


Introduction
The Lake Tumba swamps (c. 78,972 km2) of the Democratic Republic of Congo (DRC) are among the 12 priority conservation landscapes funded by the Central African Regional Programme for the Environment of the US Agency for International Development through the International Congo Basin Forest Partnership. The site was selected as a priority conservation site in 2000 because it has biomes specific to this area (Toham et al., Reference Toham, Adeleke, Burgess, Carroll, D'Amico, Dinerstein, Olson and Some2003; Burgess et al., Reference Burgess, D'Amico, Underwood, Dinerstein, Olson, Itoua, Shipper, Ricketts and Newman2004; Thieme et al., Reference Thieme, Abell, Stiassny, Skelton, Lehner, Teugels, Dinerstein, Toham, Burgess and Olson2005; Toham et al., Reference Toham, D'Amico, Olson, Blom, Trowbridge, Burgess, Thieme, Abell, Carroll, Gartlan, Langrand, Mussavu, O'Hara and Strand2006), which is one of the most extensive swampy forests in Africa. However, despite the conservation importance of the Lake Tumba swamps, known to support at least one great ape, the Endangered bonobo Pan paniscus (Kempf & Wilson, Reference Kempf and Wilson1997; IUCN, 2006), and a range of other mammalian, ornithological and herpetological diversity, the area has never been properly surveyed. The little that is known concerning the presence of the bonobo is confined to the region near Lake Tumba, particularly near Botuali (Horn, Reference Horn1976, Reference Horn1980; Mwanza et al., Reference Mwanza, Mulavwa, Ihomi and Kumugo2003; Fig. 1).
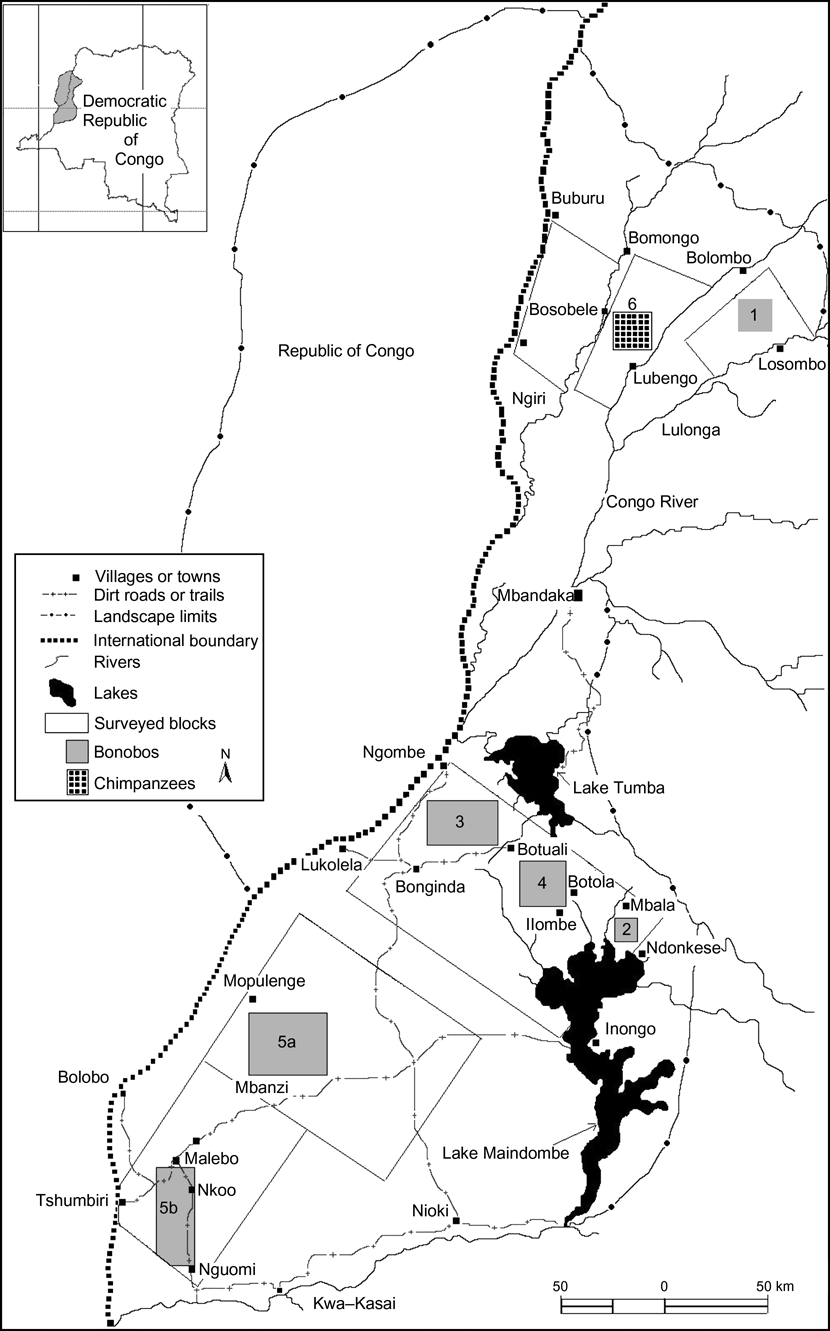
Fig. 1 Habitat blocks surveyed for great apes in the Lake Tumba area, western Democratic Republic of Congo, and the locations of the five populations of bonobo (1-5) and one population of chimpanzees (6). The limits of the Lake Tumba landscape in the main figure are indicated in the inset.
The first phase of the implementation of this project was therefore to document species distributions, with a particular emphasis on bonobo, diurnal monkeys, and large herbivores such as forest buffalo Syncerus caffer nanus. This information is a prerequisite for planning effective conservation activity (Primack, Reference Primack2000; Kormos et al., Reference Kormos, Bakkar, Bonnéhin, Hanson-Alp, Kormos, Boesch, Bakkar and Butynski2003). Furthermore, there is a need to provide up to date knowledge of great ape populations in the region because previous studies have indicated that the swampy forests of the DRC are the least represented habitat type within the country's protected area network (Inogwabini et al., Reference Inogwabini, Omari and Mbayma2005a,b).
