



Introduction
Parasitism is a highly successful lifestyle and has evolved independently at least 60 times in different groups of metazoans worldwide (Price, Reference Price1980; Poulin and Morand, Reference Poulin and Morand2004). Parasites have been traditionally divided into 2 major groups depending on their life cycle: generalists and specialists (Thompson, Reference Thompson1994, Reference Thompson2005). The generalist parasites use a wide range of definitive hosts, whereas the specialist parasites use a single definitive host to complete their life cycle. Under these 2 opposing strategies, generalist parasites infect a broad spectrum of hosts resulting in an optimal or suboptimal level of fitness, whereas specialist parasites prioritize a single optimal host in which fitness is maximized (Rigaud et al., Reference Rigaud, Perrot-Minnot and Brown2010; Lievens et al., Reference Lievens, Perreau, Agnew, Michalakis and Lenormand2018). Some studies have suggested that parasite life cycle complexity (generalists vs specialists) could influence population genetic structure (Nadler, Reference Nadler1995; Criscione and Blouin, Reference Criscione and Blouin2004; Barrett et al., Reference Barrett, Thrall, Burdon and Linde2008; Archie and Ezenwa, Reference Archie and Ezenwa2011). According to Li et al. (Reference Li, Jovelin, Yoshiga, Tanaka and Cutter2014), a specialist parasite shows significantly less genetic flow; therefore, populations are less connected and are subdivided into smaller populations, leading to strong genetic differentiation. In some cases, the populations might experience bottlenecks, decreasing the effective population size. Moreover, specialist species are more sensitive to stochastic fluctuations that can cause local extinction. In contrast, a generalist parasite shows a high effective population size, high genetic flow and a population that is structured or panmictic. Moreover, a generalist parasite may show greater persistence of populations over the long term because the generalist may be less sensitive to stochastic fluctuations in any given resource as it is able to replace a scarce resource with another (see Sehgal et al., Reference Sehgal, Jones and Smith2001; Brant and Ortí, Reference Brant and Ortí2003; Archie and Ezenwa, Reference Archie and Ezenwa2011; Li et al., Reference Li, Jovelin, Yoshiga, Tanaka and Cutter2014).
The recent application of molecular markers has helped establishing a more robust classification scheme in acanthocephalans (Near et al., Reference Near, Garey and Nadler1998; García-Varela et al., Reference García-Varela, Pérez-Ponce de León, de la Torre, Cummings, Sarma and Laclette2000). In particular, cytochrome c oxidase subunit 1 (cox1) from mitochondrial DNA is among the most useful molecular markers for defining, recognizing and delineating species and better understanding the population genetic structure in acanthocephalans (Steinauer et al., Reference Steinauer, Nickol and Ortí2007; Rosas-Valdez et al., Reference Rosas-Valdez, Morrone and García-Varela2012, Reference Rosas-Valdez, Morrone, Pinacho-Pinacho, Domínguez-Domínguez and García-Varela2020; Alcántar-Escalera et al., Reference Alcántar-Escalera, García-Varela, Vázquez-Domínguez and Pérez Ponce de León2013; Goulding and Cohen, Reference Goulding and Cohen2014; Perrot-Minnot et al., Reference Perrot-Minnot, Špakulová, Wattier, Kotlík, Düsen, Ayogdu and Tougard2018; Pinacho-Pinacho et al., Reference Pinacho-Pinacho, García-Varela, Sereno-Uribe and Pérez Ponce de León2018; García-Varela et al., Reference García-Varela, Masper, Crespo and Hérnandez-Orts2021; Sereno-Uribe et al., Reference Sereno-Uribe, López-Jiménez, González-García, Pinacho-Pinacho, Macip Ríos and García-Varela2022). Polymorphidae (Meyer, 1931) is an emblematic group of obligate endoparasites with complex life cycles that use vertebrates (marine mammals, fish-eating birds and waterfowl) as definitive hosts and invertebrates (amphipods, decapods and euphausiids) as intermediate hosts to complete their life cycle (Schmidt, Reference Schmidt, Nickol and Crompton1985; Hoberg, Reference Hoberg1986; Pichelin et al., Reference Pichelin, Kuris and Gurney1998; Nickol et al., Reference Nickol, Crompton and Searle1999, Reference Nickol, Heard and Smith2002; Kennedy, Reference Kennedy2006). Currently, the family is classified into 14 genera with approximately 129 species (Schmidt, Reference Schmidt1973; Dimitrova and Georgiev, Reference Dimitrova and Georgiev1994; Nickol et al., Reference Nickol, Crompton and Searle1999, Reference Nickol, Heard and Smith2002; Aznar et al., Reference Aznar, Pérez Ponce de León and Raga2006; Amin, Reference Amin2013; García-Varela et al., Reference García-Varela, Pérez-Ponce de León, Aznar and Nadler2013b; Presswell et al., Reference Presswell, Bennett and Smales2020). Phylogenetic analyses based on multiple molecular markers have suggested that the family is monophyletic as are the genera Hexaglandula (Petrochenko, 1950) and Southwellina (Witenberg, 1932) (see García-Varela et al., Reference García-Varela, Pérez-Ponce de León, Aznar and Nadler2011, Reference García-Varela, Pérez-Ponce de León, Aznar and Nadler2013b).
