


Introduction
Establishing the population status and distribution of threatened species is crucial for informing their management and conservation, especially when populations are isolated and fragmented. The Arabian leopard Panthera pardus nimr previously roamed across the Arabian Peninsula but now < 200 individuals are believed to persist in fragmented populations in the wild (Islam et al., Reference Islam, Volmer, al Boug, as Shehri and Gavashelishvili2020). Panthera pardus is categorized as Vulnerable on the IUCN Red List (Stein et al., Reference Stein, Athreya, Gerngross, Balme, Henschel and Karanth2020), and P. pardus nimr as Critically Endangered (Mallon & Budd, Reference Mallon, D.P. and Budd, K.2011).
The Arabian leopard is the smallest of the eight subspecies of leopard, typically weighing 18–34 kg, approximately half the weight of an African leopard Panthera pardus pardus (Spalton & Al Hikmani, Reference Spalton and Al Hikmani2006). It is adapted to desert habitats and typically has paler pelage than other leopard species, and primarily nocturnal behaviour patterns (Spalton et al., Reference Spalton, Al Hikmani, Willis and Said2006b). The species’ historical range extended from the western mountains of Saudi Arabia to Yemen and Oman, with populations in Israel, Jordan, the United Arab Emirates and the Egyptian Sinai Peninsula (Perez et al., Reference Perez, Geffen and Mokady2006; Spalton & Al Hikmani, Reference Spalton and Al Hikmani2006; Spalton et al., Reference Spalton, Al Hikmani, Jahdhami, Ibrahim, Said and Willis2006a; Islam et al., Reference Islam, Boug, Shehri and Jackson2011; Mallon & Budd, Reference Mallon, D.P. and Budd, K.2011; Soultan et al., Reference Soultan, Attum, Hamada, Hatab, Ahmed and Eisa2017). Today, populations are only known to occur in Oman and Yemen; the southern mountains of Oman are a stronghold for the subspecies, and although populations persist in Yemen they are believed to be fragmented and isolated (Al Jumaily et al., Reference Al Jumaily, Mallon, Naher and Thowabeh2006; Spalton & Al Hikmani, Reference Spalton and Al Hikmani2006; Spalton & Al Hikmani, Reference Spalton and Al Hikmani2014; Farhadinia et al., Reference Farhadinia, Rostro-García, Feng, Kamler, Spalton and Shevtsova2020).
A large proportion of the historical range of the Arabian leopard fell within Saudi Arabia (Judas et al., Reference Judas, Paillat, Khoja and Boug2006; Islam et al., Reference Islam, Boug, Judas and As-Shehri2018), where it inhabited the arid mountainous regions and maintained substantial populations in the south-western mountains (Gasperetti et al., Reference Gasperetti, Harrison and Büttiker1985; Harrison & Bates, Reference Harrison and Bates1991; Judas et al., Reference Judas, Paillat, Khoja and Boug2006; Islam et al., Reference Islam, Boug, Judas and As-Shehri2018). The species’ range is, however, estimated to have declined by > 98%, and there have been no confirmed sightings of Arabian leopards in the country since 2014 (Jacobson et al., Reference Jacobson, Gerngross, Lemeris, Schoonover, Anco and Breitenmoser-Würsten2016; Islam et al., Reference Islam, Boug, Judas and As-Shehri2018). Sixty-five leopard sightings were reported during 1998–2001 but all were unsubstantiated, and field surveys to detect leopards during 2010–2017 failed to do so (Al-Johany, Reference Al-Johany2007; Islam et al., Reference Islam, Boug, Judas and As-Shehri2018). It is of critical importance to establish whether sustainable populations of Arabian leopards persist in Saudi Arabia and to protect and conserve any remaining individuals.
The decline of the leopard in Saudi Arabia can be attributed to environmental and anthropogenic factors that include direct and indirect threats (Islam et al., Reference Islam, Boug, Judas and As-Shehri2018). Islam et al. (Reference Islam, Boug, Judas and As-Shehri2018) conducted on-site interviews, observations and a literature review and found 52 cases of leopards that were hunted, trapped or illegally killed from 1947 to 2014, when the last known leopard was recorded in Saudi Arabia (36 of these cases were substantiated, J.A. Spalton, unpubl. data). Because of the sensitive nature of leopard hunting, this is presumed to be the minimum number during this period as killings were rarely documented, especially in earlier years. Direct threats facing any remaining individuals include poisoning, the illegal sale of furs and body parts, and the wildlife trade (Judas et al., Reference Judas, Paillat, Khoja and Boug2006, Islam et al., Reference Islam, Boug, As-Shehri and Al Jaid2015, Reference Islam, Boug, Judas and As-Shehri2018). Indirect threats occur from increasing urbanization, road construction, mining and gravel extraction, which cause habitat degradation and fragmentation (Judas et al., Reference Judas, Paillat, Khoja and Boug2006; Islam et al., Reference Islam, Boug and As-Shehri2017, Reference Islam, Boug, Judas and As-Shehri2018; Farhadinia et al., Reference Farhadinia, Rostro-García, Feng, Kamler, Spalton and Shevtsova2020). Overgrazing from expanding livestock herds and poaching for meat also reduce the wild prey of Arabian leopards and contribute further to the decline of the species (Judas et al., Reference Judas, Paillat, Khoja and Boug2006; Islam et al., Reference Islam, Boug, Judas and As-Shehri2018, Reference Islam, Volmer, al Boug, as Shehri and Gavashelishvili2020). The Arabian gazelle Gazella arabica, Nubian ibex Capra nubiana, Cape hare Lepus capensis and rock hyrax Procavia capensis comprise the majority of the Arabian leopard's diet in Oman (Muir-Wright, Reference Muir-Wright1999), and their scarcity in Saudi Arabia has probably contributed to the leopard's decline as well as causing leopards to predate on livestock, resulting in retaliatory and pre-emptive killings by herders (Islam et al., Reference Islam, Boug, Judas and As-Shehri2018). Comprehensive surveys of the status of the Arabian leopard and its prey species are crucial if this subspecies is to be conserved.
