


Non-technical Summary
Evolutionary success comes to the selfish—or so most of us are taught. Those organisms that reproduce most will of course be better represented in future generations. Current theory has had great success in understanding much of evolution, but it hits a wall when it comes to understanding how ancestrally solitary organisms can aggregate to form new wholes. Current theory expects that the evolution of new wholes is undermined by the selfishness of the parts; if the parts keep reproducing, an emergent whole cannot originate. Yet every animal is a living contradiction of current theory. During development, animal cells divide in the same way their solitary ancestors reproduced, while concurrently the whole animal reproduces itself.
Are Levels Important?
Of all the ways of thinking of levels in evolutionary biology (Stanley Reference Stanley1975; Van Valen Reference Van Valen1975; Eldredge Reference Eldredge1979; Lieberman et al. Reference Lieberman, Allmon and Eldredge1993), the evolutionary transitions in individuality (Buss Reference Buss1987; Maynard Smith and Szathmáry Reference Maynard Smith and Szathmáry1995; Michod Reference Michod1999) highlight just how tricky it is to make sense of the nature of levels and their formation. These specific transitions share the characteristic of the whole consisting of parts with a free-living ancestry. A rich set of empirical examples occur, but there are not enough transitions at a single level to develop a fully satisfying account of the evolution of new levels. For example, we do not know how nascent multicellularity provides an advantage over unicellular competitors (Fenchel Reference Fenchel2019; Koehl Reference Koehl2021; Simpson Reference Simpson2023), nor do we know if division of labor in terms of multiple cell types is ancestral or derived (Sebé-Pedrós et al. Reference Sebé-Pedrós, Degnan and Ruiz-Trillo2017). These are two important examples, and no doubt other important features of the transition to multicellularity would be discovered if more examples of transitions to new levels are discovered in nature or evolved in the lab (e.g., Bozdag et al. Reference Bozdag, Libby, Pineau, Reinhard and Ratcliff2021; Halling et al. Reference Halling, Goodson, Hirschmann, Wing and Simpson2024).
In this paper, we compare two transitions in individuality from an empirical perspective—building off our own work—to develop some insights about just how important hierarchical structure is for the evolution of organisms. We hope to elucidate the varied ways in which evolutionary transitions in individuality and coloniality can occur through examining characteristics of volvocine algae and colonial bryozoans.
What follows is a sketch of the empirical utility of what paleobiologists call the “emergent fitness” perspective (Lloyd and Gould Reference Lloyd and Gould1993; Grantham Reference Grantham1995; Gould and Lloyd Reference Gould and Lloyd1998). There are other concepts of levels and what makes them identifiable or important, such as the emergent trait approach (Grantham Reference Grantham1995). We focus on tracking the evolution of fitness as it emerges. While the approach we outline may seem like a small conceptual advance, we have found that it has opened the door to more and easier empirical work, including theoretical and experimental ties to life history and metabolic theory (White et al. Reference White, Alton, Bywater, Lombardi and Marshall2022). One consequence we hope will follow from our approach is the broadening of what organisms we consider to be at or near a major transition. For example, it is common to focus on the “complex” multicellular groups (Knoll Reference Knoll2011) to understand the evolution of multicellularity (Simpson Reference Simpson, Calcott and Sterelney2011). But there are many other multicellular species and lineages (Bonner Reference Bonner2001; McShea Reference McShea2001; Costa Reference Costa2006; McShea and Simpson Reference McShea, Simpson, Sterelney, Callcott and Callebaut2011; Herron et al. Reference Herron, Rashidi, Shelton and Driscoll2013; Niklas and Newman Reference Niklas and Newman2013) that do not possess germ-soma division of labor yet have much to teach, not just about multicellularity but also about how transitions occur. And engaging with these empirical examples should help untangle the conceptual issues around levels of selection, particularly when a new level of fitness is gained. Moreover, these questions are especially tractable with experimental evolution (Pentz et al. Reference Pentz, Márquez-Zacarías, Bozdag, Burnetti, Yunker, Libby and Ratcliff2020, Reference Pentz, MacGillivray, DuBose, Conlin, Reinhardt, Libby and Ratcliff2023; Bozdag et al. Reference Bozdag, Zamani-Dahaj, Day, Kahn, Burnetti, Lac and Tong2023; Conlin and Ratcliff Reference Conlin and Ratcliff2023; Pineau et al. Reference Pineau, Demory, Libby, Lac, Day, Bravo, Yunker, Weitz, Bozdag and Ratcliff2023; Zamani-Dahaj et al. Reference Zamani-Dahaj, Burnetti, Day, Yunker, Ratcliff and Herron2023; Halling et al. Reference Halling, Goodson, Hirschmann, Wing and Simpson2024). With the framework we present of tracking fitness at multiple levels, we have a single quantitative method to probe the hierarchical structure of organisms (and species). One hope is that we can use this method to quantify the difference between simple colonies and complex multicellular organisms and the relative contribution of species selection and organismal selection to large-scale trends.
Emergent Fitness Is More Than Births and Deaths
Evolutionary success comes to the selfish—or so most of us are taught. Organisms that reproduce more will of course be better represented in future generations. Current theory has had great success in understanding much of evolution, but it cannot explain how ancestrally solitary organisms can aggregate to form new wholes. Current theory expects that the evolution of new wholes is undermined by the selfishness of the parts; if the parts keep reproducing, an emergent whole cannot originate. Yet every animal is a living contradiction of current theory. During development, animal cells divide in the same way their solitary ancestors reproduced, while concurrently the whole animal reproduces itself.
It is not that our theories are wrong; it is just that what we traditionally call evolutionary success is an overly myopic view. In the formulation we present here, one that is explicitly hierarchical in structure, we consider three general ways for frequencies of units to increase over time and so be considered successful: (1) differential persistence of units, (2) differential multiplication of units, and (3) differential expansion of units. Any combination of these can occur. Only the first two aspects of selection are incorporated into current theory. Expansive fitness looks, from the higher level, like growth. But from the lower level, it appears orthodox, as the differential multiplication of parts. Once one adopts a hierarchical framework, one can incorporate the notion of expansive fitness easily and naturally.
Expansion allows us to translate units of fitness at one level into units at another. Population growth at one level is expansion at the next level up. You can see expansion in action during animal development: through cell division, the animal grows. Metazoans derive from single-celled ancestors, and the ancestral fitness, cell division, is joined by fitness at the whole-animal level.