Members of the Polymorphidae form a monophyletic group that contains generalist and specialist species. Therefore, this family represents an interesting system to explore and compare the population genetic structure of species with 2 opposing life history strategies. Hexaglandula corynosoma (Travassos, Reference Travassos1915) is a specialist species that has been recorded as adult only in the intestine of the yellow-crowned night-heron (Nyctanassa violacea) (Linnaeus, 1758), and cystacanths have been recorded in 2 decapod species [the fiddler crabs Leptuca spinicarpa (Rathbun) and Minuca rapax (Smith)], which serve as intermediate hosts in the Americas (Nickol et al., Reference Nickol, Heard and Smith2002; Guillén-Hernández et al., Reference Guillén-Hernández, García-Varela and Pérez-Ponce de León2008; García-Prieto et al., Reference García-Prieto, García-Varela, Mendoza-Garfias and Pérez-Ponce de León2010). In contrast, the acanthocephalan Southwellina hispida (Van Cleave, Reference Van Cleave1925) Witenberg, 1932 is a generalist species and is considered one of the most abundant species of polymorphids associated with piscivorous birds throughout the world (see Amin et al., Reference Amin, Chaudhary and Singh2022). In Mexico, S. hispida has been documented in 13 piscivorous bird species (García-Prieto et al., Reference García-Prieto, García-Varela, Mendoza-Garfias and Pérez-Ponce de León2010; García-Varela et al., Reference García-Varela, Aznar, Rodríguez and Pérez-Ponce de León2012), and the cystacanth has been recorded in the red swamp crayfish Procambarus clarkii (Girard) in the USA (Font, Reference Font2007), which serves as an intermediate host that is ingested by several vertebrates that act as paratenic hosts such as frogs and freshwater and brackish fishes (Schmidt, Reference Schmidt, Nickol and Crompton1985). The paratenic hosts must be ingested by piscivorous birds to complete their life cycle (Schmidt, Reference Schmidt, Nickol and Crompton1985).
In the current study, we examined the sequences of cox1 from mitochondrial DNA of 2 closely related polymorphid species, a specialist species (H. corynosoma) and a generalist species (S. hispida), distributed sympatrically in 6 biogeographic provinces (California, Baja California, Pacific Lowlands, Tamaulipas, Veracruzan and the Yucatán Peninsula) of Mexico with the objective of characterizing and comparing the population genetic structure of 2 acanthocephalan species with opposing life history strategies.
Materials and methods
Specimen collection
A total of 75 birds, and 36 fishes, were collected between October 2006 and December 2021 in 23 localities across 6 biogeographic provinces (California, Baja California, Pacific Lowlands, Tamaulipas, Veracruzan and the Yucatán Peninsula) of Mexico (Table 1; Fig. 1). Birds and fishes were dissected within the following 4 h, and their viscera were placed in separate Petri dishes containing a 0.75% saline solution and examined under a dissecting microscope. The acanthocephalans recovered were washed in 0.75% saline solution and placed overnight in distilled water at 4°C and subsequently preserved in 100% ethanol. Birds and fishes were identified using the field guide of Howell and Webb (Reference Howell and Webb1995) and Miller et al. (Reference Miller, Minckley and Norris2005), respectively.