Finding rare and elusive species in landscapes spanning thousands of square kilometres is challenging, but technologies such as camera traps and DNA analysis can provide information on presence, population size and habitat selection (Pimm et al., Reference Pimm, Alibhai, Bergl, Dehgan, Giri and Jewell2015; Pitman et al., Reference Pitman, Fattebert, Williams, Williams, Hill and Hunter2017; Al Hikmani, Reference Al Hikmani2019; Chetri et al., Reference Chetri, Odden, Sharma, Flagstad and Wegge2019; Ghoddousi et al., Reference Ghoddousi, Bleyhl, Sichau, Ashayeri, Moghadas and Sepahvand2020). Combining these methods, along with remote sensing and satellite imagery, can improve the detection of rare species (Long et al., Reference Long, Donovan, MacKay, Zielinski and Buzas2010; Pimm et al., Reference Pimm, Alibhai, Bergl, Dehgan, Giri and Jewell2015). Methods that target different scales can be complementary (e.g. broad-scale questionnaire surveys across historical range combined with camera trapping at key sites; Zeller et al., Reference Zeller, Nijhawan, Salom-Pérez, Potosme and Hines2011; Pitman et al., Reference Pitman, Fattebert, Williams, Williams, Hill and Hunter2017). The use of individual monitoring tools, such as tracking collars, is precluded here because of the scarcity of Arabian leopards in this landscape, so we chose to employ remote and non-invasive methods.
We designed this study to determine the current status of the Arabian leopard across its historical range in Saudi Arabia. We utilized habitat modelling (Dunford et al., Reference Dunford, Martins, Mann, Spalton, Al Hikmani and Robinson2022) and three field survey methods at two scales: intensive camera-trap surveys combined with opportunistic scat collection at sites that had high potential to support extant leopard populations, and broad-scale questionnaire surveys to determine potential leopard presence within more widespread areas of the leopard's historical range. The surveys also detected other predators, prey and livestock species, and their distributions are discussed here in the context of leopard habitat suitability and the consequences for any future leopard reintroductions.
Study areas
Although Saudi Arabia is characterized by extreme heat during the summer (up to 50 °C), the study area focused on the historical leopard range that spans the cooler Sarawat Mountains, which run parallel with the Red Sea and comprise the Hejaz Mountains in the north and the Asir Mountains in the south (Fig. 1). These steep mountains, incised by deep wadis (ephemeral river valleys), rise from 300 m altitude in the north to just over 3,000 m in the south, where the Asir Mountains are dominated by a massive escarpment. The northern Hejaz mountains are lower and more broken and are dominated by high sandstone peaks and black volcanic plateaus. Although there is an aridity gradient from the more mesic south to the drier north, the entire area is arid and permanent water bodies are rare, although there can be up to 230 mm of annual rainfall at high elevations in the Asir Mountains (Hasanean & Almazroui, Reference Hasanean and Almazroui2015). The mountainous vegetation is dominated by thorn trees (including Vachellia spp. and Ziziphus spina-christi) at lower elevations and juniper and olive (Juniperus and Olea spp.) woodlands at higher elevations.