The important role of differential expansion in evolution has barely been recognized, even though it has long been associated with thought on levels of selection. Expansion may also be the most common mode of multilevel selection (Van Valen Reference Van Valen1975, Reference Van Valen1976; Simpson Reference Simpson, Calcott and Sterelney2011). Lyell (Reference Lyell1832) was the first proponent, evoking the differential expansion of species by competition as a mechanism for faunal turnover. Van Valen (Reference Van Valen1973, Reference Van Valen1975, Reference Van Valen1976, Reference Van Valen1989, Reference Van Valen1991) re-derived and named expansion from his energetic notion of fitness. Using energy control as a yardstick for fitness frees us from counting individual organisms and makes it evident that there are many ways for natural selection to change the frequency of traits in a population over time. The familiar differential persistence and multiplication can do it easily. So too can differential changes in the size of units in a population. Van Valen’s contribution to our understanding of levels of selection has been neglected, possibly due to the conceptual hurdle involved in adopting his energy notion of fitness. But Van Valen's contributions align well with Leo Buss's many insights about the evolution of individuality. Although Van Valen's energy view is useful, and may be correct, we do not have to adopt it to use expansion.
Expansion makes multilevel selection easier. Here is how it works.
Expansion Above, Demography Below
The demographic processes of birth and death that occur within a population determine how the size of that population will change over time. If there are more births than deaths, the population grows; if deaths outnumber births, the population will shrink. The net population growth rate is equal to the average birth rate minus the average death rate for constituent members. Populations grow and shrink depending on the demographic activities of their constituents.
Multiplication, where births are involved, and persistence, where deaths are involved, make up the two usual components of demographic fitness. These aspects of fitness can vary among phenotypically varying members of a population. A consequence of this fitness variation is that the frequencies of each phenotype within the population can evolve.
Now take two populations. These populations differ from each other by varying in either the set of phenotypes that their members have or in the relative fitnesses of their members’ phenotypes. As a consequence of the fitnesses of members within each population, the two populations will grow at different rates.
If the variation among demographic fitnesses is caused by variation in some phenotype, then natural selection occurs. There is nothing new here. This is just the standard view of natural selection in populations. But zoom out and focus on the whole population. From this view, no group divides or disintegrates, it just changes size. Normally, such a change in size is not called natural selection, even if it nearly satisfies the basic criteria for natural selection outlined by Lewontin (Reference Lewontin1970) of variation, heredity, and differential reproductive success. Here, all reproduction that does occur takes place among the constituents of a group. Yet the group expands, and its size may very well have ecological and evolutionary significance (Van Valen Reference Van Valen1989).
There is no substantive difference between focusing on the member or the group levels in this analysis; rather, it illustrates an important point about expansion and group-level fitness. Traditionally, group fitness is taken to be equal to the mean member fitness (termed the Group Mean Fitness Orthodoxy, for an in-depth review and discussion, see Okasha Reference Okasha2006). But distinguishing between the expansive and demographic components of fitness shows how limited this assumption is. Reproduction is different from growth, and we should not automatically equate them, even though they can be correlated in many organisms. Rather, as we shall argue, the breaking of their relationship is a crucial step in the evolution of new levels or organization.
From a conceptual point of view, expansion is likely to be important in the shift from multilevel selection type 1 (MLS1) to type 2 (MLS2) (Eldredge Reference Eldredge1979; Damuth and Heisler Reference Damuth and Heisler1988; Okasha Reference Okasha2006). Damuth and Heisler (Reference Damuth and Heisler1988) clarified the key differences between the group selection and paleobiological species-selection frameworks. In MLS1, group fitness is a function of the average fitness of group members. A Gonium colony, for example, develops eight cells that eventually form eight colonies. In contrast, for MLS2, the fitness of the whole group is its own quantity, independent of the fitnesses of members. This was first recognized as the disconnection between speciation and extinction rates and organismal fitness in the paleobiological literature on species selection (Eldredge and Gould Reference Eldredge, Gould and Schopf1972; Stanley Reference Stanley1975, Reference Stanley1979; Arnold and Fristrup Reference Arnold and Fristrup1982; Vrba and Gould Reference Vrba and Gould1986; Gould and Eldredge Reference Gould and Eldredge1988; Grantham Reference Grantham1995). In major transitions, such as the origin of multicellularity and coloniality, the dominant level of fitness shifts to the new hierarchically more-inclusive level in a way that seems to match the shift from MLS1 to MLS2 (Okasha Reference Okasha2006).
This shift is commonly thought to occur through the suppression of the lower level of fitness (Michod Reference Michod1999; Okasha Reference Okasha2006; Abbot et al. Reference Abbot, Abe, Alcock, Alizon, Alpedrinha, Andersson and Andre2011). But that is not the only evolutionary path open to organisms undergoing a major transition (Simpson Reference Simpson, Calcott and Sterelney2011, Reference Simpson2012; Simpson et al. Reference Simpson, Herrera-Cubilla and Jackson2020). Recall Lewontin's (Reference Lewontin1970) criteria for evolution by natural selection: variation, inheritance, and differential success. Evolution by natural selection will not occur if any of those three criteria are absent. Suppression of the lower level of fitness occurs by limiting differential success of constituents. But the suppression of inheritance at the lower level also occurs. For example, in the colonial bryozoan Stylopoma, heritability at the lower part level is lost (Simpson et al. Reference Simpson, Herrera-Cubilla and Jackson2020), while heritability at the level of the colony occurs (Simpson et al. Reference Simpson, Herrera-Cubilla and Jackson2020; Leventhal et al. Reference Leventhal, Jamison-Todd and Simpson2023). With this, the evolutionary potential of the parts is neutralized, but their ability to increase in numbers remains. In the example of Stylopoma, the loss of heritability permits the growth of the colony by differential birth and death of zooids, and with the presence of heritability and fitness at the colony level (Simpson et al. Reference Simpson, Herrera-Cubilla and Jackson2020; Leventhal et al. Reference Leventhal, Jamison-Todd and Simpson2023), evolution occurs at the level of the colony. Both the Stylopoma example and the thought experiment considering a case in colonies where growth is important for ecological (and therefore evolutionary) success highlight just how easy transitions to new levels of organization can be. They also impart a sense of the evolutionary paths open to undergo a transition. Selection (in our broader sense) at a higher level occurs right away, once zooids remain connected together.