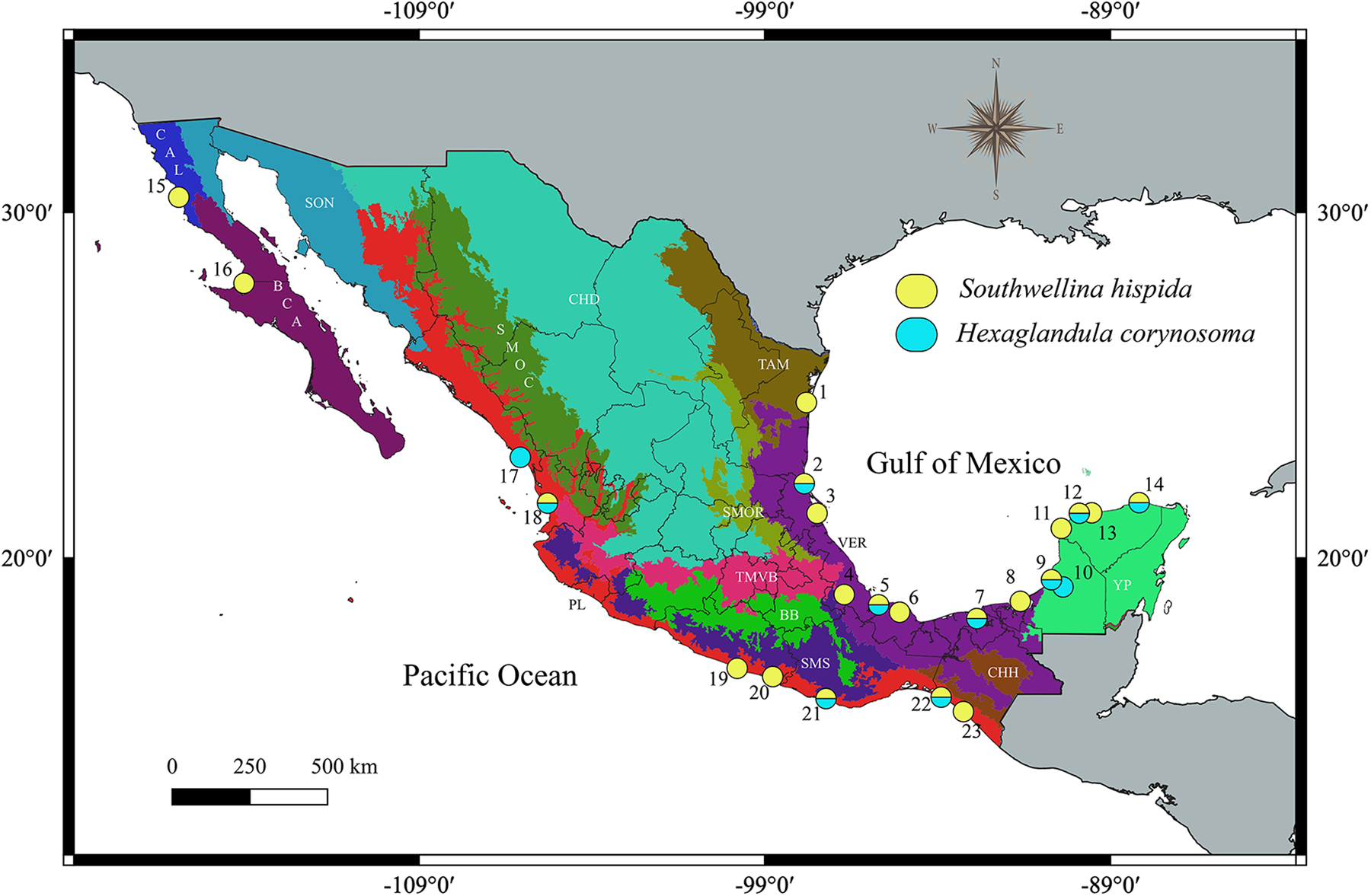
Fig. 1. Map of Mexico showing the sampled sites for the birds. Localities with a circle of yellow and turquoise colour were positive for the infection with Southwellina hispida and Hexaglandula corynosoma, respectively; localities correspond to those in Table 1.
Table 1. Specimens’ information, collection sites (CS), host name; number of host examined/infected (prevalence of infection); number of specimens analysed (n); locality, geographical coordinates, GenBank accession number for specimens studied in this study

The sample number for each locality corresponds with the same number in Fig. 1. Sequences in bold were generated in the current study.