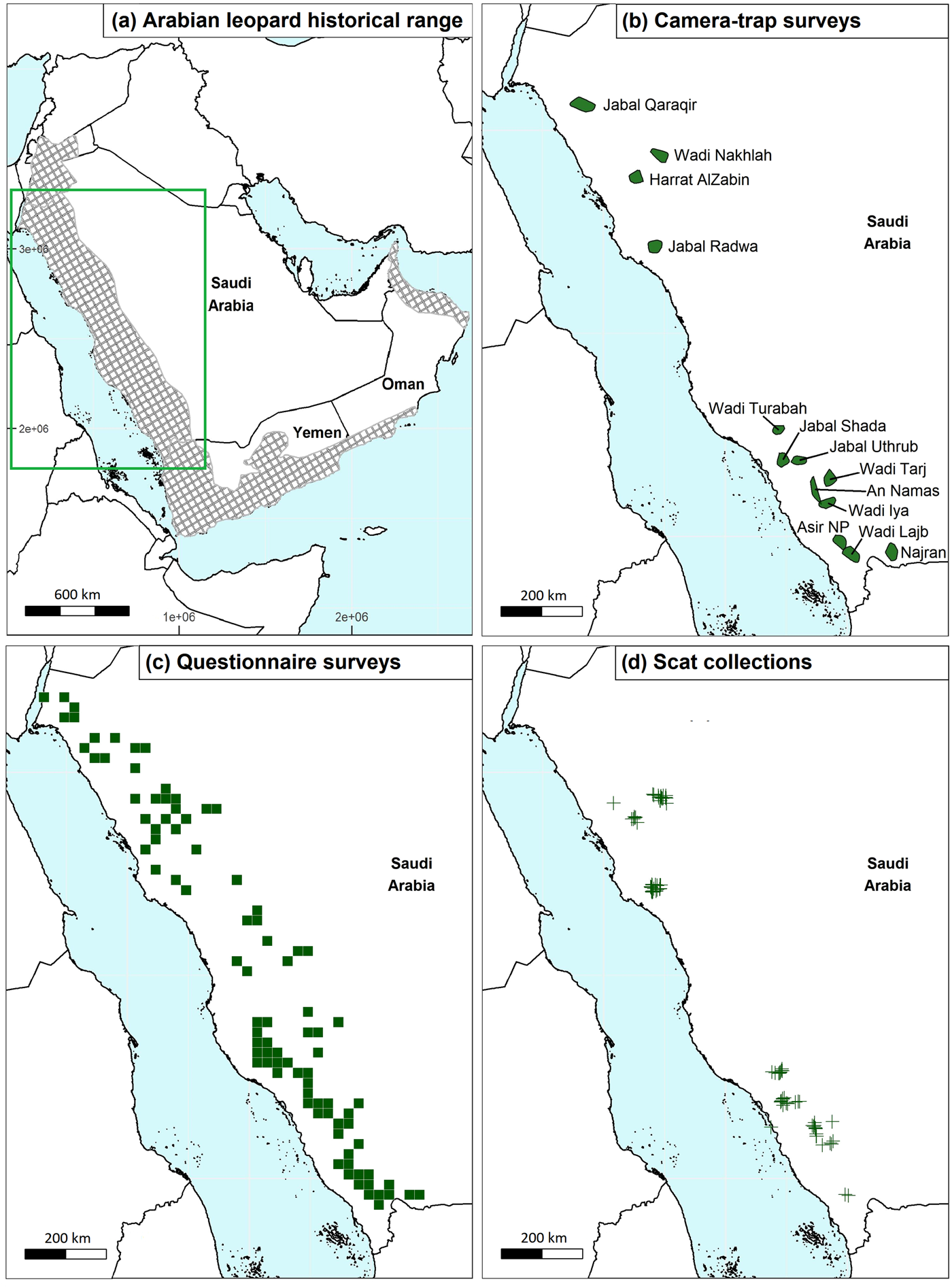
Fig. 1 (a) The historical range of the Arabian leopard Panthera pardus nimr, (b) camera-trap survey sites, (c) grid cells in which we conducted questionnaire surveys, and (d) scat collection locations (mostly at camera-trap survey sites). The rectangle in (a) indicates the area shown in (b)–(d).
Human settlements are present across the study area; tarmac highways connect the larger cities and towns, and gravel roads and tracks provide access to more remote areas (Islam et al., Reference Islam, Boug, Judas and As-Shehri2018). All camera-trap survey sites contained either permanent or seasonal human habitations and were permeable to livestock (comprising sheep, goats, cattle and camels), free-roaming feral donkeys, and poachers, regardless of the area's protection status.
Methods
Although camera trapping is a robust method for identifying species presence in challenging environments, it is an impractical method for surveying the thousands of square kilometres of the leopard's historical range. We therefore initially identified 10 sites for camera-trap surveys through focus groups and leopard habitat modelling (see below). We also conducted questionnaire interviews with local people regarding wildlife, and we identified species from DNA in carnivore scats. We then conducted four further camera-trap surveys where questionnaire or scat results suggested potential leopard presence, including three new sites and one repeat survey of one of the initial 10 sites.
