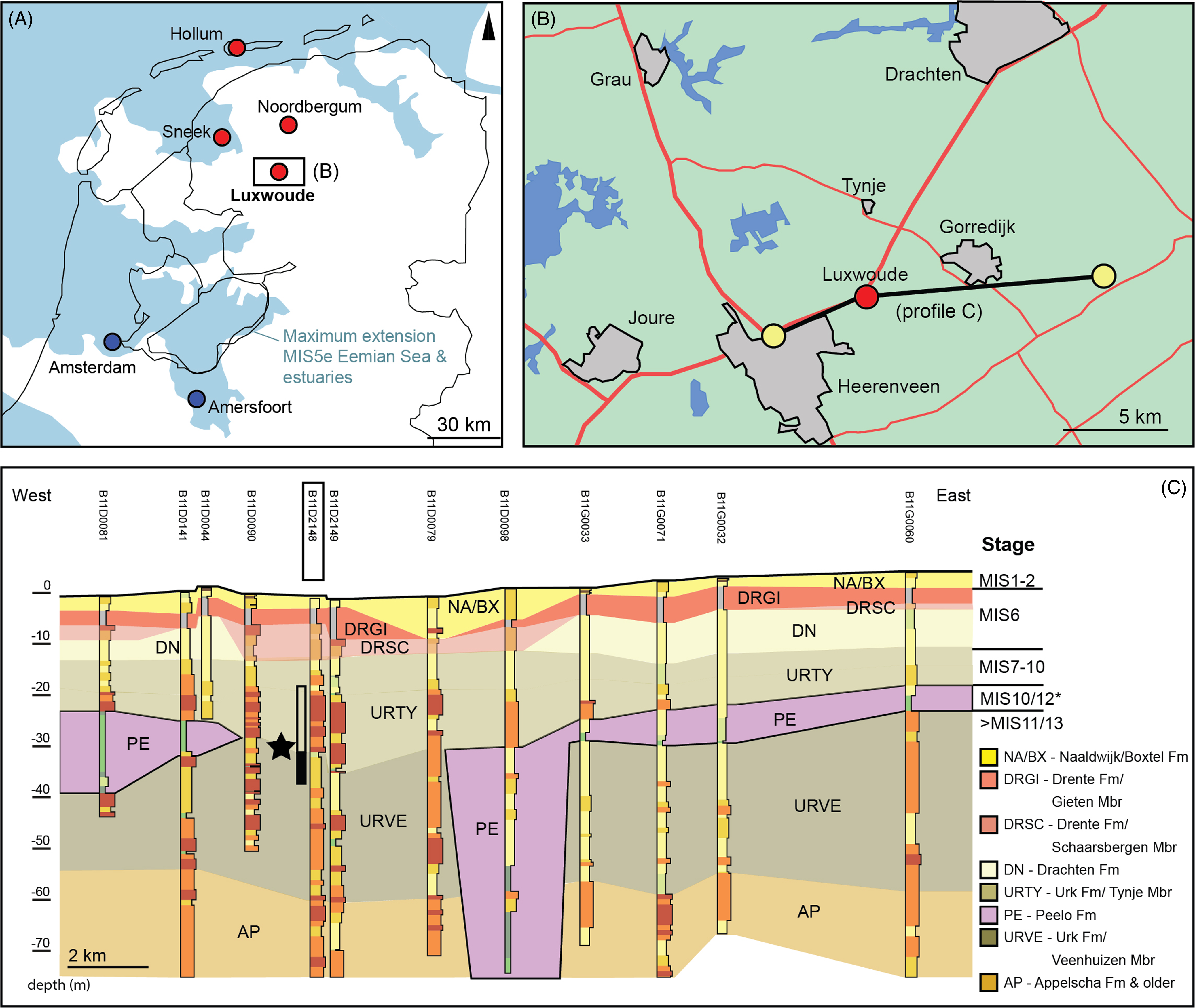
Fig. 1. Regional stratigraphic context of borehole Luxwoude. (A) Location of the Luxwoude borehole (B11D2148), localities mentioned in text and maximum extent of MIS5e Eemian sea (in blue, adapted from Busschers et al., Reference Busschers, Wesselingh, Kars, Versluijs-Helder, Wallinga, Bosch, Timmner, Nierop, Meijer, Bunnik and de Wolf2014). (B) Location of regional cross-section (C) with mollusc intervals in borehole Luxwoude indicated with black star. Relative position of the lower shelly interval (URVE, Veenhuizen Mbr) must be older than MIS10/MIS12 age Peelo Formation (*for discussion see text). The overlying shelly interval (Tynje Mbr, Urk Fm) probably represents faunas from the eroded Veenhuizen Member interval.