The objectives of the survey were to (1) document the distribution of large mammal species of conservation concern, and (2) estimate species abundances where possible, and in relative terms where sample sizes did not permit direct estimation of density. Here we report only the information gathered on bonobo and chimpanzee Pan troglodytes. In so doing we also fill the gap identified in the prioritization of great ape populations carried out by the Great Ape Survival Project, which indicated that the knowledge of the distribution of great apes in DRC was imprecise and limited (Jackson, Reference Jackson2005).
Study area
About 60% of the Lake Tumba landscape (Fig. 1) is inundated and seasonally flooded forest. Depending on the season, it is home to a large mammal assemblage that includes the bonobo, chimpanzee, Angolan pied colobus Colobus angolensis, Allen's swamp monkey Allenopithecus nigriviridis, black mangabey Lophocebus aterrimus, red colobus Piliocolobus tholonii, red-tailed monkey Cercopithecus ascanius, forest elephant Loxodonta africana, buffalo and leopard Panthera pardus (Marcot & Alexander, Reference Marcot and Alexander2004; Inogwabini, Reference Inogwabini2005). The diverse swampy biotopes have a rich diversity of fish and freshwater-dependent species such as sitatunga Tragelaphus spekei, water chevrotain Hyemoscus aquaticus, slender-snout crocodile Crocodylus cataphractus, Nile crocodile Crocodylus niloticus and hippopotamus Hippopotamus amphibius (Toham et al., Reference Toham, D'Amico, Olson, Blom, Trowbridge, Burgess, Thieme, Abell, Carroll, Gartlan, Langrand, Mussavu, O'Hara and Strand2006). Threats to the biodiversity include increases in the local human population and logging for the wenge Mellitia laurenti (Marcot & Alexander, Reference Marcot and Alexander2004). Considerable unmonitored fishing occurs and bushmeat hunting is common (Authors, pers. obs.). Poaching is insidious because of the availability of weapons and ammunitions and the location of the area at the junction of major river routes (Marcot & Alexander, Reference Marcot and Alexander2004; Inogwabini, Reference Inogwabini2005). An additional problem is the general human poverty in the area, which adds excessive strain on natural resources through unplanned and unsustainable subsistence agricultural practices (Marcot & Alexander, Reference Marcot and Alexander2004; Inogwabini, Reference Inogwabini2005; Alexander & Lerum, Reference Alexander and Lerum2006).