Camera-trap surveys
We determined the areas where we initially deployed camera traps through expert knowledge and habitat modelling. In 2019, we consulted local wildlife experts, in three focus-groups, to identify potential survey areas and the environmental factors most likely to influence Arabian leopard presence. We used these discussions to produce a habitat model based on environmental variables, which formed the basis for preliminary camera-trap site selection. Secondly, we modelled areas of likely leopard habitat using a resource selection function developed using GPS data from African leopards inhabiting semi-arid regions and verified using African leopard and Omani Arabian leopard GPS data, as described previously (Dunford et al., Reference Dunford, Martins, Mann, Spalton, Al Hikmani and Robinson2022). We used the model to classify habitats in the historical range of Arabian leopards in Saudi Arabia, and we identified areas that could potentially sustain leopard populations based on home range size (Fig. 1). In early 2021, we presented this model at a workshop with local wildlife experts, with the aim of confirming the appropriateness of the specific sites to be surveyed and identifying any sites that could contain extant leopard populations that fell outside the areas identified by the models. This workshop was attended by representatives from the Saudi Arabian statutory conservation authorities, academic institutions, non-profit organizations and Arabian leopard experts from Oman. We selected and surveyed an initial 10 sites (see Supplementary Fig. 1, for deployment dates, and Supplementary Table 1). We then selected four further sites for camera-trap surveys through further discussions with experts, anecdotal and questionnaire indications of recent leopard sightings, and scat results. We repeated the survey of one of the 10 initial sites, Harrat AlZabin (henceforth Harrat AlZabin 2). Five camera-trap survey sites were formally protected areas (Jabal Shada, Wadi Nakhlah, Harrat AlZabin, Jabal Qaraqir and Asir National Park), and although some other survey areas were in proposed or planned protected areas, these were not formalized at the time of the study.
The 14 surveys covered a total of 4,189 km2 (mean 299 ± SD 125 km2, range 117–662 km2). They spanned the south-western Hejaz and Asir mountains and areas to the north. During August 2020–March 2022, we deployed PantheraCam V6 and V7 camera traps (Panthera, New York, USA) at 586 stations across the 14 surveys. Cameras were deployed at 3–5 sites simultaneously, and each survey contained c. 40 stations. We deployed two cameras at each station up to 500 m apart and, to maximize leopard capture probability (Du Preez et al., Reference Du Preez, Loveridge and Macdonald2014; Loveridge et al., Reference Loveridge, Sousa, Seymour-Smith, Mandisodza-Chikerema and Macdonald2022), we did not set them in the same locations. We deployed each of the two cameras in places that were likely to be used by leopards such as on game/livestock trails, in valleys or on ridge lines, and set them seperately to cover at least two potential traversal routes (mean distance between cameras: 164 ± SD 183 m). We deployed the cameras at a height of c. 40 cm, on available supports such as rocks or trees.
Stations were c. 3 km apart and dispersed evenly across each survey site. We deployed the cameras for 8–12 weeks (which should be sufficient to capture any Arabian leopards in the area; Islam et al., Reference Islam, Boug, Judas and As-Shehri2018) and checked them at least once during this time to change the batteries and memory cards. If we found any cameras to have malfunctioned, we replaced them. We created a camera-trap effort matrix for each station, denoting the days when each camera recorded images and days when it was deployed but did not record images (e.g. when the batteries were depleted).
Using PantheraIDS (Panthera, New York, USA), we identified species in the camera-trap photographs, initially using a machine learning algorithm (Willi et al., Reference Willi, Pitman, Cardoso, Locke, Swanson and Boyer2018), after which one of the author team manually verified all photos and corrected any misidentifications. This enhanced the processing speed for the dataset as identifications by machine learning could be confirmed en masse rather than labelled individually. We considered photographs of the same species more than 30 min apart to be independent and included these in subsequent analyses. For each site we calculated Shannon's diversity index as it accounts for both species richness and evenness (Spellerberg & Fedor, Reference Spellerberg and Fedor2003). We included only wild species, excluding all domestic and feral species, people and vehicles.
Questionnaire surveys
We designed the questionnaires to determine the presence of leopards, other large carnivores and prey species across the leopard's historical range. We interviewed multiple participants in the same survey cell, to generate replicates (Zeller et al., Reference Zeller, Nijhawan, Salom-Pérez, Potosme and Hines2011).
We divided the historical leopard range into grid cells of 625 km2 (25 × 25 km). As surveying the whole of this area was infeasible, we aimed to survey 20% of cells (Thorn et al., Reference Thorn, Green, Keith, Marnewick, Bateman and Cameron2011; Melovski et al., Reference Melovski, von Arx, Avukatov, Breitenmoser-Würsten, Đurović and Elezi2018). We selected 247 cells as potential targets for surveying based on random sampling as well as overlap with camera-trap survey areas or potential leopard habitat (as predicted by habitat selection modelling; Dunford et al., Reference Dunford, Martins, Mann, Spalton, Al Hikmani and Robinson2022; see Supplementary Material 1 for further details). The field team surveyed 109 of these 247 grid cells, prioritized based on overlap with camera-trap survey areas, likelihood of leopard presence and logistical considerations such as the presence of people to survey (desert areas are sparsely populated).
We aimed to interview 10 participants within each cell. We selected participants based on identification of local people who were knowledgeable about wildlife in the area, both Saudis and resident expatriates. We considered people to be suitably knowledgeable regarding a particular grid cell if they had lived or worked in the area for at least 3 months during the previous year. We conducted only one interview at a time, but we sometimes conducted multiple interviews in one location, with interviews carried out in person at the homes or places of work of participants. We recorded the geographical coordinates and elevation of the interview location. Interviews were conducted during May–October 2021, in Arabic, by Saudi authors MA and SA.