To illustrate the potential evolutionary paths open for transitions, we plot in Figure 1 an illustration and comparison of the differences between MLS1 and MLS2. We simplify things by ignoring inheritance and variability and focus on just two levels of population growth rate. Along the x-axis is the growth rate of the members (e.g., the individual Chlamydomonas cells or single cells within a Gonium colony), and along the y-axis is the growth rate of the whole (populations of cells for the unicellular Chlamydomonas or populations of colonies for Gonium and Volvox). In MLS1, group fitness is defined to be equal to the average fitness of members (Okasha Reference Okasha2006), thus evolution under MLS1 is bound to be along the diagonal, where growth rates at the level of the whole are equal to the average growth rate of the parts. And only when a selection gradient moves up toward higher fitnesses, signified by the arrow in the Figure 1, does evolution occur at the group level. The situation for MLS2 is starkly different. MLS2 has no constrained definition equating finesses at the two levels (Damuth and Heisler Reference Damuth and Heisler1988; Okasha Reference Okasha2006; Simpson Reference Simpson, Calcott and Sterelney2011), and evolution of the group will occur if selection acts to move from central point in Figure 1 to anywhere within the gray box. This area has a net positive fitness at the level of the whole, and it does not matter what the part-level fitness does, including evolution along the left side of the graph, where the growth rate of constituents can decrease.
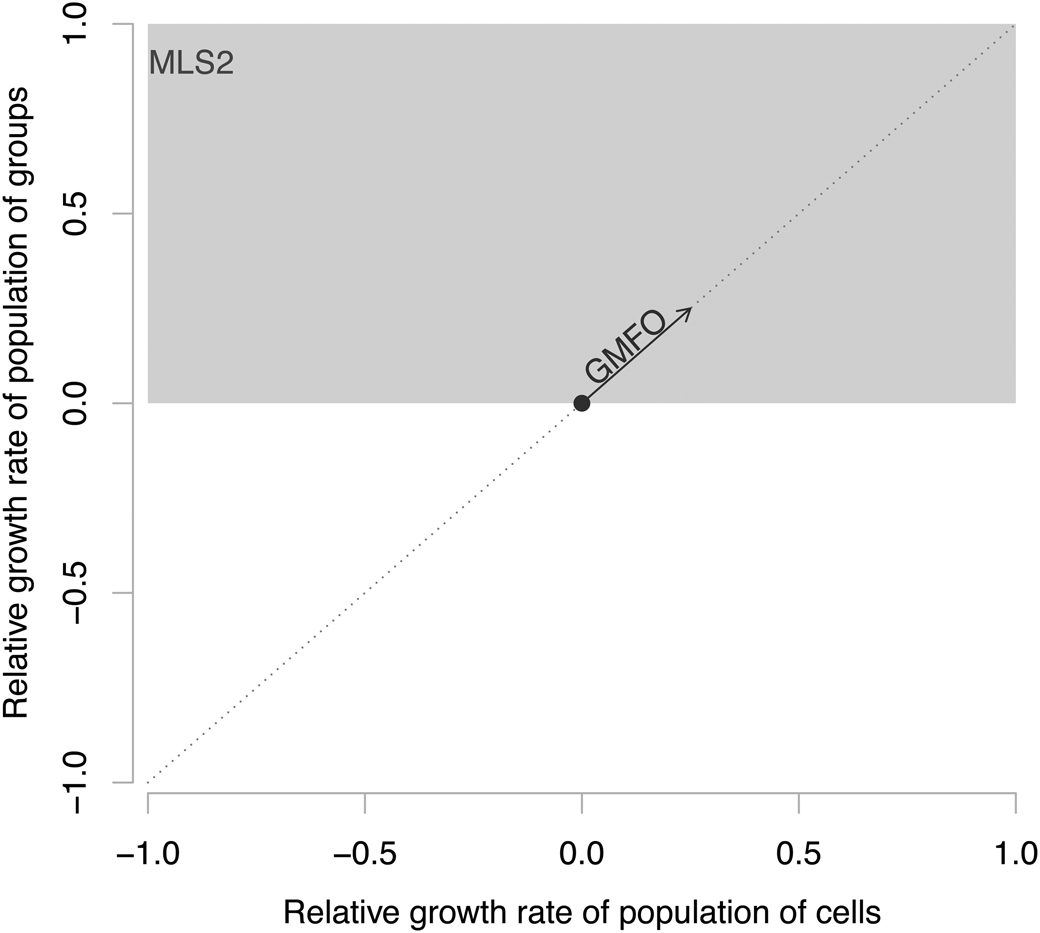
Figure 1. An illustration of a quantitative way to distinguish between group mean fitness orthodoxy (GMFO) encompassed by multilevel selection type 1 (MLS1) and multilevel selection type 2 (MLS2). GMFO occurs when group fitness is equal to the average member fitness and here would plot along the one-to-one line. In contrast, the group fitness in MLS2 is independent of the member fitness. In both, for groups to evolve, they need to increase the relative growth rate of populations of groups, so both require an increase along the y-axis. GMFO is constrained in its path, but MLS2 can occur anywhere within the gray area.
If the part-level fitness is freed up, as with the example of Stylopoma, and lacks evolutionary potential from a loss of heritability, then the demographic pattern shown on the x-axis of Figure 1 becomes a measure of growth of individual colonies.
The Decoupling of Fitnesses across Multicellular Volvocine Algae and Bryozoan Colonies
The shift from MLS1 to MLS2 and the accompanying redirection of part-level fitness into growth can easily be seen empirically in the volvocine algae and in the colonial bryozoans. In Figure 2, we plot growth rates for colonies against the growth rates of populations of colonies (similar to the plot in Fig. 1). The diagonal line signifies an MLS1 situation in which the growth rate of populations of colonies is equal to the growth rate of cells (or zooids) within the colony corresponding to the Group Mean Fitness Orthodoxy.
