



Introduction
Population dynamics of parasite species are influenced by multiple internal and external conditions and population sizes of a given parasite species can therefore vary considerably in space and time. One obvious potential driver of parasite population dynamics is the availability and abundance of suitable hosts (Arneberg et al., Reference Arneberg, Skorping, Grenfell and Read1998; Kmentová et al., Reference Kmentová, Gelnar, Mendlová, van Steenberge, Koblmüller and Vanhove2016; Song and Proctor, Reference Song and Proctor2020). Habitat age and stability, or time since colonization, are further potential drivers (Song and Proctor, Reference Song and Proctor2020). In addition to natural influences, parasites also respond to anthropogenic stressors, which has led to investigations regarding the usefulness of parasite abundance as bioindicator of various types of pollution (Poulin, Reference Poulin1992; Lafferty, Reference Lafferty1997; Blanar et al., Reference Blanar, Munkittrick, Houlahan, MacLatchy and Marcogliese2009; Vidal-Martinez et al., Reference Vidal-Martinez, Pech, Sures, Purucker and Poulin2010; Sures et al., Reference Sures, Nachev, Selbach and Marcogliese2017; Marcogliese, Reference Marcogliese2023). The effects of pollution on parasite abundance can be positive, e.g. when environmental stress increases host susceptibility, or negative, when environmental stressors weaken the parasites themselves (Blanar et al., Reference Blanar, Munkittrick, Houlahan, MacLatchy and Marcogliese2009; Marcogliese, Reference Marcogliese2023). In a recent study, the prevalence of acanthocephalan parasites in their intermediate amphipod hosts was positively correlated with local pollution levels along an anthropogenically impacted river in southern France (Fanton et al., Reference Fanton, Franquet, Logez, Cavalli and Kaldonski2022). Conversely, the abundance of acanthocephalans in the definitive fish host was negatively correlated with pollution levels in a southern Brazilian river (Lacerda et al., Reference Lacerda, Roumbedakis, Junior, Nuñer, Petrucio and Martins2018).
Acanthocephalans are endoparasitic worms with complex life cycles involving at least one arthropod intermediate and one vertebrate definitive host (Kennedy, Reference Kennedy2006). Intermediate hosts of Acanthocephala in European freshwater systems are amphipods and aquatic isopods, while definitive hosts differ between acanthocephalan genera. Species of Pomphorhynchus and Echinorhynchus, for example, complete their life cycle in fish, whereas birds serve as final hosts for Polymorphus spp. The mature worms reproduce in the digestive tract of their vertebrate hosts and their eggs are shed into the water with the feces. The intermediate hosts are then infected by ingesting the parasite eggs. The parasite larvae hatch in the intestines and develop in the body cavities of the intermediate hosts until they reach the infective stage for the definitive host (Sures and Schmidt-Rhaesa, Reference Sures and Schmidt-Rhaesa2014). Several acanthocephalan species are known to manipulate the behaviour of their intermediate hosts to enhance trophic transmission to the definitive host (Bakker et al., Reference Bakker, Frommen and Thünken2017).
Common acanthocephalan species found in Central Europe are Polymorphus minutus, Pomphorhynchus laevis, Pomphorhynchus tereticollis and Pomphorhynchus bosniacus (Ladewig et al., Reference Ladewig, Jungmann, Köhler, Schirling, Triebskorn and Nagel2006; Westram et al., Reference Westram, Baumgartner, Keller and Jokela2011; Bauer and Rigaud, Reference Bauer and Rigaud2015; Galipaud et al., Reference Galipaud, Bollache and Lagrue2019; Reier et al., Reference Reier, Sattmann, Schwaha, Harl, Konecny and Haring2019, Reference Reier, Sattmann, Schwaha, Fuehrer and Haring2020; Jirsa et al., Reference Jirsa, Reier and Smales2021; Fanton et al., Reference Fanton, Franquet, Logez, Cavalli and Kaldonski2022). Genetic data revealed considerable intraspecific genetic diversity and, in some of these species, also geographic structure across Europe (Smrzlić et al., Reference Smrzlić, Valić, Kapetanović, Marijić, Gjurčević and Teskeredžić2015, Reference Smrzlić, Čolić, Kapetanović, Šariri, Mijošek and Filipović Marijić2024; Perrot-Minnot et al., Reference Perrot‐Minnot, Špakulová, Wattier, Kotlik, Düşen, Aydoğdu and Tougard2018; Zittel et al., Reference Zittel, Grabner, Wlecklik, Sures, Leese, Taraschewski and Weigand2018; Reier et al., Reference Reier, Sattmann, Schwaha, Harl, Konecny and Haring2019, Reference Reier, Sattmann, Schwaha, Fuehrer and Haring2020, Jirsa et al., Reference Jirsa, Reier and Smales2021). Polymorphus minutus has been shown to encompass three morphologically cryptic but genetically divergent species, separated by 3.5–11% distance in cytochrome oxidase I (COI) sequences, each of which was associated with a different amphipod host species (Zittel et al., Reference Zittel, Grabner, Wlecklik, Sures, Leese, Taraschewski and Weigand2018). The cryptic species are widely distributed in Germany and France (Zittel et al., Reference Zittel, Grabner, Wlecklik, Sures, Leese, Taraschewski and Weigand2018), and one of them (Polymorphus sp. type 3) has meanwhile also been recorded further east, in Austria (Jirsa et al., Reference Jirsa, Reier and Smales2021). Somewhat lower levels of genetic divergence were detected in P. laevis (mean 2%, max. 3.5%, COI) and P. tereticollis (mean 1.8%, max. 3.8%, COI), with P. tereticollis showing a weak phylogeographic structure comprising four haplogroups across Europe (Reier et al., Reference Reier, Sattmann, Schwaha, Harl, Konecny and Haring2019). P. laevis contains two clearly distinct haplogroups (Eastern and Western European), of which only the Eastern European haplogroup was detected in Austria (Reier et al., Reference Reier, Sattmann, Schwaha, Harl, Konecny and Haring2019). The extent of intraspecific genetic diversity and population and phylogeographic structure present in Acanthocephala is expected to reflect host specificity, mobility of the host and demographic history of the parasite populations (Song et al., Reference Song, Li, Wu, Zou and Wang2014; García-Varela et al., Reference García-Varela, López-Jiménez, González-García, Sereno-Uribe and Andrade-Gómez2023; Sromek et al., Reference Sromek, Ylinen, Kunnasranta, Maduna, Sinisalo, Michell, Kovacs, Lydersen, Ieshko and Andrievskaya2023), but population level data on genetic diversity in Acanthocephala are still rather scarce.