Methods
We surveyed the area between Lake Tumba in the north and the Kwa-Kasai River in the south, bordered to the east by Lake Maindombe and to the west by the Congo River (Fig. 1). We chose this area because it is traversed by logging concessions, making knowledge of the large mammals a priority. We also surveyed areas further north, between Buburu and Bosobele, Bosobele and Lubengo, and Bolombo and Losombo (Fig. 1), where the habitats are still relatively intact.
We combined field reconnaissance (White & Edwards, Reference White, Edwards, White and Edwards2000) and line transect methods (Buckland et al., Reference Buckland, Anderson, Burnham and Laake1993) to collect presence/absence data (Manel et al., Reference Manel, Williams and Ormerod2001; Ibáñez et al., Reference Ibáñez, Caniego and Garciá-Álvarez2005; Jepsen et al., Reference Jepsen, Baveco, Topping, Verboom and Vos2005) and estimate abundance, respectively, during May-December 2005. Following the collection of preliminary data from two locations where 14 1-km transects were cut, a total length of 86 km of transects were surveyed, as determined from CV(Đ) = (b/L (no/lo))1/2, where Đ is density, b = 3, as suggested by Buckland et al. (Reference Buckland, Anderson, Burnham and Laake1993) for surveys in forest, L = required sampling distance in km, no = observations and lo = effort (km) of the preliminary study, and using a coefficient of variation CV(Đ) of 10% (Buckland et al., Reference Buckland, Anderson, Burnham and Laake1993). During the surveys we collected data on nest sites, knuckle prints, feeding remains, calls, and made direct observations. Only nest sites were used to estimate population sizes. Geographical locations were determined with a global positioning system. We also made a total of 324 km of reconnaissance trips, on both sides of the Congo River. Apart from the logistical support team, which numbered 8-10 people depending on the length of the mission, the survey teams comprised a team leader, assistant team leader, compass bearer, and transect cutter. The team leader looked for ape nests, noted changes in habitat types, and recorded all data. The assistant team leader searched for signs on the ground such as dung, pellets and evidence of humans. The compass bearer directed the transect cutter or otherwise indicated the orientation of the reconnaissance.