To assess the credibility of any reported animal sightings, we asked interviewees to identify pictures of local wildlife species: the Arabian leopard, other predators (the striped hyaena Hyaena hyaena, Arabian wolf Canis lupus arabs and caracal Caracal caracal), and prey (the Arabian gazelle, Nubian ibex, Cape hare and rock hyrax; see Supplementary Material 1 for interview details). After showing interviewees each picture, we asked whether they had seen the species or any signs of it (e.g. tracks/scat) within the last year. For the leopard, we also asked participants when they had last heard of one in the area and, if they had, when and where it had been sighted. We grouped leopard sighting dates into four categories: within the last year, within the last 10 years, within the last 20 years or > 20 years ago.
Scat collection and DNA surveys
The camera-trap survey field teams opportunistically collected samples of any carnivore scats they encountered whilst hiking to and from camera-trap stations. The teams also collected scats from other sites at which they conducted reconnaissance inspections. We stored dried scats in sealed plastic bags marked with the date, elevation and geographical coordinates of the collection site.
We extracted DNA from dry scat samples at ZooOmics (Inqaba Biotec, Pretoria, South Africa). We amplified a fragment of mitochondrial DNA (16S) for each sample and analysed this against publicly available reference sequences to determine species. We also manually compared these sequences to those of known Arabian leopards using scat samples collected from two captive individuals at the Arabian Leopard Breeding Centre in the National Wildlife Research Centre near Taif, Saudi Arabia. We targeted an additional leopard-specific fragment (NADH5; Al Hikmani, Reference Al Hikmani2019) in all confirmed felid samples; successful amplification of this fragment would conclusively identify the sample origin as that of a leopard.
Results
Camera-trap results
The camera traps recorded 941,166 photographs over a total of 82,075 trap-nights. After removing blank or unclassifiable images, we identified animals in 464,832 images, including 62,948 independent captures of animals or people, but none of the Arabian leopard.
We recorded 29 animal species (18–27 per study site; Fig. 2), with species accumulation curves reaching an asymptote by 29 days at all sites (Supplementary Fig. 2). Amongst the species with fewest detections were two of the main wild prey species of the Arabian leopard: the Arabian gazelle (24 independent captures in two surveys) and Nubian ibex (106 in eight surveys). Independent captures of all wild prey species were generally low across all sites (a total of 9,602 captures of Arabian gazelle, Nubian ibex, rock hyrax, Indian crested porcupine Hystrix indica, Cape hare and partridges Alectoris philbyi, Alectoris melanocephala and Ammoperdix heyi). Predators were recorded in high numbers, and red foxes Vulpes vulpes, Arabian wildcats Felis lybica lybica and Arabian wolves were recorded at all sites (Fig. 3). Arabian wolves were the most common large predator (2,663 independent captures in 14 surveys), whereas caracals and striped hyaenas were mostly recorded at the nine southern sites (497 and 1,807 independent captures, respectively), with the striped hyaena recorded at one site in the north, Jabal Qaraqir. The most common wild mammals recorded were Hamadryas baboons Papio hamadryas (7,528 independent captures) and red foxes (4,962), although Hamadryas baboons were only recorded at the nine southern sites.

Fig. 2 Number of independent photographic captures of all wild species, human presence and domestic and feral species at all 14 camera-trap survey sites in Saudi Arabia (Fig. 1b). (Readers of the printed journal are referred to the online article for a colour version of this figure.)

Fig. 3 Independent photographic captures per trap-night by survey site for (a) large predator species, (b) wild prey species, and (c) domestic and feral species, humans, and vehicles. (Readers of the printed journal are referred to the online article for a colour version of this figure.)
Domestic livestock herds and feral livestock were the most commonly recorded animals; we obtained twice as many independent captures of livestock (a total of 18,168 independent captures of cattle, sheep, goats, camels and donkeys) as of wild prey. We also recorded people at all sites, at 72% of our 586 camera stations (3,472 independent captures after removing photographs of field team members on days when the cameras were set up, checked and taken down). We also recorded a further 1,902 independent captures of vehicles. Feral donkeys were the most common species recorded (8,675 independent captures), but over one-third of these were at one site, Wadi Turabah. Camels, goats and sheep were prevalent at all sites (4,241, 3,439 and 1,262 independent captures, respectively), suggesting widespread human impacts across our study areas.
Asir National Park had the highest number of large predator and prey captures per trap-night and the second highest captures of livestock (Fig. 3). Wadi Iya had the second highest captures of large predators. Wadi Iya, the Najran Highlands and Wadi Lajb also had high captures of prey species and, together with Asir National Park, are the four most southerly sites, suggesting the highest densities of prey are in the southern region of the leopard's historical range. Sites in the south also had higher wild species diversity than those in the north (Shannon's Diversity Index 1.71–2.36, t Student(12) = −3.44, P < 0.01; see Supplementary Figs 3–11 for mapping of independent captures at all camera stations).