The potential ecological consequences of Acanthocephala infections of amphipods are diverse. Amphipod population and invasion dynamics can be altered by parasite-induced effects on predation risk, overall mortality, competitive ability, sensitivity to pollutants and other environmental stressors (Giari et al., Reference Giari, Fano, Castaldelli, Grabner and Sures2020; Kochmann et al., Reference Kochmann, Laier, Klimpel, Wick, Kunkel, Oehlmann and Jourdan2023). On an even broader scale, parasite-induced modifications of how amphipods interact with their biotic and abiotic environments can have implications for entire aquatic ecosystems (Giari et al., Reference Giari, Fano, Castaldelli, Grabner and Sures2020), for instance due to the reduced shredding and increased bioturbation activity of infected amphipods (Labaude et al., Reference Labaude, Rigaud and Cézilly2017; Williams et al., Reference Williams, Donohue, Picard, O’Keeffe and Holland2019).
The abundance and widespread occurrence of amphipods in streams and rivers also makes their infection status an attractive potential proxy for the ecological status of their aquatic environment, especially considering that Acanthocephala larvae can be detected with the naked eye through the amphipod cuticle (Figure 1). However, while the idea to use acanthocephalans in environmental monitoring has been brought up repeatedly (Lafferty, Reference Lafferty1997; Fanton et al., Reference Fanton, Franquet, Logez, Cavalli and Kaldonski2022; Perrot-Minnot et al., Reference Perrot-Minnot, Cozzarolo, Amin, Barčák, Bauer, Marijić, García-Varela, Hernández-Orts, Le and Nachev2023; Sures et al., Reference Sures, Nachev, Schwelm, Grabner and Selbach2023), the relationships between environmental stressors and Acanthocephala abundance are not investigated exhaustively (Blanar et al., Reference Blanar, Munkittrick, Houlahan, MacLatchy and Marcogliese2009; Lacerda et al., Reference Lacerda, Roumbedakis, Junior, Nuñer, Petrucio and Martins2018; Fanton et al., Reference Fanton, Franquet, Logez, Cavalli and Kaldonski2022). Clearly, the prevalence of acanthocephalan parasites in European streams and rivers varies depending on study site, parasite and host species (Ladewig et al., Reference Ladewig, Jungmann, Köhler, Schirling, Triebskorn and Nagel2006; Westram et al., Reference Westram, Baumgartner, Keller and Jokela2011; Bauer and Rigaud, Reference Bauer and Rigaud2015; Galipaud et al., Reference Galipaud, Bollache and Lagrue2019; Fanton et al., Reference Fanton, Franquet, Logez, Cavalli and Kaldonski2022), and more data are needed to evaluate the potential factors at various geographic scales. In the present study, we focus on several small streams that originate in the hills around the city of Graz (Styria, Austria) and cross forested, agricultural and urban landscapes before flowing into the River Mur. During previous amphipod collections, we had already noticed acanthocephalan infections of amphipods in this region, particularly at sites in the more developed, urban area. Urban stream sections are typically surrounded by artificial, impervious surfaces; are hydromorphologically impacted by stream bed modifications; and are exposed to elevated pollution due to, e.g. road run-off (Allan, Reference Allan2004). Relationships between fish parasite abundances and degrees of urbanization have been demonstrated in both freshwater and marine systems (Hernandez et al., Reference Hernandez, Bunnell and Sukhdeo2007; Taglioretti et al., Reference Taglioretti, Rossin and Timi2018; Shah Esmaeili et al., Reference Shah Esmaeili, Checon, Corte and Turra2021). To examine whether spatial variation in acanthocephalan prevalence was associated with the urban environment, prevalence estimates in urban and residential stream sections were compared with stream sections traversing the surrounding rural landscape, which is characterized by agricultural land and forests.