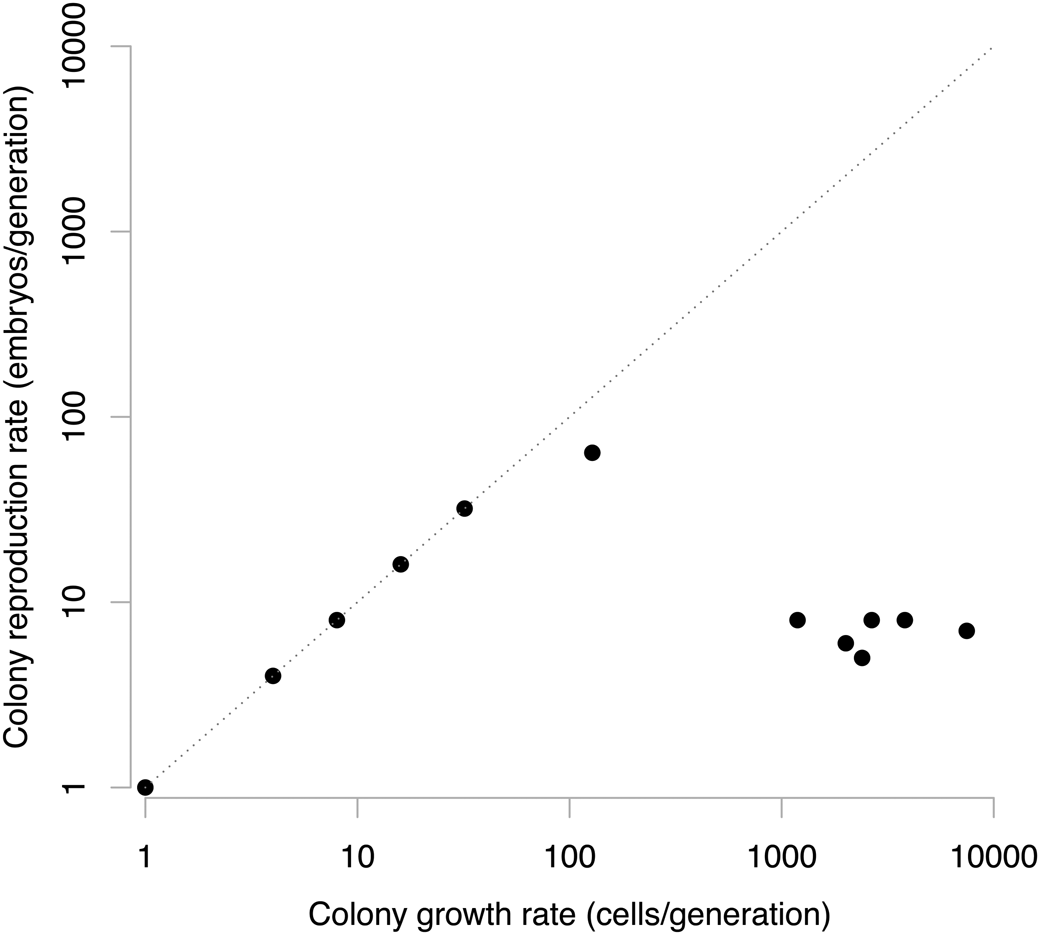
Figure 2. The cell- and colony-level demographics of 12 species of volvocales algae. The diagonal one-to-one line here is similar to that in Fig. 1, where multilevel selection type 1 (MLS1) is constrained to operate. In the volvocales, we can observe a shift from an MLS1-dominated situation in Chlamydomonas, Tetrabenia, Gonium, and Pandorina. However, Pleodorina and species of Volvox plot within an off-diagonal region characterized by multilevel selection type 2 (MLS2), where colony reproductive rates are independent of a colony's somatic growth. Data from Koufopanou (Reference Koufopanou1994).
The volvocine algae include unicellular, simple colonial, and multicellular species such as those in the polyphyletic genus Volvox. Using cell number data from Koufopanou (Reference Koufopanou1994), we plot the pattern of somatic growth rate and colony reproductive rate for a dozen species of volvocales (Fig. 2). In the unicellular Chlamydomonas, its asexual life cycle involves the division of the mother cell into 4, 8, and, rarely, up to 32 daughter cells that break out of the extracellular membrane before dispersing as unicells (Kirk Reference Kirk1998), and so it plots on the one-to-one line in Figure 2. Similarly, in the multicellular Gonium, each cell of the colony divides into a typically eight-celled offspring colony before dispersing as a unit. In Gonium, all constituent cells give rise to new colonies, and so it is a clear example of MLS1, as the cell-level growth rate is equal to the colony-level growth rate.
This coupling between fitness at the cell and colony levels continues with other volvocales genera such as Pandorina, but begins to break down with the genus Pleodorina, which consists of equal numbers of somatic and germ cells. Only the germ cells give rise to the next generation of Pleodorina colonies. This pattern is also broken in species of Volvox, with strong differentiation between germ and somatic cells. In these species, the number of offspring colonies is a function of the number of germ cells, whereas the number of somatic cells is independent and much larger (Fig. 2).
Two aspects of this pattern are important. First, from the perspective of MLS2, it is important that the fitness at the colony level is now decoupled from the fitness of cells. This occurs when species begin to plot off the one-to-one line in Figure 2 in the genera Pleodorina and Volvox. The number of offspring that a Volvox colony can produce may be correlated with the size of the colony, but it is not determined by colony size alone. Second, it is not as if the somatic cells are unimportant. From an expansion point of view, the number of somatic cells is variable, and the overall size of the colonies is a function of that somatic growth. Moreover, there is more variation in somatic cell numbers among species of Volvox than there is in the number of germ cells (and therefore offspring colonies) they produce. This is a surprising pattern: colony-level reproductive rates are highest in genera such as Eudorina that fall along the one-to-one line (indicating they are in MLS1 mode). With the shift to MLS2, a drop in colony-level reproductive rates is matched with more than an order-of-magnitude increase in somatic growth rates. Evolution along these lines would not be possible in conventional thinking. But with our multilevel view of expansion, it is easy to understand that this decrease of offspring colony production is not a sacrifice of fitness because of the importance of colony size to motility and resource acquisition (Short et al. Reference Short, Solari, Ganguly, Powers, Kessler and Goldstein2006; Solari et al. Reference Solari, Ganguly, Kessler, Michod and Goldstein2006a,Reference Solari, Kessler and Michodb; Simpson Reference Simpson2021; Crockett et al. Reference Crockett, Shaw, Simpson and Kempes2023; Halling et al. Reference Halling, Goodson, Hirschmann, Wing and Simpson2024).The expansive component of fitness (represented by the frequency of somatic cells) still remains and is therefore of critical evolutionary importance as a phenotype. So it is not the germ-soma division that strictly drives a Volvox colony to evolve in an MLS2 mode; rather, an MLS2 mode occurs due to the trade-off between colony size and colony reproductive rate. To us, at least, this makes it easier to understand that the germ-soma division acts more for structuring patterns of inheritance than it does for modulating the shift from MLS1 to MLS2 (Simpson Reference Simpson, Calcott and Sterelney2011, Reference Simpson2012; Simpson et al. Reference Simpson, Herrera-Cubilla and Jackson2020).
