


Introduction
In any given landscape, protecting a priority species may have spill-over benefits for other species (Caro, Reference Caro2003). So-called umbrella species are characterized as requiring sufficiently large areas of habitat that investment in their protection will also benefit many other species (Noss, Reference Noss1990; Andelman & Fagan, Reference Andelman and Fagan2000). Top predators often meet this criterion (Sergio et al., Reference Sergio, Newton, Marchesi and Pedrini2006; Dalerum et al., Reference Dalerum, Somers, Kunkel and Cameron2008; Rozylowicz et al., Reference Rozylowicz, Popescu, Pătroescu and Chişamera2011), with many large carnivores additionally possessing charismatic qualities and wide public recognition that can attract disproportionate conservation investments (Sergio et al., Reference Sergio, Newton, Marchesi and Pedrini2006; Karanth & Chellam, Reference Karanth and Chellam2009). The flagship status of such carnivores can bring indirect benefits to other species that are neglected or overlooked, by highlighting common threats and emphasizing their mutual dependence. A fundamental step in identifying and quantifying potential benefits for other species is to demonstrate the spatial extent of co-occurrence in the area of interest (Andelman & Fagan, Reference Andelman and Fagan2000).
In the mountains of Central Asia the Endangered snow leopard Panthera uncia (Jackson et al., Reference Jackson, Mallon, McCarthy, Chundaway and Habib2008) has been highlighted as a potential umbrella species (Bhatnagar et al., Reference Bhatnagar, Mathur and McCarthy2002; Snow Leopard Network, 2014). The snow leopard has acquired an iconic status worldwide and is being promoted as a flagship species (Snow Leopard Working Secretariat, 2013). Snow leopards are known to have large home ranges and their geographical distribution overlaps with that of other carnivores, such as the grey wolf Canis lupus, Eurasian lynx Lynx lynx and red fox Vulpes vulpes (Fig. 1). There have been few quantitative investigations of the interactions of snow leopards with these other carnivore species (Jumabay-Uulu et al., Reference Jumabay-Uulu, Wegge, Mishra and Sharma2014; Li et al., Reference Li, Schaller, McCarthy, Wang, Jiagong and Cai2013), which all tend to be regarded negatively by local herding communities and are consequently vulnerable to retaliatory killing (Ale et al., Reference Ale, Yonzon and Thapa2007; Worthy & Foggin, Reference Worthy and Foggin2008; Alexander et al., Reference Alexander, Chen, Damerell, Youkui, Hughes, Shi and Riordan2015a).

Fig. 1 Distribution of the snow leopard Panthera uncia (dark grey shading), the Eurasian lynx Lynx lynx (light grey, a), grey wolf Canis lupus (light grey, b) and red fox Vulpes vulpes (light grey, c). Total overlap in the distribution of snow leopards and that of wolves, lynx and red foxes is 97, 100 and 100%, respectively (IUCN, 2015).
As a first step in assessing the potential impacts of snow leopard conservation on sympatric carnivores, we investigated the presence of other carnivores within snow leopard areas. We combined fine-scale camera trap data with wider landscape-scale data from sign surveys to determine the degree to which other carnivores use the same travel routes as snow leopards. We also examined temporal activity patterns and the consequences for potential interspecific interactions.
Methods
Fieldwork was conducted in Qilianshan National Nature Reserve, in the south-central part of Gansu Province, China. Camera trap data were collected from a total of 72 camera trap stations, each consisting of a single camera trap unit, within a 480 km2 area (Fig. 2). A pilot study conducted at this site in 2012 had confirmed the presence of snow leopards. Sixty camera trap stations were active during January–March 2013 (Alexander et al., Reference Alexander, Gopalaswamy, Shi and Riordan2015b). During April 2013–July 2014 we maintained 23 of these camera trap stations and added an additional 12 stations to maximize the number of capture events. As our target species was the snow leopard, the locations of camera trap stations were chosen based on the high density of snow leopard signs, and on the presence of natural pathways (such as ridges and valleys) that individual snow leopards were likely to use. We recorded capture histories for each carnivore species, also noting the time of capture. To avoid pseudoreplication, when the same species was captured more than once during a period of < 30 minutes only the first occasion was recorded as an independent capture event.