Questionnaire results
We completed 843 questionnaire interviews across 109 grid cells. In each grid cell we conducted 7.7 ± SD 4.4 (range 1–28) interviews. The majority of the participants were farmers (72.5%; others included retired or unemployed people and military personnel) and lived or worked in the region (35.5% and 52.7%, respectively).
Participants correctly identified leopards from photographs 96.6% of the time. Of the participants that correctly identified the leopard image, 731 (89.8%) had never heard of one in the area, whereas 83 participants (10.2%) had either heard of a leopard in the area or seen one. Sixty-six of these sightings were from > 20 years ago, five were from within the last 20 years, nine were from within the last 10 years and three were from within the last year. The latter three were all direct sightings in different locations, with participants reporting seeing the leopard multiple times near a house, crossing a road or in the mountains. These three respondents identified all of the wildlife species correctly from the photographs.
Amongst those participants that correctly identified the species in the images, predator sightings were common, with 82.9% having seen an Arabian wolf within the last year, 59.6% having seen a striped hyaena and 39.5% having seen a caracal. Large prey species were far less commonly seen than predators: 17.7% of participants had seen a Nubian ibex and 8.5% had seen an Arabian gazelle in the last year. Smaller prey species were more commonly seen, with 65.8% of respondents having seen a Cape hare and 78.2% having seen a rock hyrax in the last year (Fig. 4, Supplementary Fig. 12; see Supplementary Material 2 for additional details).

Fig. 4 Sightings of large predators and potential prey species within the last year reported from questionnaire surveys conducted in the historical range of the Arabian leopard in Saudi Arabia. Grid cells are shaded if at least one questionnaire participant correctly identified the species from a photograph and reported sighting the species either directly or indirectly. (Readers of the printed journal are referred to the online article for a colour version of this figure.)
Scat and DNA results
We obtained scats from 10 survey sites, and of the 143 scats analysed, the species of 91 could be identified. The remaining 51 samples were too degraded for DNA to be amplified. The majority of the identified samples were identified as domestic dog (70), but striped hyaena (4), caracal (3), red fox (7), Arabian wildcat (6) and feral domestic cat (1) were also detected. It is probable that many of the samples identified as feral dogs actually originated from Arabian wolves; these species are genetically similar but we did not have an Arabian wolf sample to confirm identity unequivocally. One further sample, from Harrat AlZabin, was tentatively identified as leopard. However, this scat was too degraded to make a definitive assessment; it originated from a felid and was genetically distinct from caracal but probably too large to be a wildcat scat. Attempts to verify the identity using the NADH5 genetic primer failed because of the degradation of the DNA fragments extracted from this scat, so we obtained no conclusive species identity.
Discussion
We used data from camera-trap surveys, questionnaires with local people and DNA analysis to survey the historical range of the Arabian leopard in Saudi Arabia in an effort to establish whether the leopard is still present. Despite a search effort that is, to our knowledge, amongst the most comprehensive attempts ever undertaken to locate a wild felid, we found no conclusive evidence of presence. Furthermore, even the prime wildlife areas surveyed showed substantial degradation from anthropogenic activities, and most sites were affected by livestock grazing and had high human presence. The likelihood of a sustainable population of Arabian leopards surviving in this landscape, with limited wild prey, is low.
Our camera-trap surveys in Saudi Arabia spanned 586 stations in 14 sites, with 464,832 images of animals or people. In comparison, a study with 12 camera-trap stations obtained 251 records of Arabian leopards in Oman during 1997–2000 (Spalton et al., Reference Spalton, Al Hikmani, Willis and Said2006b), and in 2013 in the desert landscapes of Namibia, eight African leopards were identified using 51 camera-trap stations (Edwards et al., Reference Edwards, Aschenborn, Gange and Wiesel2015). These findings suggest that we should have captured evidence of leopards if they were present. Furthermore, both expert and model predictions suggested that the Saudi Arabian sites surveyed were the areas in which leopards were most likely to persist. In addition, species accumulation curves reached an asymptote at every site by 29 days, well within the 75-day mean activity duration of the cameras (Supplementary Fig. 2). Consequently, it seems unlikely that leopards were present at any of our survey sites and that sustainable populations of leopards are still present in this landscape.
The questionnaire results revealed three potential leopard sightings across the species' historical range in the year prior to the interviews. Although it is encouraging that there could be a few individuals left in the wild, these sparse reports suggest these would probably be lone individuals that are not part of a viable population. The interviewees reporting these leopard sightings confidently described the scenario and correctly identified all of the species in images shown during the interview, suggesting they were familiar with the local wildlife and should be able to correctly identify a leopard. However, no sightings were corroborated by other respondents in the same area or by focal camera trapping in these areas in the second phase of the camera-trap surveys (see Supplementary Fig. 13 for leopard sighting locations).