The colonial bryozoans also plot in the region of the graph, signifying a disconnect between growth rates and reproductive rates (Fig. 3). Unfortunately there are no solitary relatives to serve as points of comparison, but data from five living Caribbean species (Jackson and Wertheimer Reference Jackson and Wertheimer1985) show that growth rates, in terms of zooids added to the colony per day, are larger than the daily rate of larval production recorded by the presence of ovicells, which are reproductive members of colonies. As with species within the volvocales, these bryozoans are clearly operating under an MLS2-type situation, with colony fitness and evolution occurring primarily at the colony level. The asexual reproduction of zooids is likewise now expressed phenotypically as growth rates of the colony.
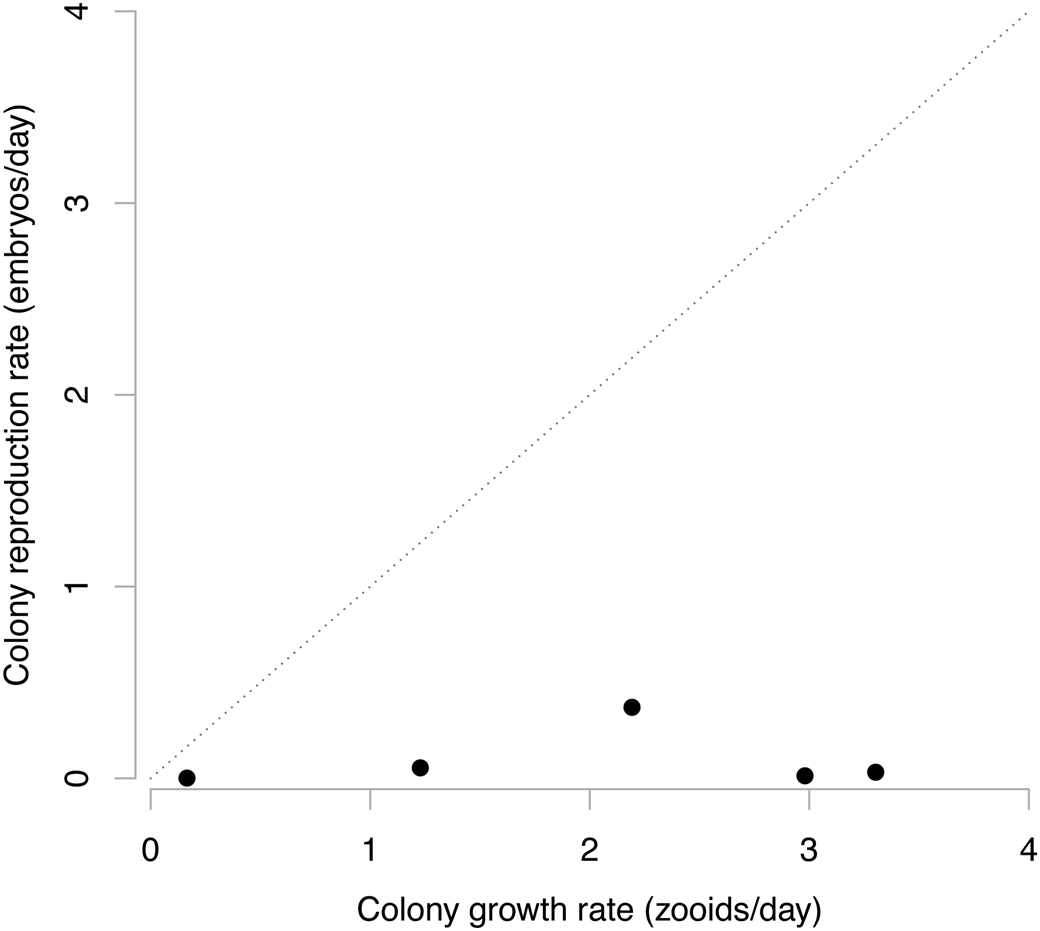
Figure 3. The zooid- and colony-level demographics of five species of cheilostome bryozoans. The diagonal one-to-one line here is similar to that in Fig. 1, where multilevel selection type 1 (MLS1) is constrained to operate. All bryozoan species plot in the off-diagonal region where multilevel selection type 2 (MLS2) dominates and maintain significant colony growth through the option of zooid birth and death rates.
Occupancy in Macroevolution
The existence of morphological stasis within species is the main piece of evidence that requires microevolution to be screened off from and independent of macroevolution (Eldredge and Gould Reference Eldredge, Gould and Schopf1972; Gould Reference Gould2002). Stasis is common in nature (Hunt Reference Hunt2007) and acts to isolate evolution within species from the macroevolutionary processes acting among species (Stanley Reference Stanley1975). Although stasis is not necessary for species selection to operate, stasis does simplify the action of species selection (Simpson Reference Simpson2017). Conceptually, stasis neutralizes within-species change, allowing among-species evolutionary processes to act. This is akin to the traditional thinking of germ-soma division of labor, which neutralizes any within-organism change and permits among-organism selection to act. Is there any pattern at this species scale that resembles expansion? Perhaps the rise and fall of geographic range size (Jernvall and Fortelius Reference Jernvall and Fortelius2004; Foote Reference Foote2007; Foote et al. Reference Foote, Crampton, Beu, Marshall, Cooper, Maxwell and Matcham2007; Liow and Stenseth Reference Liow and Stenseth2007; Liow et al. Reference Liow, Skaug, Ergon and Schweder2010) may be a species-level manifestation of expansion.