Fig. 2 (a) Location of Qilianshan National Nature Reserve, Gansu Province China. (b) Locations of camera traps and grid cells in which transect surveys were conducted in the Reserve.
The times of each independent event were partitioned into hourly intervals to establish 24-hour activity patterns for each species. Temporal interspecific interactions were investigated by estimating the kernel density of temporal overlap (Ridout & Linkie, Reference Ridout and Linkie2009). We compared the temporal activity patterns of snow leopards with those of the red fox and the Eurasian lynx by estimating the coefficient of overlap (Δ4 and Δ1), ranging from 0 (no overlap) to 1 (complete overlap). Confidence intervals at 95% were obtained as percentile intervals from 500 bootstrap samples (Linkie & Ridout, Reference Linkie and Ridout2011). Analyses were performed using the package overlap v. 0.2.4 (Meredith & Ridout, Reference Meredith and Ridout2014) in R v. 3.1.3 (R Development Core Team, 2014).
During January–March 2014 we conducted sign surveys along routes that snow leopards were expected to use, within 49 grid cells of a larger area of the Reserve (Fig. 2). Mammalian carnivore sign was recorded in 100 m segments along transects, and categorized according to species. We recorded only tracks that were easily recognizable, identified unambiguously and considered to be recent (< 1 month old), with sharply defined edges and shapes. Multiple signs of the same species in the same 100 m segment were not recorded separately. Signs of snow leopard, lynx, grey wolf and red fox were differentiated on the basis of size, shape and context-specific information; for example snow leopard tracks tend to be larger than those of lynx (Mallon, Reference Mallon1984), although those of a subadult snow leopard are of a similar size to those of an adult lynx (Jackson & Hunter, Reference Jackson and Hunter1996). We therefore ascribed such smaller tracks to lynx unless they were accompanied by tracks of an adult snow leopard, given that young snow leopards stay with their mothers for an estimated 18–22 months after birth (Jackson, Reference Jackson1996). We did not record canid tracks if signs of human presence were also found in the same area, to avoid recording tracks of domestic dogs.
Results
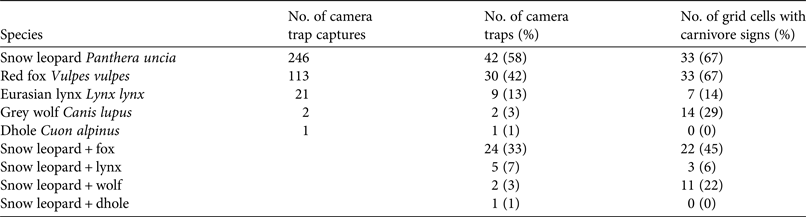
Cameras were active for a collective total of 8,498 trap days. We obtained 246 captures of snow leopard, 113 of red fox, 21 of lynx, two of grey wolf and one of dhole Cuon alpinus. We captured snow leopards at 42 of the 72 (58%) camera trap stations. Red foxes were captured at 30 (42%), lynx at nine (13%), and grey wolves at two (3%) of the stations (Table 1). Red foxes tended to be captured at camera trap stations that also captured snow leopards: 24 camera traps captured both species and accounted for the majority (79%) of fox captures. Lynx also tended to be captured at the same locations as snow leopards, with five camera traps capturing both species and accounting for the majority (67%) of lynx captures.
Table 1 Results of camera trap and transect surveys of carnivores in Qilianshan National Nature Reserve, China, with species, no. of camera-trap captures, no. of camera traps that captured the species, and no. of grid cells in which signs of the species were recorded on transects.