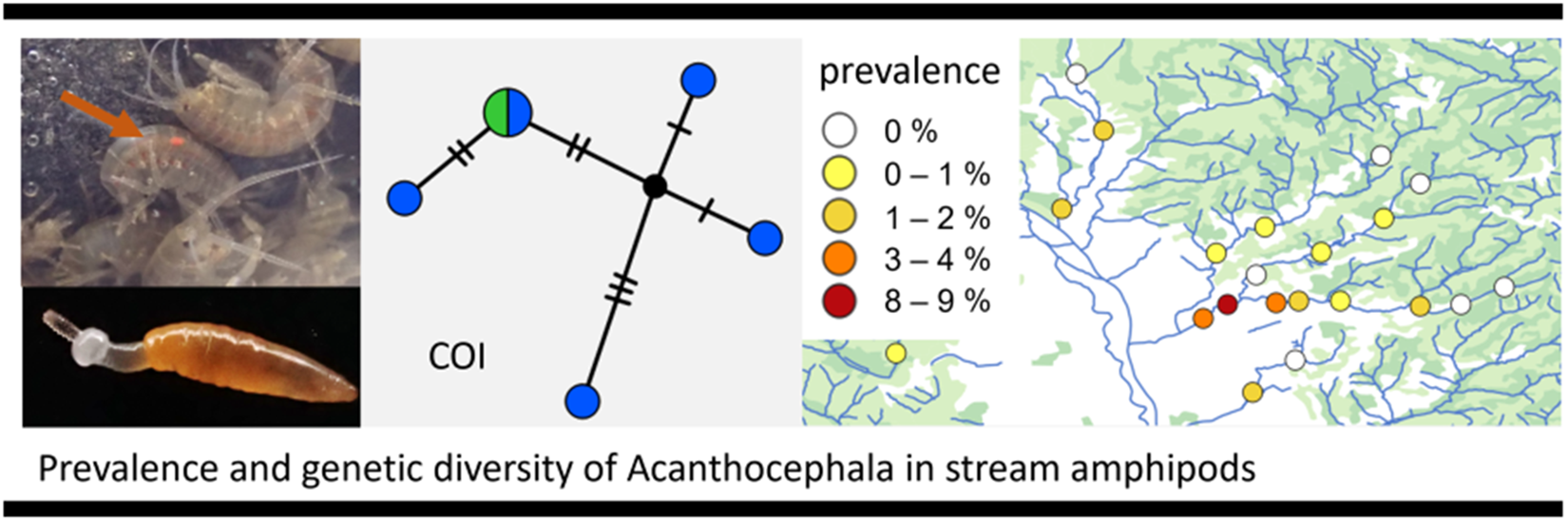
Figure 1. A group of Gammarus fossarum, one of which is visibly infected with an orange-coloured acanthocephalan (arrow). Photo: K. M. Sefc.
Furthermore, we assessed species diversity and intraspecific genetic diversity of Acanthocephala in the study region. To this aim, the acanthocephalan larvae were extracted from their amphipod hosts and identified to species level using DNA barcoding (Hebert et al., Reference Hebert, Cywinska, Ball and DeWaard2003). We then used the DNA sequence data to estimate genetic diversity in the parasite populations, and joined the new sequences with published datasets to relate the local diversity in our study region to the phylogeographic patterns across the distribution ranges.
Material and methods
Parasite prevalence
Gammarus fossarum were collected from 25 sampling sites in small streams in and around the city of Graz, Austria, located in the eastern foothills of the European Alps (Figure 2, Tables 1 and S1). The sampling sites comprised five stream drainage systems which discharge into the river Mur (Figure 2, Table 1). Land use around the sampling sites varied and included built-up areas in the city, as well as agricultural and forested areas in the periphery. Stream widths at the sampling sites ranged from 0.5 to 3 m and water depths ranged from 10 to 30 cm. To assess temporal variation in parasite prevalence, one site (#12) was sampled once a month from April to August 2023, and once more in October 2023 (Table S2). The other sites were sampled between mid-April and early July 2023. Amphipods were collected by disturbing the stream bed and detritus patches, upon which the animals were flushed into hand nets by the water current. Sampling effort was not standardized across sites, but we aimed at sample sizes of 100 or more amphipods to allow the estimation of low infection prevalence. All amphipods collected in the net were taken to the laboratory to estimate parasite prevalence, and zoobenthos other than amphipods were returned to the stream. Temperature, conductivity and oxygen concentration and saturation were measured at sampling sites using a handheld multiparameter probe (pHenomenal MU 6100H by VWR).