To estimate bonobo density we first calculated D using the software Distance v. 4.1 (Thomas et al., Reference Thomas, Buckland, Burnham, Anderson, Laake, Borchers and Strindberg2002). Distance assumes that all objects at the centre line of the transect are detected (Buckland et al., Reference Buckland, Anderson, Burnham and Laake1993) and estimates D by n / (2*L*ESW), where n = number of sightings, L = total transect length, and ESW = effective strip width of the transect. We then used the formula of McClanahan (Reference McClanahan1986), as applied under different circumstances (Barnes et al., Reference Barnes, Blom, Alers and Barnes1995; Hall et al., Reference Hall, Inogwabini, Williamson, Omari, Sikubwabo and White1997), E = D/(r*d), where E = estimated density, D = density of signs, r = daily rate of nest production ( = 1), and d = bonobo nest decay rate (90 days; Reinartz et al., Reference Reinartz, Inogwabini, Mafuta and Lisalama2006). To avoid unwarranted extrapolation of density estimates over large areas we did not incorporate areas that were unlikely to be suitable for bonobos (Inogwabini & Omari, Reference Inogwabini and Omari2005; Reinartz et al., Reference Reinartz, Inogwabini, Mafuta and Lisalama2006). We did this by using the classification tool of the geographical information system ArcView v. 3.2 (ESRI, Redlands, USA) to identify open habitats (villages, savannah and large water bodies), permanent swampy forest, and young regenerating forest. At the edges of Lake Tumba we excluded the 2,874 km2 that is seasonally flooded. For the Malebo area, which comprises 1,044 km2 of mixed mature forests, 127 km2 of degraded forests and 751 km2 of savannah, we excluded savannah and degraded habitat in calculating the southernmost population. Because of small sample sizes from the edges of Lake Tumba, data from the Ngombe-Botuali and Botuali-Ilombe areas (Fig. 1) were pooled to estimate densities because habitats in these areas are similar. For the area of Mopulenge-Mbanzi where we did not have sufficient transect replicates to provide abundance estimates, and in the areas between Buburu and Bosobele, Bomongo and Lubengo, and Bolombo and Losombo (Fig. 1) where we had only made reconnaissance trips, we calculated encounter rates, which provide a relative indication of abundance (Strayer, Reference Strayer1999; Sutherland, Reference Sutherland1999; Walsh & White, Reference Walsh and White1999; Remis, Reference Remis2000; Sutherland, Reference Sutherland2000). Locations were plotted using ArcView, which was also used to differentiate forest types using Landsat satellite images provided by the Observatoire Satellital des Forêts d'Afrique Centrale, working in collaboration with the University of Maryland, USA and the Central African Regional Programme for the Environment.
Results
We located bonobo groups in five areas: (1) Bolombo-Losombo, (2) Mbala-Donkese, (3) Ngombe-Botuali, (4) Botuali-Ilombe, and (5) Mompulenge–Mbanzi-Malebo–Nguomi (Fig. 1). There were two sub-groups in the latter area, separated by 50 km of unbroken gallery forest (5a: Mopulenge-Mbanzi, 5b: Malebo-Nguomi) that may provide a link between them. One group of chimpanzees was found in the area between Bosobele and Lubengo (6). Estimates of bonobo densities and populations in sites 1, 3, 4 and 5b are given in Table 1, and nest site encounter rates of bonobos in sites 1 and 5a and chimpanzees in site 6 are given in Table 2.
Table 1 Bonobo densities and population estimates in four of the surveyed areas (numbers in Fig. 1), with size of the area surveyed and area of suitable bonobo habitat.

Table 2 Encounter rates for bonobos and chimpanzees in three of the surveyed areas (numbers in Fig. 1), with size of the area surveyed and area of suitable habitat.

Discussion
The first nationwide reviews of the distribution of bonobo in the Democratic Republic of Congo, using a combination of questionnaires, rapid assessment techniques and literature reviews (Fenart & Deblock, Reference Fenart and Deblock1973; Kano, Reference Kano1984; Thompson-Handler et al., Reference Thompson–Handler, Malenky and Reinartz1995), indicated that bonobo were present in 38 locations. However, only a few of these populations were confirmed by field surveys, including the 15 populations in Salonga National Park (Van Krunkelsven et al., Reference Van Krunkelsven, Inogwabini and Draulans2000; Inogwabini & Omari, Reference Inogwabini and Omari2005; Reinartz et al., Reference Reinartz, Inogwabini, Mafuta and Lisalama2006) and the populations in Wamba (Kano & Mulavwa, Reference Kano, Mulavwa and Susman1984) and Lomako (Malenky & Stiles, Reference Malenky and Stiles1991; Malenky & Wrangham, Reference Malenky and Wrangham1994; Dupain et al., Reference Dupain, Van Krunkelsven, Van Elsacker and Vereheyen2000). In the area between Lake Tumba and Lake Maindombe (Fenart & Deblock, Reference Fenart and Deblock1973; Horn, Reference Horn1976, Reference Horn1980; Mwanza et al., Reference Mwanza, Mulavwa, Ihomi and Kumugo2003) available information on bonobo remained fragmentary and imprecise. Only the Botuali-Ilombe population (4, Fig. 1) had been previously described (Horn, Reference Horn1976, Reference Horn1980; Mwanza et al., Reference Mwanza, Mulavwa, Ihomi and Kumugo2003). Populations in the areas of Mompulenge–Mbanzi-Malebo–Nguomi (5, Fig. 1) were confirmed by our surveys. The popualtions we located in the Ngombe-Botuali (3, Fig. 1) and Botuali-Ilombe areas (4, Fig. 1) lie in the area of the Botanankasa-Kunungu population described by Fenart & Deblock (Reference Fenart and Deblock1973), and the Mbala-Donkese population (2) lies in the area of the Lukolela and Tumba populations sketchily mapped by Kano (Reference Kano1984) and Horn (Reference Horn1976). The Bolombo-Losombo group (1, Fig. 1) has never been previously described.