Morphological analyses
Selected adult acanthocephalans were gently punctured with a fine needle in the trunk, stained with Mayer's paracarmine, destained in 70% acid ethanol, dehydrated in a graded ethanol series, cleared in methyl salicylate and mounted in Canada balsam. Specimens were examined using a compound microscope Leica DM 1000 LED equipped with bright field (Leica, Wetzlar, Germany). The acanthocephalans were identified by conventional morphological criteria following Petrochenko (Reference Petrochenko1958). In addition, descriptions of H. corynosoma and S. hispida were consulted as needed (Schmidt, Reference Schmidt1973; Nickol et al., Reference Nickol, Heard and Smith2002; Amin et al., Reference Amin, Chaudhary and Singh2022). For scanning electron microscopy, 2 adult specimens of each species were dehydrated with an ethanol series, critical point dried, sputter coated with gold and examined with a Hitachi Stereoscan Model S-2469N scanning electron microscope operating at 15 kV from the Instituto de Biología, Universidad Nacional Autónoma de México (UNAM). Adult specimens were deposited in the Colección Nacional de Helmintos, under numbers CNHE: 11823 and 11824, Instituto de Biología, Universidad Nacional Autónoma de México, Mexico City
DNA isolation, amplification and sequencing
A total of 97 specimens, 41 identified as H. corynosoma and 56 as S. hispida were placed individually in tubes and digested overnight at 56°C in a solution containing 10 mm Tris–HCl (pH 7.6), 20 mm NaCl, 100 mm Na2-EDTA (pH 8.0), 1% Sarkosyl and 0.1 mg mL−1 proteinase K. Following digestion, DNA was extracted from the supernatant using the DNAzol reagent (Molecular Research Center, Cincinnati, Ohio, USA) according to the manufacturer's instructions. The cox1 of the mitochondrial DNA was amplified using the forward primer 5′-AGTTCTAATCATAA(R)GATAT(Y)GG-3′ and reverse primer 5′-TAAACTTCAGGGTGACCAAAAAATCA-3′ (Folmer et al., Reference Folmer, Black, Hoeh, Lutz and Vrijenhoek1994). Polymerase chain reactions (PCRs) (25 μL) consisted of 10 μL of each primer, 2.5 μL of 10× buffer, 2 mm MgCl2 and 1 U of Taq DNA polymerase (Platinum Taq, Invitrogen Corporation, São Paulo, Brazil). PCR cycling parameters for the molecular marker consisted of denaturation at 94°C for 1 min, 35 cycles of 94°C for 1 min, 40°C for 1 min and 72°C for 1 min, followed by a post-amplification incubation at 72°C for 10 min. Sequencing reactions were performed using ABI Big Dye (Applied Biosystems, Boston, Massachusetts, USA) terminator sequencing chemistry, and reaction products were separated and detected using an ABI 3730 capillary DNA sequencer. Contigs were assembled and base-calling differences resolved using Codoncode Aligner version 9.0.1 (Codoncode Corporation, Dedham, Massachusetts, USA) and submitted to the GenBank dataset (Table 1).
Alignments, population genetic structure and historical demographic
Newly obtained sequences in the current research of S. hispida were aligned with 22 other sequences of S. hispida (EF467867–868, HM636468–469, FJ824172–189) downloaded from GenBank (Table 1), forming a dataset of 78 sequences with 646 characters, and the new sequences of H. corynosoma were aligned with other sequences of H. corynosoma (EU189485–486, EU189488 and EF467869), downloaded from GenBank (Table 1), forming a dataset of 46 sequences with 644 characters. Sequences of each dataset were aligned separately using software ClustalW with default parameters implemented in MEGA version 7.0 (Kumar et al., Reference Kumar, Stecher and Tamura2016).
To analyse the molecular information in the framework of population genetics, we grouped individuals of H. corynosoma and S. hispida into populations considering the biogeographic provinces (California, Baja California, Pacific Lowlands, Tamaulipas, Veracruzan and the Yucatán Peninsula). Intrapopulation variation was summarized using standard statistics: number of haplotypes (H), number of segregating sites (S), haplotype diversity (H d), nucleotide diversity (P i) and average number of nucleotide differences (K), were all calculated using the program DnaSP v.5.10 (Rozas et al., Reference Rozas, Sánchez-DelBarrio, Messeguer and Rozas2003). To examine haplotype frequency among the populations of H. corynosoma and S. hispida a statistical network was constructed independently, using the program PopART with the median joining algorithm (Bandelt et al., Reference Bandelt, Forster and Röhl1999). The degree of genetic differentiation among the populations was estimated using the fixation indices F st (Hudson et al., Reference Hudson, Boos and Kaplan1992), with the program Arlequin v.3.5 (Excoffier and Lischer, Reference Excoffier and Lischer2010). To investigate the genetic variation among populations or within populations, the analysis of molecular variance was performed, considering genetic distance among the haplotypes using Arlequin v.3.5. To investigate the population history and demography, Tajima's D (Tajima, Reference Tajima1989) and Fu's F s (Fu, Reference Fu1997) were calculated using DnaSP v.5.10 (Rozas et al., Reference Rozas, Sánchez-DelBarrio, Messeguer and Rozas2003). The values were considered significant when the P values were less than 0.05.