Snow leopards were most commonly detected in the evening and early night (17.00–21.00), with a peak around sunset (19.00–20.00; Fig. 3). Detections of snow leopards were also common during 01.00–07.00. Captures of red foxes were most common early in the night (21.00–midnight) and in the early hours of the morning (03.00–06.00), peaking during 05.00–06.00. The lynx detection pattern was less obvious because of the small sample size but suggests continuous detections, with a somewhat decreased frequency during the day. The two detections of wolves were at 20.30 and 23.43. Kernel density estimates suggested a high degree of temporal overlap between the snow leopard and the red fox (Fig. 3), as indicated by a high overlap coefficient estimate (Δ4 = 0.72, 95% CI 0.61–0.78). There was also a high degree of temporal overlap between the snow leopard and the lynx (Δ1 = 0.85, 95% CI 0.80–0.95) but this should be interpreted with caution given the small number of lynx captures (n = 21).
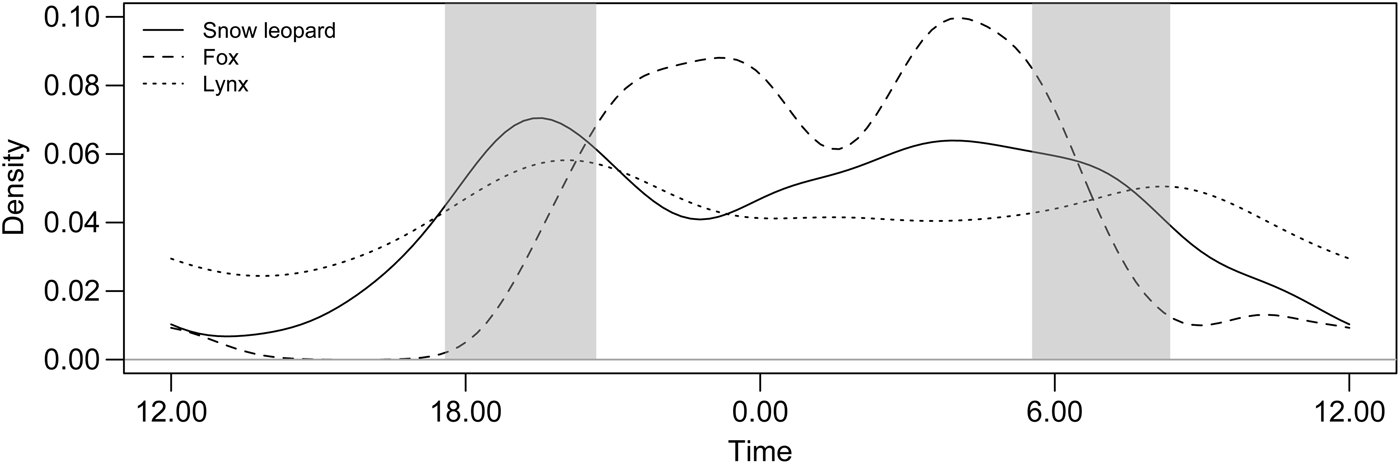
Fig. 3 Activity patterns of the snow leopard, red fox and Eurasian lynx, based on camera trap surveys in Qilianshan National Nature Reserve (snow leopard, n = 246; red fox, n = 113; Eurasian lynx, n = 21), presented as kernel density estimates. The light grey bands indicate sunrise (05.52–08.37) and sunset (17.56–20.48).
At the landscape scale a total of 244 km of transects were surveyed within 49 grid cells covering 784 km2 (Table 1). Snow leopard signs were detected most frequently, occurring in 33 grid cells and on 34% of transects. Signs of red fox were also widespread, detected in 33 grid cells and on 28% of transects. Grey wolf and lynx were detected in 14 (naïve occupancy = 29%) and seven (naïve occupancy = 14%) grid cells, respectively. Concurrent signs of both snow leopard and red fox were detected in 22 grid cells (Table 1). Signs of both snow leopard and lynx were detected in three grid cells, and of both snow leopard and wolf in 11 grid cells.