The general pattern of the bonobo's distribution in the Lake Tumba landscape is similar to the patchy distribution of the species elsewhere (Kano, Reference Kano1984; Alers et al., Reference Alers, Blom, Sikubwabo, Masunda and Barnes1992; Inogwabini & Omari, Reference Inogwabini and Omari2005; Kortlandt, Reference Kortlandt1995). This pattern had been attributed to a combination of ecological, historical and evolutionary factors such as major flooding, epidemics, hunting pressure, forest exploitation, food availability, topography, and forest and land use history (Badrian & Malenky, Reference Badrian, Malenky and Susman1984; Kano & Mulavwa, Reference Kano, Mulavwa and Susman1984; Sabater Pi & Vea, Reference Sabater Pi and Vea1990; Malenky & Stiles, Reference Malenky and Stiles1991; Malenky & Wrangham, Reference Malenky and Wrangham1994; Kortlandt, Reference Kortlandt1995; Inogwabini & Omari, Reference Inogwabini and Omari2005). Human history in the area may also have influenced bonobo distribution patterns. Interviews with local communities (B.-I. Inogwabini et al., unpubl. data) indicate that the Bateke people inhabiting the Malebo area believe that bonobos are their ancestors, and therefore a traditional taboo prevents the Bateke from killing bonobo
While the lower ranges of estimates of bonobo density (0.24–1.8 bonobos km-2) in the area south of Lake Tumba are similar to reports from other areas (Sabater Pi & Vea, Reference Sabater Pi and Vea1990; Dupain et al., Reference Dupain, Van Krunkelsven, Van Elsacker and Vereheyen2000; Van Krunkelsven et al., Reference Van Krunkelsven, Inogwabini and Draulans2000; Van Krunkelsven, Reference Van Krunkelsven2001; Reinartz et al., Reference Reinartz, Inogwabini, Mafuta and Lisalama2006) and the encounter rate of bonobo in the Bolombo-Losombo area is lower than that in Salonga National Park (Inogwabini & Omari, Reference Inogwabini and Omari2005; Reinartz et al., Reference Reinartz, Inogwabini, Mafuta and Lisalama2006), the upper limit of the density estimate in the Malebo-Nguomi area (3.4 individuals km-2) is probably the highest estimate ever reported. The forest-savannah mosaic of this area may provide year-round fruit sources, and thus support larger groups, with bonobos falling back on savannah fruits when forest resources are scarce. It has been estimated that there are only c. 20,000 bonobos in the wild (Caldecott & Miles, Reference Caldecott and Miles2005) and the mean estimate of 2,297 in the the Malebo-Nguomi area (Table 1) therefore represents c. 12% of the known wild population, making this site particularly important for the conservation of the species. We were not able to estimate density in the Mopulenge-Mbanzi area but this population is also located in a forest-savannah mosaic habitat and the high encounter rate (1.2 individuals per km) may indicate that the area has a similarly high bonobo density.