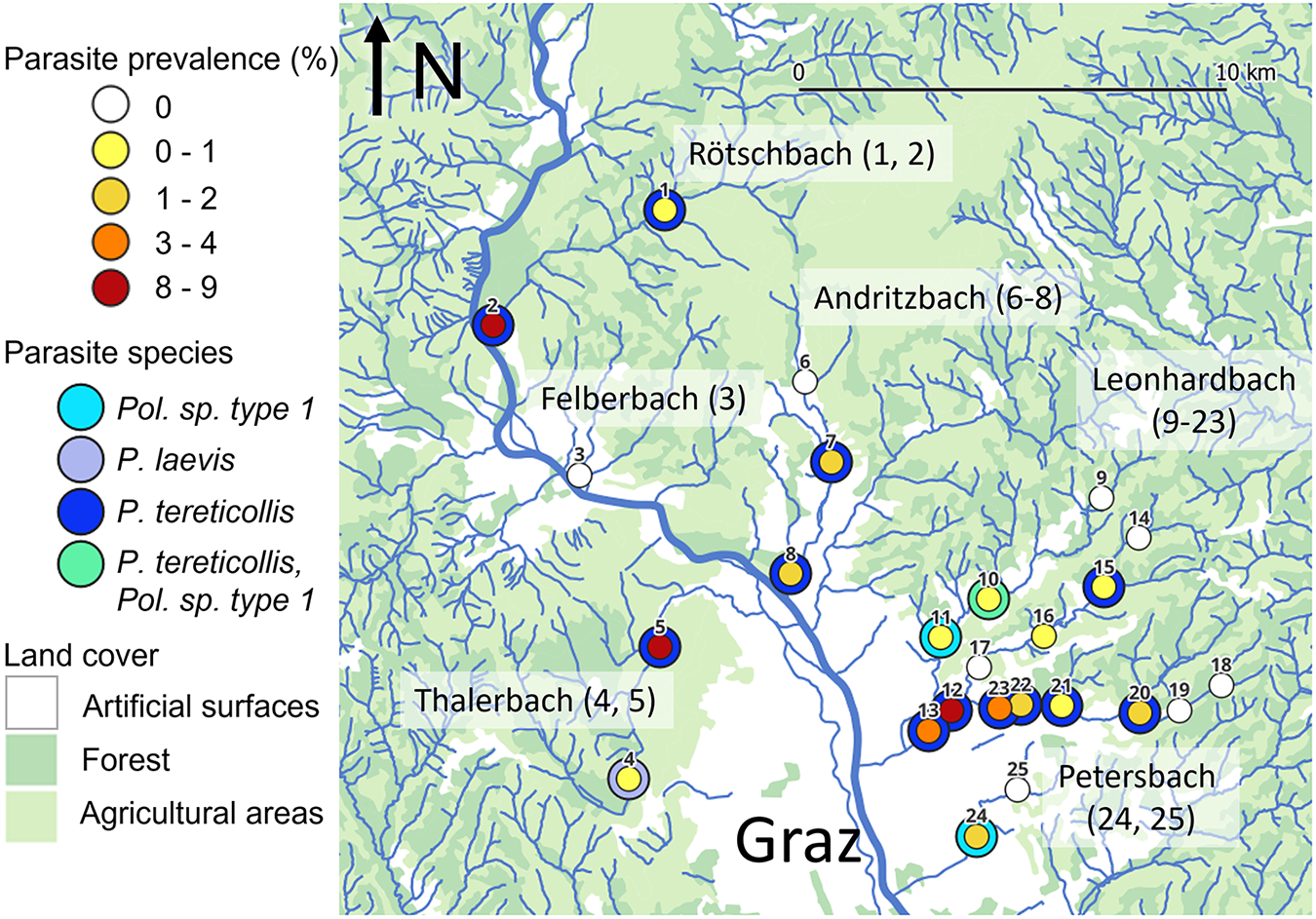
Figure 2. Acanthocephala prevalence and species at the study sites. Study sites are numbered from 1 to 25 as in Table 1. The colour of the inner circle represents the prevalence category, and the colour of the outer ring represents the parasite species found at the site (if any). The River Mur is drawn in a thick line, and the six tributary stream systems are named according to the stream name at the confluence with the Mur, with the corresponding site numbers in parentheses. The map was created in QGIS 3.42.2, using the European Union’s Copernicus Land Monitoring Service information (https://doi.org/10.2909/71c95a07-e296-44fc-b22b-415f42acfdf0) and GIS stream data provided by the Umweltbundesamt GmbH (https://docs.umweltbundesamt.at/s/8rYyRjFDse8Za4D).
Table 1. Sampling sites and acanthocephalan parasitism
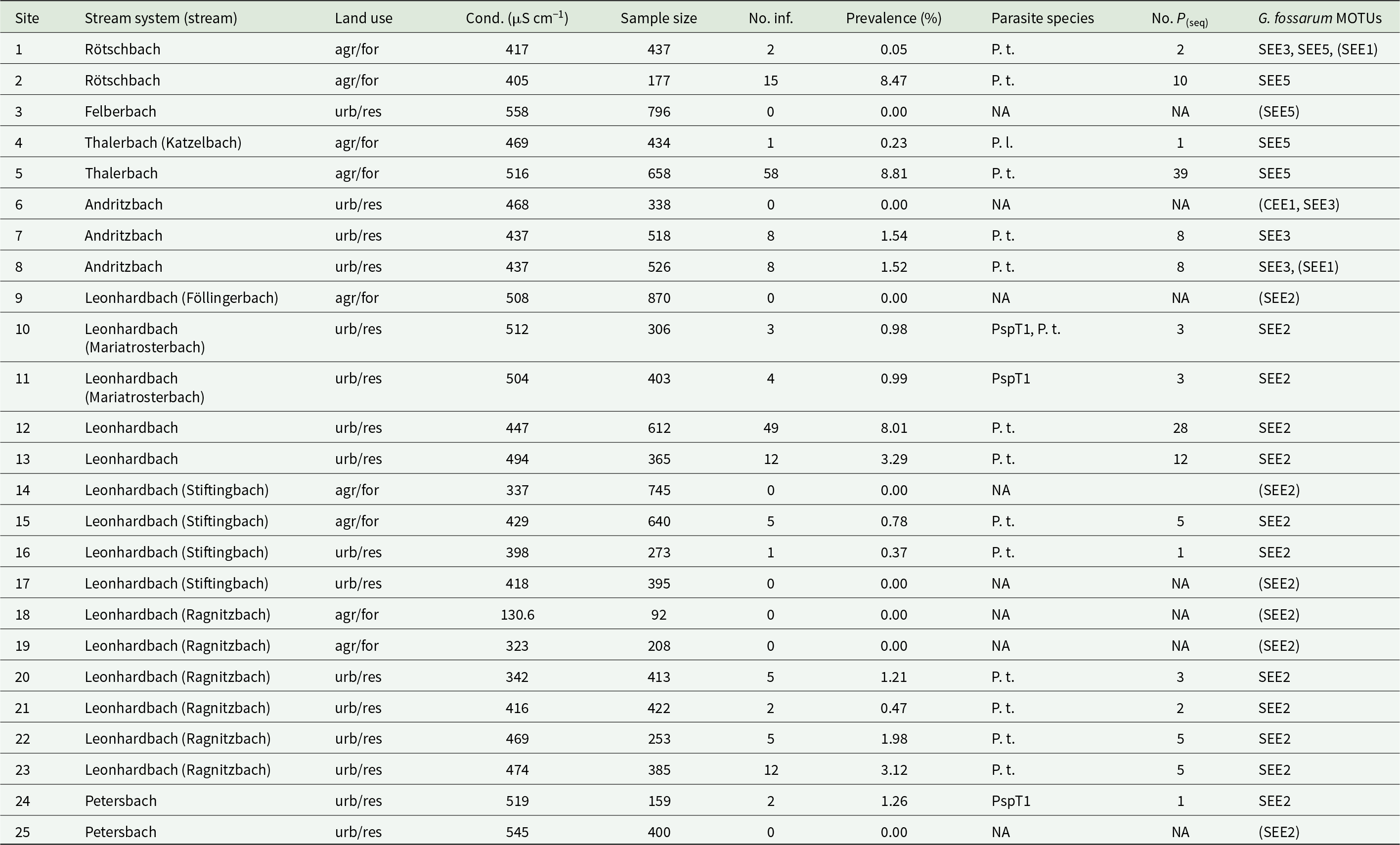
Site numbering and stream systems correspond to Figure 2; stream names that differ from the name of the stream system are given in parentheses. ‘Sample size’ refers to the number of examined amphipods, and ‘No. inf.’ is the number of infected amphipods. Land use is scored as agricultural and forest (‘agr/for’) and urban and residential (‘urb/res’). Abbreviations for parasite species are as follows: P. t., Pomphorhynchus tereticollis; P. l., Pomphorhynchus laevis; PspT1, Polymorphus sp. type 1 (Zittel et al., Reference Zittel, Grabner, Wlecklik, Sures, Leese, Taraschewski and Weigand2018). ‘No. P(seq)’ indicates the number of Acanthocephala individuals for which COI barcode sequences were obtained. Conductivity (cond.) was measured at the time of amphipod sampling. G. fossarum MOTUs identified at the study sites are indicated by clade (SEE or