Results
Morphological identification
The acanthocephalans recovered from diverse definitive hosts such as herons, gulls, cormorants, pelicans and hawks on both coasts of Mexico show similar morphological characteristics compared with those assigned to S. hispida by García-Varela et al. (Reference García-Varela, Aznar, Rodríguez and Pérez-Ponce de León2012) and Amin et al. (Reference Amin, Chaudhary and Singh2022), including (i) an elongated cylindrical trunk with 2 fields of somatic spines on the anterior region of the trunk, (ii) a cylindrical proboscis with a swollen region, (iii) proboscis hooks arranged in 16–17 longitudinal rows of 12–15 hooks per row, (iv) a double-walled proboscis receptacle and (v) 4 tubular cement glands in males (Fig. 2A–E). The acanthocephalans recovered from the intestine of the yellow-crowned night-heron (N. violacea) in coastal Mexico show morphological characteristics that match those assigned to H. corynosoma by Nickol et al. (Reference Nickol, Heard and Smith2002), Guillén-Hernández et al. (Reference Guillén-Hernández, García-Varela and Pérez-Ponce de León2008) and Amin et al. (Reference Amin, Chaudhary and Singh2022), including (i) an elongate cylindrical trunk, swollen anteriorly with a single field of somatic spines in the anterior region of the trunk; (ii) a cylindrical proboscis; (iii) proboscis hooks arranged in 16 longitudinal rows of 11–12 hooks per row; (iv) a double-walled proboscis receptacle and (v) 6 tubular cement glands in males (Fig. 3A–E).


Population genetic structure and demographic analysis
The mitochondrial marker was successfully amplified for 56 S. hispida individuals and 41 H. corynosoma individuals. The complete alignment of the cox1 dataset contained 78 S. hispida individuals with a total length of 646 bp, whereas the cox1 dataset of H. corynosoma contained 46 individuals with a total length of 644 bp. No insertions or deletions were detected in any of the sequences, and when the sequences were translated into proteins, no stop codons were found.
The haplotype network built for the generalist species S. hispida did not show a phylogeographic structure; in fact, of the 55 mtDNA haplotypes detected, 50 were unique (singlets), and 5 others were shared haplotypes (H4, H11, H17, H29 and H30). The most frequent haplotype (H4, n = 12) corresponded to specimens from 4 biogeographic provinces (Pacific Lowlands, Baja California, Veracruzan and the Yucatán Peninsula). Most of the identified haplotypes were separated from one another by 1, 2, 3 or up to 5 substitutions (see Fig. 4). The haplotype diversity was very high (H d = 0.970) and nucleotide diversity was low (p i = 0.00555) among the populations from the 4 biogeographic provinces sampled (Pacific Lowlands, Baja California, Veracruzan and the Yucatán Peninsula) of Mexico. Neutrality tests (Tajima's D, −2.473 and Fu's F s, −34.252) were negative for all regions (see Table 2), indicating an excess of rare alleles greater than what would be expected under neutrality and suggesting a recent population expansion of S. hispida. The F st values were estimated to assess genetic differentiation among populations from the 4 biogeographic provinces analysed (Pacific Lowlands, Baja California, Veracruzan and the Yucatán Peninsula). Despite the large geographic distances, the F st values were low, ranging from −0.007 to 0.058 (Table 3), which indicates that the populations were poorly genetically differentiated from one another. The sampled definitive and paratenic host species of S. hispida were grouped into 8 families (Fig. 5). The host haplotype network did not show a pattern; 30 haplotypes were found in herons scattered throughout the network. In the paratenic hosts sampled (cichlid fishes), 11 haplotypes were detected (H3, H6, H7, H8, H10, H17, H30, H32, H33, H34 and H35), suggesting that the paratenic hosts are able to harbour and transmit diverse haplotypes to definitive hosts (Fig. 5).

Fig. 4. Haplotype network of samples of S. hispida, built with the gene cox1 from mitochondrial DNA. Each circle represents a haplotype, with size proportional to the haplotype's frequency in the populations. Mutational steps are symbolized by dashes. Biogeographic provinces: Veracruzan (VER); Yucatán Peninsula (YUC); Pacific Lowlands (PLN); Tamaulipas (TAM); California (CAL) and Baja California (BCA).

Fig. 5. Host haplotype network of samples of S. hispida, built with the gene cox1 from mitochondrial DNA. The paratenic and definitive hosts were grouped by families. Each circle represents a haplotype, with size proportional to the haplotype's frequency in the populations. Mutational steps are symbolized by dashes.
Table 2. Molecular diversity indices and neutrality tests calculated for cox1 datasets among the populations of S. hispida used in this study

n, number of sequences; H, number of haplotypes; S, number of segregating sites; H d, haplotype diversity; P i, nucleotide diversity; K, average number of nucleotide differences.
Table 3. Pairwise F st values estimated for cox1

VER, Veracruzan; YP, Yucatan Peninsula; BCAL, Baja California; PL, Pacific Lowlands.
F st average among 3 biogeographic provinces = 0.03672.
Significance level = 0.05.