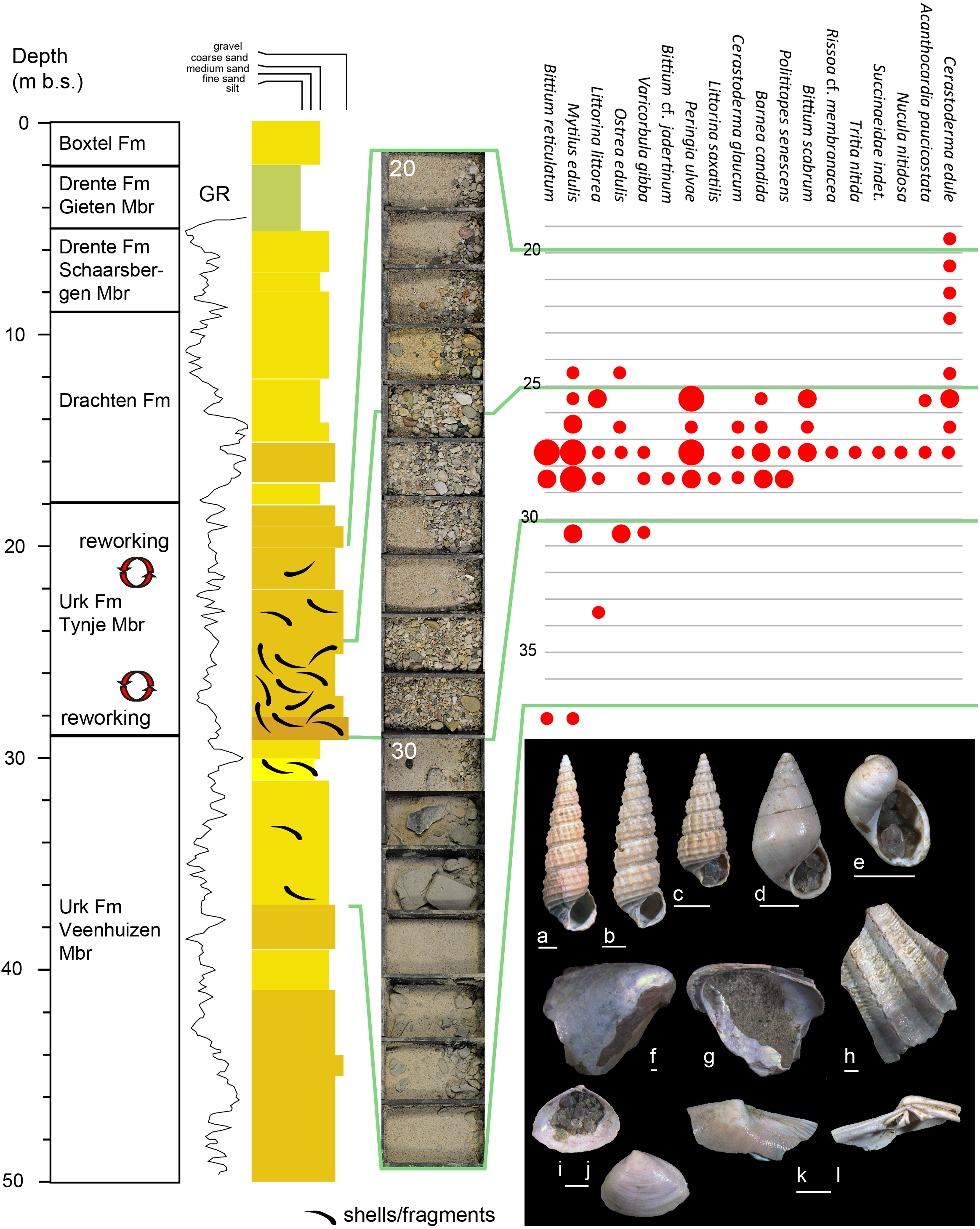
Fig. 2. Sediment samples, lithostratigraphy and mollusc fauna of Late Quaternary intervals of borehole B11D2148 (Luxwoude) and representative mollusc species found in this drilling. GR – Gamma Ray. Small red dots denote a single occurrence, intermediate between 2 and 5 fragments/specimens and large dots over 5 fragments/specimens. (a) RGM.962642 Bittium reticulatum (37–38 m b.s.), (b) RGM.962647 Bittium scabrum (27–28 m b.s.), (c) RGM.962644 Bittium cf. jadertinum (28–29 m b.s.), (d) RGM.962647 Peringia ulvae (27–28 m b.s.), (e) RGM.962648 Succinaeidae indet. (27–28 m b.s.), (f, g) RGM.962643 Mytilus edulis (30–31 m b.s.), (h) RGM.962650 Acanthocardia paucicostata (27–28 m b.s.), (i, j) RGM.962645 Varicorbula gibba (28–29 m b.s.), (k, l) RGM. 962646 Polititapes senescens (28–29 m b.s.). Scale bars 1 mm.