CEE, sensu Wattier et al., Reference Wattier, Mamos, Copilaş-Ciocianu, Jelić, Ollivier, Chaumot, Danger, Felten, Piscart and Žganec2020) and an internal lab number; MOTUs that were not infected by Acanthocephala are written in parentheses. Further information on sampling sites is given in Table S1.
Gammarus fossarum was the only amphipod species present at the study sites. Sample sizes ranged from 92 to 870 amphipods per site (mean = 432.8, median = 403; Table 1). In the laboratory, visibly infected amphipods (based on orange acanthella or cystacanth stages that could be detected through the host cuticle) were separated from non-infected amphipods and euthanized in carbonated water. Infected amphipods were counted and individually weighed on a laboratory scale. Then, the acanthocephalan larvae were extracted from their hosts and preserved in 99% ethanol for DNA barcoding. With few exceptions, each host contained a single parasite larva (at the high-prevalence site #5, 2 host individuals each contained 4 larvae). The host tissues were stored separately.
The uninfected amphipods were also euthanized in carbonated water. Three of the smallest and three of the largest individuals were weighed individually to estimate the weight range, and the remaining individuals were weighed in bulk. The mean weight of uninfected amphipods was determined based on the sum of the individual and bulk weights. The number of uninfected amphipods was determined by automatic counting. To do this, the amphipods were spread out on a semi-translucent surface placed on a visible light transilluminator, making sure that individuals did not touch each other, and photographed. The ‘Analyze Particles’ function of ImageJ was used to count the amphipods on the photos.