The haplotype network of the specialist species H. corynosoma did not show a phylogeographic structure for the 42 mtDNA haplotypes detected; 41 were unique (singlets), and only 1 was shared (H2, n = 4) between 2 biogeographic provinces (Veracruzan and the Yucatán Peninsula). Most of the identified haplotypes were separated by several substitutions (see Fig. 6). The haplotype diversity (H d = 0.993) was very high and nucleotide diversity was low (p i = 0.01166) among the populations from the 3 biogeographic provinces sampled (Pacific Lowlands, Veracruzan and the Yucatán Peninsula). Neutrality tests (Tajima's D, −1.875 and Fu's F s, −33.964) were negative for all regions (see Table 4), which indicates an excess of rare alleles greater than what would be expected under neutrality, suggesting a recent population expansion of H. corynosoma. The F st values were estimated to assess genetic differentiation among the populations from the 3 biogeographic provinces analysed (Pacific Lowlands, Veracruzan and the Yucatán Peninsula). Despite the large geographic distances, the F st values were low, ranging from −0.019 to 0.059 (Table 5), which indicates that the populations were poorly genetically differentiated from one another.

Fig. 6. Haplotype network of samples of H. corynosoma, built with the gene cox1 from mitochondrial DNA. Each circle represents a haplotype, with size proportional to the haplotype's frequency in the populations. Mutational steps are symbolized by dashes. Biogeographic provinces: Veracruzan (VER); Yucatán Peninsula (YUC) and Pacific Lowlands (PLN).
Table 4. Molecular diversity indices and neutrality tests calculated for cox1 datasets among the populations of H. corynosoma used in this study

n, number of sequences; H, number of haplotypes; S, number of segregating sites; H d, haplotype diversity; P i, nucleotide diversity; K, average number of nucleotide differences.
Table 5. Pairwise F st values estimated for cox

VER, Veracruzan; YP, Yucatan Peninsula; PL, Pacific Lowlands
F st average among 3 biogeographic provinces = 0.05609.
Significance level = 0.05.
Discussion
To the best of our knowledge, S. hispida and H. corynosoma are 2 species that use piscivorous birds as definitive hosts and decapods as intermediate hosts, share a common ancestor within Polymorphidae and are distributed sympatrically along the coastline of Mexico (Guillén-Hernández et al., Reference Guillén-Hernández, García-Varela and Pérez-Ponce de León2008; García-Prieto et al., Reference García-Prieto, García-Varela, Mendoza-Garfias and Pérez-Ponce de León2010; García-Varela et al., Reference García-Varela, Pérez-Ponce de León, Aznar and Nadler2013b). The generalist species S. hispida was described in Japan by Van Cleave in Reference Van Cleave1925, and since then it has been recorded in the body cavities of fishes and reptiles as a cystacanth (larval stage) and as an adult form in the intestines of diverse piscivorous birds in Asia, Europe and the Americas (see Amin et al., Reference Amin, Chaudhary and Singh2022). In Mexico, adults of S. hispida have been recorded in 15 piscivorous bird species (García-Prieto et al., Reference García-Prieto, García-Varela, Mendoza-Garfias and Pérez-Ponce de León2010; García-Varela et al., Reference García-Varela, Aznar, Rodríguez and Pérez-Ponce de León2012). It is well known that helminths with a broad spectrum of definitive hosts show phenotypic plasticity in their morphological traits. Schmidt (Reference Schmidt1973), García-Varela et al. (Reference García-Varela, Aznar, Rodríguez and Pérez-Ponce de León2012) and Amin et al. (Reference Amin, Chaudhary and Singh2022) documented the phenotypic plasticity of S. hispida recovered from diverse definitive hosts including in traits such as body size, leminisci, proboscis receptacle, testes, cement glands and the reproductive system in females. However, diagnostic characters such as proboscis shape, number of proboscis hooks and the presence of 2 fields of spines on the anterior trunk region in both sexes did not vary among specimens, including our specimens collected from the coasts of the Gulf of Mexico and the Pacific Ocean slopes (see Fig. 2A–E). In Brazil, the specialist species H. corynosoma was described as parasite of the yellow-crowned night heron (N. violacea) by Travassos in Reference Travassos1915. Since then, H. corynosoma has been recorded in Puerto Rico, the USA and Mexico in the same definitive host species (Cable and Quick, Reference Cable and Quick1954; Nickol et al., Reference Nickol, Heard and Smith2002; Guillén-Hernández et al., Reference Guillén-Hernández, García-Varela and Pérez-Ponce de León2008). Our specimens identified as H. corynosoma agree morphologically with those previously described by Nickol et al. (Reference Nickol, Heard and Smith2002) and Guillén-Hernández et al. (Reference Guillén-Hernández, García-Varela and Pérez-Ponce de León2008). For example, an elongated trunk, swollen anteriorly; covered with spines on the anterior region of the trunk; a cylindrical proboscis with 16 longitudinal rows of 11–12 hooks each; a conical neck; ovoid testes located in the swollen portion of the trunk and long, tubular cement glands (see Fig. 3A–E).