The population of chimpanzees that we discovered between Bosobele and Lubengo is isolated from other known chimpanzee populations and inhabits a swampy habitat with dominant vegetation consisting of Pandanus sp. and Raffia sese that is markedly different from the described habitat type of this species. The encounter rate of chimpanzee signs (0.019 nest sites km-1) was lower than in either the Kahuzi-Biega National Park in eastern DRC (Inogwabini et al., Reference Inogwabini, Hall, Vedder, Curran, Yamagiwa and Basabose2000) or Luki in western DRC (Lomboto, Reference Lomboto2007). Chimpanzees are the least documented great apes in DRC (Hart & Hall, Reference Hart and Hall1996; Hall et al., Reference Hall, White, Inogwabini, Omari, Morland, Williamson, Walsh, Sikubwabo, Saltonstall, Dumbo, Kiswele, Vedder and Freeman1998; Omari et al., Reference Omari, Hart, Butynski, Birhashirwa, Upoki, M'keyo, Bashonga and Bagurubumwe1999; De Merode et al., Reference De Merode, Hillman-Smith, Nicholas, Ndey and Likango2000; Inogwabini et al., Reference Inogwabini, Hall, Vedder, Curran, Yamagiwa and Basabose2000), with precise information available only for populations in Ituri (Hart & Hall, Reference Hart and Hall1996), the Kahuzi-Biega National Park (Inogwabini et al., Reference Inogwabini, Hall, Vedder, Curran, Yamagiwa and Basabose2000), and the Itombwe forest (Omari et al., Reference Omari, Hart, Butynski, Birhashirwa, Upoki, M'keyo, Bashonga and Bagurubumwe1999). The conservation status of chimpanzee in DRC is similar to that of the species across its range, with populations having decreased in the last 5 decades (Butynski, Reference Butynski, Kormos, Boesch, Bakkar and Butynski2003; Kormos et al., Reference Kormos, Bakkar, Bonnéhin, Hanson-Alp, Kormos, Boesch, Bakkar and Butynski2003; Kormos & Boesch, Reference Kormos and Boesch2003). Therefore, despite an apparently low abundance as determined from the encounter rate, the chimpanzee population of the Bosobele-Lubengo area warrants a more detailed survey.
The discovery of both bonobo and chimpanzee in areas where they had not been previously described is good news for the conservation of these species. The bonobo were located in an area that extends the known range of the species further west. The high density estimates in a habitat type that would appear to be marginal suggests that extrapolations of abundance of this species based on habitat modelling (Kortlandt, Reference Kortlandt1995) need to be revised, and that only field surveys can provide realistic estimates. Furthermore, the discovery of this population in a savannah-forest mosaic habitat necessitates further ecological and ethological studies as the population may have adapted behaviours that have not been previously documented.
Acknowledgements
This publication was made possible through financial support provided by the US Agency for International Development through the Central African Regional Environmental Program–Congo Basin Forest Partnership. The authors’ views expressed in this publication do not necessarily reflect the views of the United States Agency for International Development or the United States Government. We would like to thank the DRC Government for permission to work in the Lake Tumba landscape. We thank the WWF National and Central African Regional coordination for unfailing support to the Lake Tumba project. The research was designed at the University of Kent under the supervision of Professor Nigel Leader-Williams. Constructive comments from two anonymous reviewers helped improve the paper.
Biographical sketches
Bila-Isia Inogwabini's work has included surveys of Grauer's gorilla, eastern chimpanzees and forest elephant in the eastern Democratic Republic of Congo (then Zaire). He commenced his research on bonobos in 1997, and between 2002 and 2004 contributed to the region-wide elephant survey initiated by CITES under its programme for the monitoring of illegal killing of elephants (MIKE). He now coordinates the WWF project in the Lake Tumba landscape. Bewa Matungila, Longwango Mbende and Mbenzo Abokome were team leaders for the region-wide elephant survey initiated by CITES under its MIKE programme. They joined the WWF's Lake Tumba Project in 2005 where they are in charge of the monitoring of large mammals. Tshimanga wa Tshimanga joined WWF's Lake Tumba Project in 2005 as the project's GIS officer.