Acanthocephala prevalence was calculated as the proportion of infected amphipods in the sample collected at each site. To examine potential drivers of spatial variation in Acanthocephala prevalence, we tested for correlations with land cover, conductivity and distance from source. Land cover in a 100 m radius around each site was determined based on the CORINE land cover 2018 dataset (using the European Union’s Copernicus Land Monitoring Service information, https://doi.org/10.2909/71c95a07-e296-44fc-b22b-415f42acfdf0) and classified as urban and residential areas (CORINE class ‘artificial surfaces’) or natural surface (combining the CORINE classes ‘agricultural areas’ and ‘forest and seminatural areas,’ which alternate at small spatial scales in the study area). Conductivity measures the presence of ions (e.g. chloride, sulphite and nitrate) in the water and is related to water quality (Harwell et al., Reference Harwell, Surratt, Barone and Aumen2008; Thompson et al., Reference Thompson, Brandes and Kney2012; Skarbøvik and Roseth, Reference Skarbøvik and Roseth2015) and the ecological status of water bodies (Potapova and Charles, Reference Potapova and Charles2003; Zampella et al., Reference Zampella, Bunnell, Laidig and Procopio2006). Land cover and conductivity represented variation in environmental conditions, which might be associated with variation in environmental quality. Distance from source reflects the locations of the sampling sites in upper, middle and lower parts of the streams and might be connected to parasite prevalence in amphipods, if Acanthocephala eggs shed by the definite hosts accumulated downstream. Distances between sampling sites and the source of the respective streams along the stream course were calculated using the GIS stream layer provided by the Umweltbundesamt GmbH (https://docs.umweltbundesamt.at/s/8rYyRjFDse8Za4D). We designated the farthest upstream spring outlet as source point for this calculation. We fitted a general linear model (GLM) with a negative binomial error distribution (R package MASS, function glm.nb; Venables and Ripley, Reference Venables and Ripley2022), including the number of infected amphipods as response variable. To account for differences in the total number of amphipods that were collected per site, sample size was included as offset variable. Model predictors were surrounding land cover (nominal variable with 2 levels: ‘artificial surface’, i.e. urban and residential areas; and ‘natural surface’, i.e. agricultural, forest and seminatural areas), conductivity (metric variable, in μS cm−1) and distance from source (metric variable, in kilometres). An alternative model (overdispersed binomial logit model; R package dispmod, function glm.binomial.disp; Scrucca, Reference Scrucca2018) yielded qualitatively identical results. Collinearity among predictors was measured by the variance inflation factor (VIF) calculated in the R package ‘car’ (Fox and Weisberg, Reference Fox and Weisberg2019). VIFs of 1 indicate no collinearity, while VIFs >5 indicate high collinearity between predictors and coefficients cannot be estimated reliably. For site #12 (which had been sampled repeatedly), we summed the numbers of infected and uninfected amphipods collected in April, May and June, and took the mean of the conductivity and oxygen saturation values measured at these time points. Omitting site #12, covariation between parasite prevalence and sampling time was tested in a separate GLM, with ‘day’ (starting with 0 for the first day of sampling) as predictor.
DNA barcoding and genetic diversity
DNA was extracted from whole Acanthocephala larvae using a standard Chelex protocol (Richlen and Barber, Reference Richlen and Barber2005). The DNA barcode region of the COI gene was amplified using primers LCO1490-JJ2 (5ʹ-CHACWAAYCAYAARGAYATYGG) and HCO2198-JJ2 (ANACTTCNGGRTGNCCAAARAATCA) (Astrin et al., Reference Astrin, Höfer, Spelda, Holstein, Bayer, Hendrich, Huber, Kielhorn, Krammer and Lemke2016). The polymerase chain reactions (PCRs) contained 0.35 µL of a 2.5 mM dNTP mix, 0.5 µL of a 50 mM MgCl2 solution, 0.3 µL BioThermRed™ Taq DNA Polymerase (5 U µL−1) and 1.0 µL 10× buffer (including 15 mM MgCl2), 1.0 µL of each primer (10 mM stocks) and 1.5 µL template DNA in a total volume of 10 µL. The temperature regime consisted of initial denaturation for 3 min at 94 °C followed by 45 cycles of denaturation (94 °C for 30 sec), annealing (49 °C for 35 sec) and extension (72 °C for 1 min), with a final extension at 72 °C for 7 min.
Sanger sequencing of amplicons failed for many parasite samples because the PCR products were contaminated with amplicons derived from host DNA and from parasite pseudogenes. We therefore used Oxford Nanopore Technology (ONT) to obtain sequence reads. For this purpose, both the forward and the reverse PCR primers carried 13-bp tags (Srivathsan et al., Reference Srivathsan, Hartop, Puniamoorthy, Lee, Kutty, Kurina and Meier2019) on the 5ʹ end, resulting in sample-specific tag combinations that allowed demultiplexing of sequence reads. For sequencing library preparation, 2 µL of PCR products were pooled and the pool was purified using AMPure XP magnetic beads (Beckman Coulter, Brea, CA, USA). The concentration of the amplicon pool was measured with a Qubit 4 Fluorometer using the Qubit dsDNA HS Assay Kit (Invitrogen by Thermo Fisher Scientific, Waltham, MA, USA). Further library preparation was performed using the Ligation Sequencing Kit (SQK-LSK112 for the first batch of samples, SQK-LSK114 for the second batch of samples, both ONT), the NEBNext Ultra II End repair/dA-tailing Module (E7546, New England Biolabs) for DNA strand end repair and the NEBNext Quick Ligation Module (E6056, New England Biolabs) for adapter ligation, and followed the corresponding protocols. Sequencing of the first batch of samples (yielding 86 COI sequences) was performed on a Flongle flow cell (R9.4.1; FLO-FLG001, ONT) primed with the Flongle Flow Cell Priming Kit (EXP-FSE001, ONT); the remaining samples (yielding 50 COI sequences) were sequenced on the Flongle flow cell R10.4.1; FLO-FLG114 (ONT) primed with the Flongle Flow Cell Priming Kit (EXP-FSE002, ONT). Raw ONT data were basecalled using guppy_basecaller of Guppy version 6.4.2 with a quality threshold of five (–min_qscore 5) and the basecalling model dna_r9.4.1_450bps_sup.cfg. The ONTbarcoder software (Srivathsan et al., Reference Srivathsan, Lee, Katoh, Hartop, Kutty, Wong, Yeo and Meier2021) was used to demultiplex the sequencing reads, and AmpliconSorter (Vierstraete and Braeckman, Reference Vierstraete and Braeckman2022) was used to sort the demultiplexed reads by species and to obtain consensus barcode sequences for parasites and their hosts. Using the same materials as in the first batch of the present parasite sequencing (SQK-LSK112 library kit; R9.4.1, FLO-FLG001 cell; EXP-FSE001 priming kit), nanopore-based COI barcode sequences were shown to be consistent with and of the same quality as traditional Sanger sequencing (Koblmüller et al., Reference Koblmüller, Resl, Klar, Bauer, Zangl and Hahn2024). The parasite consensus sequences were based on 9-1496 reads per sample. Only 5% of all consensuses (n = 136 parasite sequences) were built from less than 50 reads, and these weakly supported consensuses were identical to the most common haplotype of P. tereticollis in the dataset. The DNA sequence length was 655 bp. The DNA sequences were checked for compliance with the COI reading frame and blasted against GenBank for parasite species identification. The acanthocephalan COI sequences are available at GenBank (accession numbers PV792653-PV792788). Host sequences were compared to our in-lab database of local genetic lineages of G. fossarum (molecular operational taxonomic units, MOTUs; diBatista Borko, Sefc, et al., unpublished data).