Geological succession and faunas
The geological framework of the subsurface near borehole B11D2148 is captured by the DGM geological model and the REGIS II hydrogeological model (https://www.dinoloket.nl/en/subsurface-models version 2.2). Both nation-wide models are based on a limited amount of borehole data. For this study, a more detailed cross-section has been assembled using an additional set of regional boreholes as available via https://www.dinoloket.nl/en/subsurface-data. The cross-section (Fig. 1C) shows the lithostratigraphic record down to a depth of 70 m. Near the surface Holocene and Late Pleistocene sediments occur. Marine Eemian sediments – stratigraphically defined near Amersfoort in the central Netherlands and closest occurrence near Sneek to the NW (Fig. 1, Busschers et al., Reference Busschers, Wesselingh, Kars, Versluijs-Helder, Wallinga, Bosch, Timmner, Nierop, Meijer, Bunnik and de Wolf2014) – have not been deposited near Luxwoude (Fig. 1A). Below the Late Pleistocene succession, glacial till, fluvioglacial and periglacial intervals are present that were deposited during MIS6 (Drente and Drachten Formation: detailed descriptions and interpretations can be found at www.dinoloket.nl/stratigrafische-nomenclator). Underlying these glacial units a succession of fine gravelly sands with mollusc remains occurs, which has been attributed to the Urk Formation (cf. Bosch, Reference Bosch1990). The Urk Formation contains an upper gravelly coarse sandy interval (19–29 m b.s.) attributed to the Tynje Member and a lower interval (29–59 m b.s.) with a mostly fine sandy top attributed to the Veenhuizen Member (see below). The Urk Formation is predominantly composed of Rhine-derived sediments which are older than MIS6 (Busschers et al., Reference Busschers, Kasse, Van Balen, Vandenberghe, Cohen, Weerst, Wallinga, Johns, Cleveringa and Bunnik2007; Peeters et al., Reference Peeters, Busschers, Stouthamer, Bosch, Van den Berg, Wallinga, Versendaal, Bunnik and Middelkoop2016). The two members are regionally separated by glacial sediments of the Peelo Formation (formed during the Elsterian glaciation); however, in this borehole the Peelo Formation is absent due to later erosion (Fig. 1C). The mollusc faunas, which are the subject of this paper, occur in both units between 19 and 38 m depth.