Geographic ranges are not static, but rather tend to expand and contract symmetrically over the duration of genera and species. This specific rise and fall pattern is difficult to explain by physical processes that unfold over long timescales, such as changes in habitat area or climate, because taxa at different stages in their duration are not affected by these changes in the same way. Unless all taxa are the same age when environments change extent (as could be the case with the turnover pulse hypothesis [Vrba Reference Vrba1993, Reference Vrba2005]), a drastic reduction in range size due to a regression, for example, will be experienced by old and young taxa and therefore spreads the effects of one event over the whole path of the rise and fall pattern. This does not preclude a physical control of the absolute magnitude of geographic range size or of the number of species rising or falling in range size at any point in Earth history (Spiridonov et al. Reference Spiridonov, Stankevič, Gečas, Brazauskas, Kaminskas, Musteikis, Kaveckas, Meidla, Bičkauskas and Ainsaar2020, Reference Spiridonov, Balakauskas and Lovejoy2022; Spiridonov and Eldredge Reference Spiridonov and Eldredge2024).
We do suspect that biotic interactions may have the power to limit ranges so that genera keep the geographic ranges of others at bay. Biotic interactions also vary over the duration of a taxon as it evolves and the composition of its community shifts, which may provide a mechanism for the expansion and contraction of geographic range over time. The biotic nature of the rise and fall pattern has not been demonstrated, and a likely alternative is that this is just due to averaging random walks (Foote Reference Foote2007; Foote et al. Reference Foote, Crampton, Beu, Marshall, Cooper, Maxwell and Matcham2007). Yet these rise and fall patterns predict an individual taxon's extinction (Jernvall and Fortelius Reference Jernvall and Fortelius2004; Tietje and Kiessling Reference Tietje and Kiessling2013; Kiessling and Kocsis Reference Kiessling and Kocsis2016; Žliobaitė et al. Reference Žliobaitė, Fortelius and Stenseth2017), which suggests more signal than noise in the patterns. More work is needed to test for the role of biotic interactions in determining range sizes.
If this pattern is biotic in origin, then geographic occupancy represents a quantifiable measure of microevolution within species, even if those species exhibit morphological stasis, and an opportunity to further develop a quantitative theory of how species selection and microevolution interact (Jablonski Reference Jablonski2017a,Reference Jablonskib; Simpson Reference Simpson2017) that is equivalent to the quantitative theory of organismal and colonial multilevel selection with expansion, persistence, and multiplication outlined earlier. Much like the role of sterile workers in the conventional thinking in the evolution of eusociality, stasis does not permit the contribution of microevolutionary change to macroevolutionary trends, because there is no within-species change. Instead, geographic range size evolution could well be a species-level version of expansion, because range size increases tend to be associated with the abundance of individuals of a species present in time. The main drivers of a large-scale trend may be species-level selection, but the role of species-level expansion could also be important through the covariance of geographic range size with extinction and speciation rates (Payne and Finnegan Reference Payne and Finnegan2007) and inheritance of range sizes among species (Jablonski Reference Jablonski1987; Hunt et al. Reference Hunt, Roy and Jablonski2005).
Adaptation and Multilevel Selection
The number of cell types in multicellular organisms and body types in colonial invertebrates is a convenient measure of the amount of adaptation that has taken place at the level of the group (Bell and Mooers Reference Bell and Mooers1997; McShea Reference McShea2001; McShea and Changizi Reference McShea and Changizi2003; McShea and Simpson Reference McShea, Simpson, Sterelney, Callcott and Callebaut2011; Simpson Reference Simpson2012). The majority of colonial animals possess only one body type (e.g., either polymorph types in bryozoans or caste types in eusocial insects), and the majority of multicellular organisms possess only one to seven cell types (Simpson Reference Simpson2012). When new cell or body types do evolve, reproductive division of labor always evolves first (Simpson Reference Simpson2012) and, as such, would plot off the MLS1 one-to-one line in Figures 1–3 and well within the MLS2 territory.
Simpson (Reference Simpson2012) compiled the relative frequency of reproductive and non-reproductive members and the number of body types for colonial animals. Using the bryozoan component of his dataset and data from volvocales (Koufopanou Reference Koufopanou1994), we can compare among species the number of body types (or cell types for volvocales) in a colony and the relative frequency of non-reproductive members (Fig. 4). The relative frequency of non-reproductive members measures the distance from the MLS1 one-to-one line: for MLS1 species, the frequency of non-reproductive members will be 0, but for those species far into the MLS2 area, the frequency can range from nonzero to an asymptote near 1. We find hints that the more cell types in volvocales and body types in bryozoans, the rarer reproductive members become (Fig. 4). This means that the more colony-level adaptation that occurs, the deeper into MLS2 territory (farther from the MLS1 line) the species sits. While this metric does not take into account the absolute size of the colony, it does mean that the somatic part of the colony become numerically dominant.
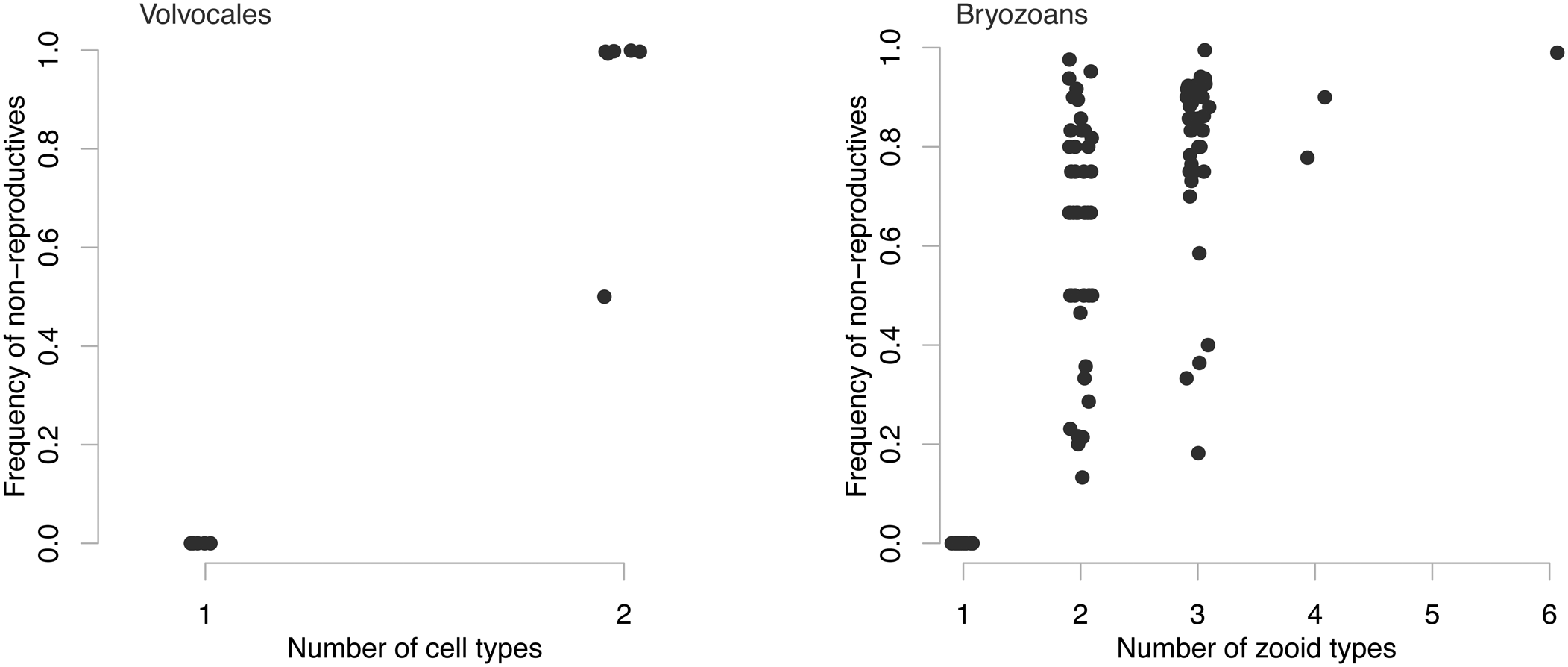
Figure 4. Patterns of group-level adaptation as a function of the relative frequency of non-reproductive colony members for both volvocine algae and bryozoans. Data from Koufopanou (Reference Koufopanou1994) and Simpson (Reference Simpson2012).