In addition to the sequences generated in this study, publicly available COI sequences of P. tereticollis and P. laevis were retrieved from GenBank (accession numbers provided in Supplementary Table S3). These sequences were combined with the Nanopore-derived consensus sequences and DNA sequence alignments were performed in MEGAX vs 10.2.5 (Kumar et al., Reference Kumar, Stecher, Li, Knyaz and Tamura2018). Statistical parsimony haplotype networks were plotted in PopART (Templeton et al., Reference Templeton, Crandall and Sing1992; Leigh and Bryant, Reference Leigh and Bryant2015) using the TCS method (Templeton et al., Reference Templeton, Crandall and Sing1992) to visualize genetic relationships among the parasite haplotypes.
Results
Temporal variation in parasite prevalence
Acanthocephala prevalence was measured repeatedly, once a month from April to August and again in October, at study site #12 to assess variation over time. Prevalence estimates ranged from 7% to 9% in April (8.5%), May (7.2%) and June (9.0%), then dropped in the samples taken at the end of July (1.0%) and August (1.9%), and returned to 7% in October (6.9%). Sample sizes and number of infected amphipods are given in Table S2. At the other collection sites, prevalence was estimated only once between mid-April and early July and no decline in parasitism was observed over this period (GLM estimate = 0.01, z = 0.62, P = 0.53).
Spatial variation in parasite prevalence
Acanthocephala prevalence at sampling sites ranged from 0% (8 sites) to 8.8% (Table 1; Figure 2). The distribution of prevalence estimates was bimodal, with the majority of sites below 4% and three sites above 8%. The sites with the highest prevalence estimates (>8%) were located in the downstream reaches of three different streams and surrounded by both natural and artificial land surfaces. In contrast, prevalence estimates ranged from 0% to 2% in the upstream reaches, all of which were located in forested and agricultural areas. Neither surrounding land cover nor conductivity measured at the time of sampling were statistically linked to parasite prevalence, but prevalence increased significantly with distance from source (Figure 3; land cover, artificial vs natural, GLM estimate = 0.59, z = 1.14, P = 0.25; conductivity, GLM estimate = 0.0003, z = 0.071, P = 0.94; distance from source, GLM estimate = 0.36, z = 5.23, P = 1.7 × 10−7; VIF(land cover) = 1.10, VIF(conductivity) = 1.06, VIF(distance) = 1.04). A model excluding the 3 sites with the highest prevalence (>8%) yielded similar results.
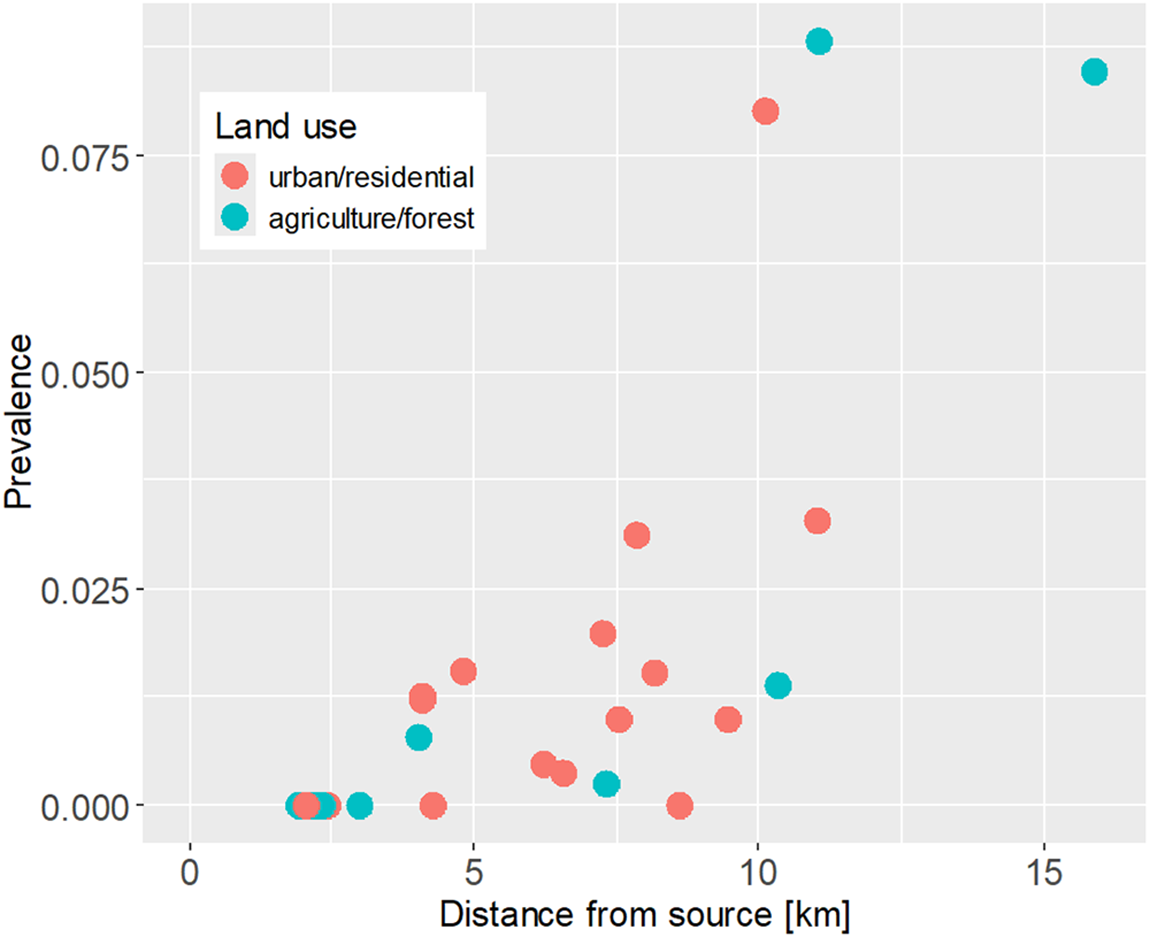
Figure 3. Increase of parasite prevalence with distance from stream source. Each dot represents a study site, with colours corresponding to surrounding land use.
Identification of parasite and host species