The intraspecific genetic divergence estimated in the current study among the 78 isolates of S. hispida and the 46 isolates of H. corynosoma ranged from 0.00 to 1.5% and from 0.00 to 2.6%, respectively. These values of intraspecific genetic divergence are similar to those previously reported for isolates of polymorphid species such as Andracantha sigma (Presswell et al., Reference Presswell, García-Varela and Smales2017) recovered from 3 definitive hosts, the Otago shag, Leucocarbo chalconotus (Gray), spotted shag Phalacrocorax punctatus (Sparrman) and Otago little blue penguin, Eudyptula novaehollandiae (Forster) from New Zealand, which ranged from 0.00 to 0.32% (Presswell et al., Reference Presswell, García-Varela and Smales2017); similar to those among 14 adults, 3 acanthella and 4 cystacanths of Pseudocorynosoma constrictum (Van Cleave, 1918) Aznar et al. (Reference Aznar, Pérez Ponce de León and Raga2006) recovered from 7 wild duck species and the freshwater amphipod Hyalella azteca (Saussure) in central Mexico, which ranged from 0.0 to 3.0% (García-Varela et al., Reference García-Varela, Pinacho-Pinacho, Sereno-Uribe and Mendoza-Garfías2013a) and similar to those among 19 adults recovered from 3 fish-eating bird species and 33 cystacanths recovered from 19 freshwater fish species identified as Polymorphus brevis (Van Cleave, 1916) Travassos, 1926, which ranged from 0.00 to 1.6% (Alcántar-Escalera et al., Reference Alcántar-Escalera, García-Varela, Vázquez-Domínguez and Pérez Ponce de León2013). Furthermore, the values from this study were also similar to those among adults and cystacanths of Corynosoma hannae Zdzitowiecki, 1984 recovered from the New Zealand sea lion (Phocarctos hookeri Grey), Stewart Island shag (L. chalconotus Gray), spotted shags (P. punctatus Sparrman), yellow-eyed penguins (Megadyptes antipodes Hombron and Jacquinot), New Zealand brill (Colistium guntheri Hutton) and New Zealand sole (Peltorhamphus novaezeelandiae Gunther), which ranged from 0.00 to 2.8% (Hernández-Orts et al., Reference Hernández-Orts, Brandão, Georgieva, Raga, Crespo, Luque and Aznar2017) and those among Corynosoma australe (Johnston, 1937) recovered from the California sea lion (Zalophus californianus Lesson), South American sea lions (Otaria flavescens Shaw), South American fur seals (Arctocephalus australis Zimmermann), Magellanic penguins (Spheniscus magellanicus Forster) and cystacanths recovered from marine fishes in Argentina, which ranged from 1.0 to 1.7% (García-Varela et al., Reference García-Varela, Masper, Crespo and Hérnandez-Orts2021).
The haplotype network genealogy generated in this study based on cox1 sequences from S. hispida (generalist) and H. corynosoma (specialist) did not show a phylogeographic structure; therefore, the haplotypes could not be grouped into their own geographic clusters. In fact, the specimens of S. hispida were not correlated with their definitive host family (see Figs 4 and 5). The populations analysed for both species of acanthocephalans were classified into biogeographical provinces separated by geographical barriers as follows: mountains, the dry lowlands of the Isthmus of Tehuantepec, the Balsas Depression and the central Trans-Mexicana Volcanic Belt (Barrier et al., Reference Barrier, Velasquillo, Chavez and Gaulon1998; Ferrari et al., Reference Ferrari, Orozco-Esquivel, Manea and Manea2012; Morrone et al., Reference Morrone, Escalante and Rodríguez-Tapia2017). Despite the large geographic distances, the F st values estimated among the populations of both species were very low (Tables 3 and 5), indicating that the populations were poorly genetically differentiated from each other; this can be explained by the migration patterns of birds along the coasts of Mexico. Historical events leave signatures in the DNA, and neutrality tests can infer the demographic history of populations. In both species analysed (generalist and specialist), the estimated values of Fu's F s and Tajima's D among the populations were negative (see Tables 2 and 4). In addition, high haplotype diversity and an excess of low-frequency haplotypes were detected in both networks (Figs 4 and 6), although the haplotypes differed from one another by fewer than 5 nucleotide substitutions. These findings confirmed that both populations of acanthocephalans had experienced rapid population growth in the past.