Similarly, the scat survey suggested there is unlikely to be a population of leopards surviving in any of the locations where scats were collected, as we found no confirmed leopard DNA. Our survey was successful at gathering data on other medium and large carnivores living in the western Hejaz and Asir mountains. Caracal and striped hyaena DNA were only found at the southern sites, and the northern sites were dominated by Arabian wolf or feral dog DNA. No scats were analysed from Jabal Qaraqir, the one northern site where striped hyaenas were recorded by the camera traps, explaining why they were not detected in the DNA survey there. Overall, the DNA analysis results largely corresponded with the species detected in our camera-trap surveys, suggesting that DNA analysis of scat samples is a useful alternative method of surveying wildlife, and carnivores in particular. Although a few scats collected were still moist, the majority were desiccated because of the desert climate, and it is encouraging that DNA could be extracted from 64% of these.
The discovery of a potential leopard scat in Harrat AlZabin, in the north, would be the first record of a free-roaming leopard in Saudi Arabia since 2014. However, even if the scat was of leopard origin, in combination with the camera-trap survey results this suggests there are no viable leopard populations in Saudi Arabia. Furthermore, although we conducted a camera-trap survey in Harrat AlZabin during November 2020–March 2021 and a second larger survey in this location during December 2021–March 2022, no conclusive evidence of leopards was found. Additionally, the most recently confirmed leopard sighting, in 2014, was in the Asir Mountains in the south (Islam et al., Reference Islam, Boug, Judas and As-Shehri2018). The scat from Harrat AlZabin was collected in November 2020, so if it did originate from a leopard it is possible the individual could have moved out of the area or been killed in the interim. Because of the poor quality of the sample we caution against making management decisions without further evidence of contemporary leopard presence in Harrat AlZabin.
There were substantiated killings of leopards during 1970–2014 in Saudi Arabia, with a mean of 7.5 leopards per decade (Islam et al., Reference Islam, Boug, Judas and As-Shehri2018), despite no official records being kept. More recently, 2014–2022 is the longest period in which no leopard killings have been reported, potentially suggesting an absence of the species. Considering the wide prevalence of camera-equipped smartphones in Saudi Arabia and the novelty value of leopard sightings, it seems improbable that viable leopard populations could remain undetected for such a relatively long period. Leopards are occasionally photographed with smartphone cameras in Oman (J.A. Spalton, pers. obs.), suggesting they would probably be recorded in Saudi Arabia if they were present. Historically, leopard populations in southern Saudi Arabia would have been connected to populations in northern Yemen; however, the ongoing military conflict in the region and the recent construction of a border fence are likely to significantly impede leopard movement, reducing the likelihood of natural dispersal of leopards into Saudi Arabia from Yemen.
Considering the relatively intact carnivore guild we recorded, we would expect there to be more numerous wild prey, and the low abundance or apparent absence of wild prey species recorded in both the camera-trap and questionnaire surveys highlights a point of concern for the survival of wild leopards in Saudi Arabia (Carbone & Gittleman Reference Carbone and Gittleman2002; Karanth et al., Reference Karanth, Nichols, Kumar, Link and Hines2004; Drouilly et al., Reference Drouilly, Nattrass and O'Riain2018). Arabian gazelles are one of the primary prey species of leopards (Muir-Wright, Reference Muir-Wright1999), but we detected them at only two of the 14 sites surveyed, with only 24 independent records out of the total of 62,948 for all animals. Nubian ibexes were not detected in six of the 14 surveys, and Cape hares were not detected in four. The questionnaire surveys similarly reported a dearth of Arabian gazelles and Nubian ibexes, but there were more reported sightings of Cape hares in the questionnaire surveys than captured by the camera traps. This could be because the camera-trap surveys were designed to detect relatively large mammal species and may have under-detected smaller species such as the Cape hare and rock hyrax.
In contrast to the low numbers of wild prey recorded, the camera-trap survey recorded livestock at > 80% of the camera stations and in all 14 surveys. The wealth of livestock as potential prey is likely to be supporting the relatively large predator guild (see Supplementary Figs 3–11 for mapping of independent species records; Puls et al., Reference Puls, Teichman, Jansen, O'Riain and Cristescu2021); however, livestock predation will probably result in shooting, poisoning or trapping of predators by herders in attempts to decrease livestock losses. Retaliatory killing of leopards (Islam et al., Reference Islam, Boug, Judas and As-Shehri2018) is a threat that would need to be addressed if leopards are to survive in this landscape. Modelling of both African and Arabian leopard habitat selection suggests areas with high human presence are avoided (Fattebert et al., Reference Fattebert, Robinson, Balme, Slotow and Hunter2015; Islam et al., Reference Islam, Boug, Judas and As-Shehri2018; Dunford et al., Reference Dunford, Martins, Mann, Spalton, Al Hikmani and Robinson2022). South African leopard populations in protected areas have also been shown to be limited by the impacts of people and domestic animals (Rogan et al., Reference Rogan, Distiller, Balme, Pitman, Mann and Dubay2022). Prey density was the most important variable determining habitat suitability for Persian leopards Panthera pardus saxicolor in Iran (Khosravi et al., Reference Khosravi, Hemami and Cushman2017), and is also likely to be an important factor in Saudi Arabia.