The Tynje Member interval consists of coarse-grained gravelly sands with molluscs (Fig. 2). The sandy deposits have a pink-brownish appearance and are rich in carbonate. The basal part of this unit (25–29 m) is particularly coarse-grained and rich in shells and shell fragments. The shells concentrate around the level of deepest erosion where the Peelo Formation was entirely removed by the river Rhine and the Tynje Member filled the incision (Fig. 1C). The mollusc fauna found in the Tynje Member is dominated by warm-temperate and temperate marine species, with mixed preservations styles. Five species occur, mostly in good preservation (fine surface detail preserved, little surface wear), that hitherto have been used as Eemian (MIS5e) indicator species in the North Sea Basin, viz. Bittium reticulatum, B. scabrum, B. jadertinum, Polititapes senescens and Acanthocardia paucicostata (Fig. 2; Van Leeuwen et al., Reference Van Leeuwen, Beets, Bosch, Burger, Cleveringa, van Harten, Herngreen, Kruk, Langereis, Meijer, Pouwer and de Wolf2000; Wesselingh et al., Reference Wesselingh, Rijken, van Nieulande and Pouwer2012; Busschers et al., Reference Busschers, Wesselingh, Kars, Versluijs-Helder, Wallinga, Bosch, Timmner, Nierop, Meijer, Bunnik and de Wolf2014; Meijer et al., Reference Meijer, Pouwer, Cleveringa, de Wolf, Busschers and Wesselingh2021). Some incompatible autecological indicators co-occur: B. cf. jadertinum and Nucula cf. nitidosa suggest polyhaline to euhaline conditions, Cerastoderma glaucum euryhaline conditions and Succinaidae indet. is an amphibious species common in freshwater lowland settings. Littorina saxatilis is an intertidal to shallow subtidal species, whereas Varicorbula gibba does not live above the storm wave base. Most likely this interval of the Tynje Member represents local fluvial reworking of marine deposits with faunas that for the most part lived in warm, clear water and shallow marine environments with seagrass and/or seaweed (Meijer et al., Reference Meijer, Pouwer, Cleveringa, de Wolf, Busschers and Wesselingh2021). The marine species found match those of the underlying 30–38 m interval of the Veenhuizen Member (see next section). In the overlying interval (19–25 m depth) abraded to very abraded fragments of some common marine bivalve species occur, most commonly C. edule in medium coarse sand with gravel. The coarse-grained deposits and severe wear indicate reworking in a fluvial setting.
The lower unit of the Urk Formation is referred to as the Veenhuizen Member, present at 29–59 m b.s. (Fig. 1; TNO-GSN, 2021). This unit is composed of fine- to medium-grained sands that locally contain clay pebbles (likely representing thin clay layers disturbed during drilling of the borehole), mica flakes and some glauconite. Sample 30–31 m depth is dominated by Mytilus fragments. In sample interval 33–34 m, a large fragment of a worn littoral Littorina littorea was found. Two slightly worn occurrences of Bittium and Mytilus were found in sample interval 37–38 m (Fig. 2a, f, g). However, the Bittium has almost perfect preservation of fine surface details and periostracum at the body whorl. The fine grained nature of the sediments at this depth, the presence of clay layers and marine molluscs suggest that at least part of Veenhuizen Member was deposited in a marine setting.
Age constraints
The mollusc fauna is located below MIS6 dated sediments of the Drente Formation (i.e. Busschers et al., Reference Busschers, Van Balen, Cohen, Kasse, Weerts, Wallinga and Bunnik2008; Busschers et al., Reference Busschers, Wesselingh, Kars, Versluijs-Helder, Wallinga, Bosch, Timmner, Nierop, Meijer, Bunnik and de Wolf2014; Kars et al., Reference Kars, Busschers and Wallinga2012) and early MIS6 dated sediments of the Drachten Formation (Kars et al., Reference Kars, Busschers and Wallinga2012; Busschers et al., Reference Busschers, Wesselingh, Kars, Versluijs-Helder, Wallinga, Bosch, Timmner, Nierop, Meijer, Bunnik and de Wolf2014). The faunal assemblage is therefore certainly older than MIS5e. Moreover, since we observe (1) a partial match between molluscs from the basal Tynje interval with those found in the underlying Veenhuizen Member, (2) high mollusc concentrations occurring right at the position of deepest erosion at the base of the Tynje Member and (3) no mollusc-bearing sources in the direct vicinity of the borehole occur, we consider it plausible that the molluscs from the interval 19–29 m b.s. of the Tynje Member were reworked from the older Veenhuizen Member.
The shell-bearing sediments of the Veenhuizen Member are stratigraphically positioned below the Peelo Formation (Figs. 1 and 2), which was deposited during MIS 10 (e.g. Zagwijn, Reference Zagwijn and Turner1996; Meijer & Cleveringa, Reference Meijer and Cleveringa2009) or MIS 12 (Gibbard, Reference Gibbard and Preece1995; Lauer & Weiss, Reference Lauer and Weiss2018; see Lee et al., Reference Lee, Busschers and Sejrup2012 for discussion). Consequently, the lusitanian molluscs present in the 29–38 m depth interval must be older than MIS10. Taking into account the interglacial nature of the molluscs, a MIS11 stage or older is therefore most likely. Additionally, the stratigraphic position of this section is similar to sediments of the Noordbergum Interglacial as defined by Zagwijn (Reference Zagwijn and Turner1996) about 25 km north of the borehole site. It implies these molluscs, including the lusitanian faunas in the basal 4 metres of the Tynje Member, also have an MIS11 or older age.