In this study, we found that 2 species of acanthocephalans with opposing life history strategies (generalist and specialist) showed similar population genetics patterns. This pattern was not consistent with the specialist–generalist variation hypothesis (SGVH), which predicts that: (i) populations of specialists may be less connected and more subdivided into smaller populations than generalists; (ii) specialists are expected to have lower effective population sizes than generalists and to be composed of populations with less gene flow and (iii) generalists will show high effective population sizes, high genetic flow and highly structured populations (see Dennis et al., Reference Dennis, Dapporto, Fattorini and Cook2011; Li et al., Reference Li, Jovelin, Yoshiga, Tanaka and Cutter2014). According to the SGVH, the populations of S. hispida (generalist species) resemble the specialist model. To date, only a few phylogeographic studies of generalist acanthocephalans have been conducted using cox1 as a molecular marker. For example, Profilicollis altmani (Perry, 1942), which has a broad distribution across North and South America and parasitizes multiple species of intermediate hosts and diverse species of marine birds that act as definitive hosts (gulls, ducks, sanderling and common tern), showed a lack of population genetic structure with high haplotype diversity and low nucleotide diversity, suggesting that P. altmani have experienced a period of rapid population growth in the past (see Goulding and Cohen, Reference Goulding and Cohen2014). Similarly, Profilicollis novaezelandensis Brockerhoff and Smales (Reference Brockerhoff and Smales2002) is a parasite that has been recorded as an adult in gulls (Larus spp.) and oystercatchers (Haematopus spp.) and as cystacanths in the shore crab Hemigrapsus crenulatus (Milne-Edwards) distributed along the east coast of New Zealand's South Island (Brockerhoff and Smales, Reference Brockerhoff and Smales2002); population genetic analyses from 50 P. novaezelandensis individuals from 8 localities showed a lack of population genetic structure with high haplotype diversity and low nucleotide diversity (Hay et al., Reference Hay, Jorge and Poulin2018).
The 2 species of acanthocephalans analysed here are considered to be typical components of the helminth fauna of piscivorous birds in the Americas. Of the 19 piscivorous birds recorded as definitive hosts, 11 belong to the family Ardeidae, which represent 58% of the host diversity, suggesting that the ardeids could be the ancestral hosts of S. hispida and H. corynosoma with secondary and independent colonization events to other piscivorous birds. In particular, S. hispida was recorded in diverse piscivorous birds such as pelicans, cormorants, eagles, hawks, anhingas and royal terns with a lower level of infection (prevalence values <20%) than those of ardeid hosts (prevalence values from 80 to 100%), suggesting that these piscivorous birds may act as suboptimal definitive hosts.
The population genetic structures of S. hispida and H. corynosoma showed similar patterns. However, these species use different strategies to complete their life cycles. The main ecological difference between S. hispida and H. corynosoma is the inclusion of paratenic hosts in S. hispida (fishes from the families Cichlidae, Eleotridae, Characidae, Lutjanidae, Centropomidae, Sparidae, Scianidae and Paralichthyidae) (García-Prieto et al., Reference García-Prieto, García-Varela, Mendoza-Garfias and Pérez-Ponce de León2010). The paratenic hosts act as a trophic bridge and facilitate the transmission between the intermediate host and definitive host, but apparently do not play a central role in the population genetic structure of S. hispida within its distribution along the coasts of Mexico.
Acknowledgements
We thank Laura Márquez and Nelly López Ortiz from LaNabio for their help during the sequencing of the DNA fragments. We also thank Berenit Mendoza Garfias for her help in obtaining the scanning electron microphotographs. A. L.-J. and M. T. G.-G. acknowledge the support of the Programa de Posgrado en Ciencias Biológicas, UNAM and CONACYT (A. L.-J. CVU No. 706119; M. T. G.-G. CVU No. 956064), for granting a scholarship to complete her PhD and his Master program, respectively. L. A.-G. acknowledges the Coordinación de la Investigación Científica and Dirección General de Asuntos de Personal Académico (DGAPA-UNAM), Mexico, for granting the Postdoctoral Fellowship.
Author's contributions
M. G.-V. and A. L. S.-U. conceived and designed the study. M. G.-V., A. L.-J. and M. T. G.-G. conducted data gathering. A. L.-J. and M. T. G.-G. performed statistical analyses. A. L. S.-U., L. A.-G. and M. G.-V. wrote and edited the article.
Financial support
This research was supported by the Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (PAPIIT-UNAM) IN201122.
Conflict of interest
No conflict of interest exists among the authors.
Ethical standards
The sampling in this work complies with the current laws and animal ethics regulations of México. Specimens were collected under the Cartilla Nacional de Colector Científico (FAUT 0202) issued by the Secretaría del Medio Ambiente y Recursos Naturales (SEMARNAT), to M. G.-V.












 Open access
Open access