Discussion
The results of our survey at various scales confirm that other carnivores occur in areas of snow leopard occurrence, albeit with low detection rates for the lynx, wolf and dhole. Even in the smaller scale camera-trap survey all five carnivores (snow leopard, lynx, wolf, red fox and dhole) were observed. These simple relationships suggest there is some potential for the protection of snow leopard habitat to benefit other species. Camera traps recorded snow leopards and lynx using the same travel route at five locations. If, as intimated by these findings, snow leopards and other carnivores share the same travel routes, there is an opportunity to consider protective measures at the local scale that would simultaneously benefit multiple species; for example, preventing the use of poaching traps along such routes or minimizing human disturbances (such as fences and roads) would have multiplier effects on other species. This would be particularly relevant in Qilianshan National Nature Reserve and other areas in China (Xu et al., Reference Xu, Jiang, Li, Guo, Da and Cui2008) where development projects (including road building, mining and hydroelectric dams) are underway. The use of predator-proof corrals (which protect livestock from predation by all carnivores) and better husbandry practices (including control of herding dogs, which often come into conflict with carnivores) should also be encouraged (Aryal et al., Reference Aryal, Brunton, Ji, Barraclough and Raubenheimer2014; Alexander et al., Reference Alexander, Chen, Damerell, Youkui, Hughes, Shi and Riordan2015a).
It is important to recognize that snow leopard conservation efforts could potentially increase the population numbers of carnivores that have a negative impact on local livelihoods. This could lead to exacerbation of livestock depredation and further human–wildlife conflicts. In the study area lynx and wolves are seen as the main culprits of livestock depredation, and their increased numbers could have negative consequences for snow leopards (Alexander et al., Reference Alexander, Chen, Damerell, Youkui, Hughes, Shi and Riordan2015a). Umbrella effects of conservation efforts must therefore be taken into consideration not only in terms of potential benefits for other threatened species but also because of the complex interactions inherent in human–wildlife conflict involving an assemblage of carnivore species.
This study is exploratory and further assessments are required to evaluate the ecological significance of sympatric carnivore co-occurrence across these mountain landscapes. Our survey design, which targeted snow leopards, may have led to low capture rates of wolves and lynx, thus limiting inferences on dynamics and interactions. These low rates may also be attributable to lower densities of these species, or differential space use between lynx, wolves and snow leopards. Further studies on interspecific interactions within this carnivore assemblage should attempt to broaden the range of sampling sites beyond those preferred by snow leopards. A non-targeted approach, however, carries a risk of low detection probabilities for all species. We therefore recommend combining information from surveys that target each species of interest. Sign surveys can be carried out over large areas more readily then camera-trap surveys but must be conducted carefully to minimize misidentification of species.
We provide evidence that a number of carnivores share the snow leopard's habitat at multiple scales. This raises the possibility that efforts to conserve the snow leopard could have both positive and unintended negative spill-over effects. Although simple measures could leverage broad-based synergistic impacts among species, complex interactions arising from human–wildlife conflicts could influence final outcomes.
Acknowledgements
We acknowledge support from the Second National Survey of Terrestrial Wildlife in China, State Forestry Administration of China, and National Natural Science Foundation of China (Grant No. 31470567). PR thanks the Starr Foundation for support during the undertaking of this work. We thank Qilianshan National Nature Reserve, and Wang Youkui, A. Cheng and Joelene Hughes for their help and support during fieldwork in Gansu Province, China.
Biographical sketches
Justine Shanti Alexander is studying the conservation of snow leopards in high-elevation areas of China. Jeremy Cusack is interested in optimizing camera-trap surveys targeted at community-level inferences and processes. Chen Pengju is interested in the conservation of snow leopards and wild ungulates in China. Shi Kun translates academic research into conservation through training local wildlife managers, building a collaborative network, and applied research. Philip Riordan's work in the UK, China and Africa focuses on top predators, their ecosystems and their interactions with people.