Arabian leopards might have been extirpated whilst other predators persist because of threats that affect leopards to a greater extent than other species. Trophy hunting, consistently returning to poisoned prey items, slow reproduction rates and historically low densities are likely to have exacerbated leopard population declines (Islam et al., Reference Islam, Boug, Judas and As-Shehri2018). Caracals, however, although the most similar local carnivore to Arabian leopards, can persist in relatively high numbers near human settlements and adapt their behaviour to changing landscapes and opportunities, so they are probably less affected by stochastic events and developmental activities (Leighton et al., Reference Leighton, Bishop, Merondun, Winterton, O'Riain and Serieys2022).
Although we did not conclusively detect leopards in Saudi Arabia, we would expect their distribution to be influenced by interactions with other carnivores (Thapa et al., Reference Thapa, Malla, Subba, Thapa, Lamichhane and Subedi2021). Although multi-species occupancy modelling was not our objective, the occurrences of caracals and striped hyaenas were concentrated in the south and Arabian wolves were found throughout the study region (Fig. 4). Other carnivores could provide scavenging opportunities for the leopard, although groups of Arabian wolves could impose costly competition for scarce resources, dominate a lone leopard at a kill or pose a direct threat to young leopards (Balme et al., Reference Balme, Batchelor, de Woronin Britz, Seymour, Grover and Hes2013; Elbroch et al., Reference Elbroch, Lendrum, Newby, Quigley and Thompson2015). Further research into the historical and current carnivore guild structure and interactions would shed light on how these species might support or suppress recovering Arabian leopard populations in the region. The disparity in predator and prey species distributions and abundances suggests that the most intact wildlife guild occurs in the south and that the higher prey density there could potentially support a leopard population.
Conclusions
It is unlikely that any viable populations of the Arabian leopard remain in Saudi Arabia. Although one or two lone animals might survive, extensive camera-trapping, questionnaire and DNA surveys did not provide any conclusive evidence of leopard presence. Regardless of whether leopards have been completely extirpated or reduced to a small number of isolated individuals, it is almost certain that leopard populations will not recover in Saudi Arabia without artificial supplementation. Ex situ conservation and future reintroduction of leopards to their historical range are likely to be the only means of restoring the species in Saudi Arabia. Initially, the restoration of wild prey populations would be essential, to deter leopards from hunting livestock and to prevent conflict with herders. Further research into leopard habitat and prey species, the predator guild and coexistence with people in Oman and Yemen, where populations of Arabian leopards persist, could inform leopard re-establishment efforts in Saudi Arabia.
Acknowledgements
This work was supported by the Royal Commission for AlUla, the National Center for Wildlife and the Prince Mohammed Bin Salman Reserve in partnership with Panthera. We thank Maya Bankova-Todorova, Salima Raai and Musa Abdullah Elhassan for their logistical and administrative support; everyone who assisted with fieldwork, including Lucy Smyth, Josh van der Ploeg, Mathew Smith, Clement Brunet, Jacques Proust, Charles Spalton and Nic Proust; and Lawrence Steyn for assisting with image classification.
Author contributions
Conceptualization and methodology: CED, JPBF, JAS, GB, AAlmalki, EG, GM; funding acquisition: GB, AAlmalki, EG, GM; project administration: JPBF, GB, AAlmalki, EG, DRM, GM; fieldwork: CED, JPBF, MDR, JAS, MD, KJPP-F, RDB, AEB, AAlshehri, MA, SA, GM; data curation: CED, JPBF, MDR, JAS, MD, KJPP-F, RDB, AEB, NLR, GM; visualization: CED, MDR, GM; analysis and writing: CED, GM; revision: all authors.
Conflicts of interest
None.
Ethical standards
The regional governing bodies, the Royal Commission of AlUla and the Kingdom of Saudi Arabia Ministry of Interior provided permission for the questionnaire and field surveys (permit numbers: 27/645 and 149137). This research abided by the Oryx guidelines on ethical standards and the questionnaire and study methods followed the American Psychological Association core principles. Sensitization to the study was conducted opportunistically when local people were encountered during camera deployment, and they were informed of the purpose of the surveys. Stickers identifying the cameras as the property of Royal Commission of AlUla, in Arabic, were placed on all cameras. Data protocols include Advanced Encryption Standard (AES) 256-bit encryption, both when at rest and during data transit. All images, including those of people, were identified using a machine learning classifier and confirmed manually before being securely stored on an AES 256-bit encrypted server with access limited to approved personnel only. Images of people were securely managed within the Royal Commission of AlUla and Panthera. We were legally required to share photographs of illegal activity recorded within the Prince Mohammed Bin Salman Reserve (i.e. photographs of people carrying weapons or wildlife carcasses) with anti-poaching units. Prevailing local laws and regulations were followed regarding personal privacy and data protection.






 Open access
Open access