But can species selection lead to adaptation at the species level? Given that species selection is common (Simpson and Müller Reference Simpson, Müller, Müller and Asher2012), species-level traits can be heritable (Jablonski Reference Jablonski1987; Hunt et al. Reference Hunt, Roy and Jablonski2005), and extinction and speciation are unlikely to be a function of organismal fitness (Simpson Reference Simpson2016), could populations of species not evolve adaptations? Here, we suspect Gould (Reference Gould2002) is right, that the population size of species involved is too small for effective selection. Moreover, the constituent organisms of a species can and do evolve, even if stasis is a common feature (Hunt Reference Hunt2007; Hunt et al. Reference Hunt, Hopkins and Lidgard2015). In this way, species selection is different from the evolution of colonies, because in colonies, the heritability of members is neutralized (Simpson et al. Reference Simpson, Herrera-Cubilla and Jackson2020). What is then likely to occur in cases of species selection is an interaction between the species and organismal levels rather than a clearly dominant level, as is the case in colonial animals. One example of this interplay between levels is in the evolution of photosymbiosis and coloniality in scleractinian corals (Simpson Reference Simpson2013). In this two-level and multivariate case, species selection acts in opposition to the phenotypic variation determined by organismal microevolution, and the result is a non-trend where, for 200 Myr, 50% of all species are photosymbiotic.
It is easier for levels of selection to be oppositional when phenotypes are multivariate (Lande and Arnold Reference Lande and Arnold1983; Simpson Reference Simpson2013, Reference Simpson2017). Additionally, the fact that the coral photosymbiosis example is structured the way it is, because two species (host and symbiont) potentially interact at cross-purposes, suggests that other symbioses might also be prone to similar constraints on their macroevolution. If true, then constraints due to the inherently multivariate selection on species pairs may explain the relative rarity of symbiotic clades over macroevolutionary timescales. Although symbioses are extremely common across the tree of life, individual clades with symbiotic species are often species poor, as, for example, in bivalves (Vermeij Reference Vermeij2013). At the same time, our levels of selection view also highlights how special clades with ubiquitous symbionts, such as eukaryotes, are, because these transitions must have involved an evolutionary discovery that overcame the dual challenges inherent in multivariate and multilevel selection.
Fitness Is a Vector
The clean hierarchical relationship between expansion and demographic fitness among two adjacent levels makes it easy to shift conceptual focus between the part level and the level of wholes. But this view has some consequences that require consideration.
Fitness is usually assumed to be a scalar, expressible by a single number. Each organism (or other unit) has a propensity to contribute to the future only by reproducing or persisting. The usual demographic aspects of fitness, reproduction and persistence, can easily be multiplied to give the total fitness of the unit. Fitness, in the traditional scalar view, is the product of reproduction and persistence and may be modified by adding or subtracting any group selection that may occur. But growth too can influence the contribution of an evolutionary entity to the future. In most familiar organisms, say a blue jay or a puppy, this is not a reasonable approximation, because whatever growth, and therefore expansive fitness, accrues in an organism is lost when it dies. But consider a colonial organism with indeterminate growth such as a bryozoan to see how growth can be important. One large bryozoan colony can easily contain as many zooids as 100 small colonies and can even persist for many hundreds or thousands of zooid generations. Differential colonial growth can influence the frequency of traits in a population as readily as differential reproduction does because of the ecological importance of overgrowth interactions (Buss and Jackson Reference Buss and Jackson1979; McKinney and Lidgard Reference McKinney and Lidgard1992; Lidgard et al. Reference Lidgard, McKinney and Taylor1993, Reference Lidgard, Di Martino, Zágoršek and Liow2021; Jablonski et al. Reference Jablonski, Lidgard and Taylor1997; Sepkoski et al. Reference Sepkoski, McKinney and Lidgard2000; Liow et al. Reference Liow, Di Martino, Voje, Rust and Taylor2016, Reference Liow, Di Martino, Krzeminska, Ramsfjell, Rust, Taylor and Voje2017, Reference Liow, Reitan, Voje, Taylor and Di Martino2019). This example also highlights the hierarchical relationship between expansion and demographic fitness components; zooid-level demography leads to colony growth.
The hierarchical equivalence between expansion and demographic fitness is not evidence against a higher level of selection. Rather, it allows us to translate fitness at one level to fitness at the next level up in a clean and clear way. Expansion is the projection of the reproductive and persistence aspects of fitness of the parts onto the whole. In the absence of these two fitness components of the whole, the whole still has expansive fitness. But even if the whole multiplies, it may still expand. At each level, all three components of fitness, the two demographic components of persistence and multiplication and the one expansive component, are independent of each other. There may of course be interactions between components, but they are neither universal nor necessary.