Implications
The mollusc faunas in the Quaternary intervals of the Luxwoude borehole are dominated by polyhaline, warm-temperate (‘lusitanian’ and ‘celtic’) and shallow marine species (cf. Cleveringa et al., Reference Cleveringa, Meijer, van Leeuwen, de Wolf, Pouwer, Lissenberg and Burger2000; Van Leeuwen et al., Reference Van Leeuwen, Beets, Bosch, Burger, Cleveringa, van Harten, Herngreen, Kruk, Langereis, Meijer, Pouwer and de Wolf2000; Meijer et al., Reference Meijer, Pouwer, Cleveringa, de Wolf, Busschers and Wesselingh2021). Five species hitherto used as indicators for the warm MIS5e (Eemian) intervals are now – for the first time – retrieved from Middle Pleistocene intervals in the southern North Sea Basin, viz. Bittium reticulatum, B. scabrum, B. cf. jadertinum, Acanthocardia paucicostatum and Polititapes senescens. The taxonomy of European Bittium species is not well-established and we follow the distinction proposed by Verduin (Reference Verduin1982); van der Linden & Wagner (Reference Van der Linden and Wagner1990) and Wesselingh et al. (Reference Wesselingh, Rijken, van Nieulande and Pouwer2012). Within the Luxwoude samples, only A. paucicostatum is a thermophile species, but within Eemian assemblages the aforementioned species typically are accompanied by other warm-temperate species (e.g., Meijer et al., Reference Meijer, Pouwer, Cleveringa, de Wolf, Busschers and Wesselingh2021). The low number of specimens in the limited sediment volume that was available implies that larger sediment samples almost certainly will provide further lusitanian species.
Middle Pleistocene interglacial marine mollusc faunas have previously been reported at several – generally isolated – localities in the southern North Sea Basin (Meijer, Reference Meijer2003; Meijer & Cleveringa, Reference Meijer and Cleveringa2009; Barlow et al., Reference Barlow, Long, Gehrels, Saher, Scaife, Davies, Penkman, Bridgland, Sparkes, Smart and Taylor2017; Meijer et al., Reference Meijer, Pouwer, Cleveringa, de Wolf, Busschers and Wesselingh2021). These mostly concern very small and poorly preserved faunas with the exception of the recently reported fauna found in borehole Hollum on Ameland, about 50 km north of Luxwoude, that was assigned a MIS7 age on the basis of amino acid racemisation analyses (Meijer et al., Reference Meijer, Pouwer, Cleveringa, de Wolf, Busschers and Wesselingh2021; age model cf. Meijer & Cleveringa, Reference Meijer and Cleveringa2009). The earlier reported faunas are considered to be of the Celtic type (similar as Holocene marine faunas) lacking any of the thermophilous species characteristic of lusitanian/Eemian assemblages. The Middle Pleistocene faunas reported in this study point to submarine conditions very similar to the Eemian (i.e. warmer than the present day) and warmer settings than those of MIS1 and MIS7 faunas reported by Meijer et al. (Reference Meijer, Pouwer, Cleveringa, de Wolf, Busschers and Wesselingh2021). Several lusitanian species are well capable of living in the southern North Sea, but these are unable to survive the cold waters of northern Scotland (Meijer & Preece, Reference Meijer and Preece1995). Their access into the North Sea Basin is therefore unlikely to have been through a northern route. Hence, these species are considered to be good indicators for a southern Channel connection into the North Sea, similar to the conditions existing today.
Even though the new finding shows that lusitanian Bittium-Polititapes dominated faunas cannot be considered a unique Eemian indicator assemblage, marine Middle Pleistocene intervals in the southern North Sea Basin are seldom encountered because of their occurrence at much greater depths only retrievable by boreholes. Conversely, MIS5e intervals are more accessible because of their shallow position, and their age can often be well-constrained through direct dating and their occurrence between distinct Saalian and Weichselian glacial and proglacial sediments. This implies that even though the assemblages are not uniquely Eemian, in the vast majority of cases they will be so, but we urge caution with the use of Bittium-dominated faunas as the sole indicator for MIS 5e.
Acknowledgements
Charles Barnard (Naturalis) assisted in sorting the samples. We thank the borehole crew of De Ruiter and Ate Oosterhof from VITENS N.V. who made the samples available. We are very much indebted to reviewers Richard Preece and Svend Funder for their suggestions and corrections.



 Open access
Open access