This hierarchical relationship between fitness at many levels makes clear that the usual conception of fitness as a single scalar value is erroneous. The demographic processes of selection at each of multiple levels can be and often are decoupled from one another. Each level contributes its own value to a vector of fitness. Here, each element of the vector of fitnesses measures the fitness of a single level. The expansive component of fitness at one level is equal to the demographic components one level down and is recorded in the element in the vector representing fitness one level down. Van Valen (Reference Van Valen2003) argued for considering that fitness is a vector, each element corresponding to a timescale of evolution. These concepts are related to each other because, for the most part, higher levels of selection take place over longer timescales.
Conclusions
The conceptual advance of Damuth and Heisler (Reference Damuth and Heisler1988) came from recognizing the key differences between models of kin selection and altruism (in MLS1) and those for species selection (MLS2). For major transitions generally, it is likely that there is a shift from MLS1 to MLS2 (Okasha Reference Okasha2006). We see this in Figure 2 for the volvocales.
Nevertheless, how the shift from MLS1 to MLS2 occurs has been challenged by conceptual issues focused on how the cells or animals within a colony lose their fitness (Okasha Reference Okasha2006) rather than how a new level of fitness is gained. We see in Volvox and bryozoan colonies that new levels of fitness are gained once reproductive division of labor evolves (Figs. 2, 3). With expansion, Van Valen (Reference Van Valen1976, Reference Van Valen1989, Reference Van Valen1990) shows us another way to understand this shift that does not require colony members to be sterile or lose their reproductive abilities. With expansion, colony members can maintain reproductive abilities, but evolution can occur at the level of the colony, if the colony is the dominant level of heritability (Rice Reference Rice2004; Simpson Reference Simpson2012; Simpson et al. Reference Simpson, Herrera-Cubilla and Jackson2020; Leventhal et al. Reference Leventhal, Jamison-Todd and Simpson2023).
Similarly, species selection and related phenomena such as the effect hypothesis (Vrba Reference Vrba1980) or species sorting (Vrba and Gould Reference Vrba and Gould1986) are mired in conceptual issues that echo those that surround the major transitions. Species selection and the heritability of species-level traits seem to have all the ingredients to produce macroevolutionary trends. Yet trends are relatively rare given how common differential rates of speciation and extinction are—even for traits such as geographic range that are known to have some heritability (Jablonski Reference Jablonski1987; Hunt et al. Reference Hunt, Roy and Jablonski2005). We suspect that this pattern among macroevolutionary trends mirrors the frequencies of major transitions that unambiguously result in complex multicellularity or eusociality. The clearest patterns of higher levels of selection are those in which there is no ambiguity in what levels are acting; in nature, those situations may, in fact, be rare. Moreover, once the spatial or temporal scale of the clade of species becomes large enough, interactions and competition among species’ populations, termed “avatars” by Damuth (Reference Damuth1985), may start to dominate and erode any directional trends in a way similar to the variable magnitude and direction of natural selection commonly observed in nature (e.g., Grant and Grant Reference Grant and Grant2002). Large spatial and temporal scales may also pool variable geographic and environmental contexts leading to geological-scale processes playing a role in controlling diversity dynamics (Spiridonov et al. Reference Spiridonov, Stankevič, Gečas, Brazauskas, Kaminskas, Musteikis, Kaveckas, Meidla, Bičkauskas and Ainsaar2020, Reference Spiridonov, Balakauskas and Lovejoy2022).
But expansion, and its empirical utility in cleanly identifying the hierarchical nature of levels (expansion at one level is produced by differential reproduction and survival of the level below), allows us to study the rich variety of cases in which organisms live at the interface of two levels. Bryozoans serve as a wonderfully rich example of living at the interface of the zooid and colony levels. The life-history aspects of growth and sexual reproduction vary dramatically among species (McKinney and Jackson Reference McKinney and Jackson1991; Herrera et al. Reference Herrera, Jackson, Gordon, Smith and Grant-Mackie1996a,Reference Herrera, Jackson, Hughes, Jara and Ramosb) as do colony-level phenotypes such as polymorphism (Hughes and Jackson Reference Hughes and Jackson1990; Simpson et al. Reference Simpson, Jackson and Herrera-Cubilla2017; Schack et al. Reference Schack, Gordon and Ryan2018, Reference Schack, Gordon and Ryan2020).
With expansion in our intellectual and quantitative tool kit, we are likely to see that bacterial multicellularity is not so simple. How does the growth of bacterial colonies compare to the rate of formation of new colonies? If group-living bacteria form new colonies, why do they not evolve more complex forms? One reason may be that the structure of bacterial genomes tends to lose genes, and group living, along with the reduction in effective population size that accompanies it, increases the rate of gene loss though drift (Bingham and Ratcliff Reference Bingham and Ratcliff2024). Bingham and Ratcliff's (Reference Bingham and Ratcliff2024) insight that colony structure necessarily reduces effective population size matches the patterns we see in Figures 2 and 3 for eukaryotic unicellular and animal colonies. For bacteria, the process leads to a more erosion-prone genome, but for eukaryotes, this reduction in effective population sizes leads to more effective genomic gains (Bingham and Ratcliff Reference Bingham and Ratcliff2024).
Expansion is a simple tool, but even a cursory application of it can yield insights. Of all the examples of transitions to multicellularity and colonial or social living in animals, only eusocial Hymenoptera represent the conventional case wherein colonies reproduce, while the majority of members are sterile. There is a richness in every other example we know of: examples where cells divide and organisms reproduce and the balance of what level dominates evolution can be tipped by patterns of heredity. Cases in which ancestral levels of fitness remain, but are channeled into development. We are sure we will find lots of new multilevel biology now that we know how to look for it.
Acknowledgments
C.S thanks Doug Erwin, Dan McShea, Jeremy Jackson, Wolfgang Kiessling, and Leigh Van Valen for discussions about expansion over the years. Thanks to Dan McShea, Mark Patzkowsky, and Andrej Spiridonov for helpful and insightful reviews. This paper is dedicated to John Damuth, who first walked the path we follow in this paper. A.H. was supported by NSF GRFP and S.E.L. by a NMNH predoctoral fellowship, Richard K Bambach Student Research Award from the Paleontological Society, and the Benson Fellowship.
Competing Interests
The authors declare no competing interests.





 Open access
Open access