COI barcode sequences were successfully obtained from 136 of 154 attempted Acanthocephala samples (in the remaining cases, PCR failed or only host sequences were obtained). The majority of the sequences (n = 129) were identified as P. tereticollis by comparison to archived DNA barcodes (Figure 2). One sample was identified as P. laevis (Figure 2). The COI haplotype of this sample was identical to ones found previously in the River Mur in Austria (Reier et al., Reference Reier, Sattmann, Schwaha, Harl, Konecny and Haring2019), and clustered with the ‘eastern lineage’ of P. laevis (Perrot‐Minnot et al., Reference Perrot‐Minnot, Špakulová, Wattier, Kotlik, Düşen, Aydoğdu and Tougard2018) in a network of European samples (Supplementary Fig. S1). Finally, six samples (from three sites in two streams, Figure 2) corresponded to Polymorphus sp. type 1 (Zittel et al., Reference Zittel, Grabner, Wlecklik, Sures, Leese, Taraschewski and Weigand2018), a member of the Polymorphus minutus species complex. With the exception of site #10, where both Polymorphus sp. type 1 and P. tereticollis were detected, only one Acanthocephala species was detected per site (Figure 2).
By sequencing the PCR products produced from the Acanthocephala DNA extracts, we also obtained the DNA barcode sequences of 129 G. fossarum hosts. The hosts represented five distinct mitochondrial genetic lineages (MOTUs; Table 1) across all study sites, which fell within the SEE and CEE clades defined by Wattier et al., Reference Wattier, Mamos, Copilaş-Ciocianu, Jelić, Ollivier, Chaumot, Danger, Felten, Piscart and Žganec2020. The parasitized amphipods belonged to the three most frequent MOTUs in the study streams. No relationships between host MOTU, parasite species and parasite prevalence were apparent in the present dataset (Table 1).
Genetic diversity of parasite COI haplotypes
We detected 10 distinct haplotypes in P. tereticollis (n = 129 individuals; Figure 4), and the estimated haplotype diversity (He) was 0.32. Two haplotype clades were separated from each other by nine mutations (1.4% divergence). One of the clades corresponds to haplogroup 3 defined by Reier et al. (Reference Reier, Sattmann, Schwaha, Harl, Konecny and Haring2019), which comprises P. tereticollis collected across Europe (Supplementary Fig. S3). The other clade corresponds to haplogroup 4 (Reier et al., Reference Reier, Sattmann, Schwaha, Harl, Konecny and Haring2019) and has so far only been found in southeastern Austria (in the present study, and approximately 50 km further north in the River Mur, Reier et al., Reference Reier, Sattmann, Schwaha, Harl, Konecny and Haring2019, Supplementary Fig. S3). The most common haplotype in the present study, which was shared by the majority of individuals (67%), belonged to this clade and occurred at all sites where P. tereticollis were collected (Figure 4). Additional haplotypes were detected in all stream systems, with the highest diversity in the Rötschbach (h = 5 haplotypes; sites #1 and #2, n = 12 sequenced individuals), and the lowest diversity in the Leonhardbach system (h = 2 haplotypes; sites #9–#23, n = 67 sequenced individuals, Figure 4). Four of the sampling sites contained more than 1 P. tereticollis haplotype (#2, h = 4; #5, h = 3; #8, h = 4; #21, h = 2).
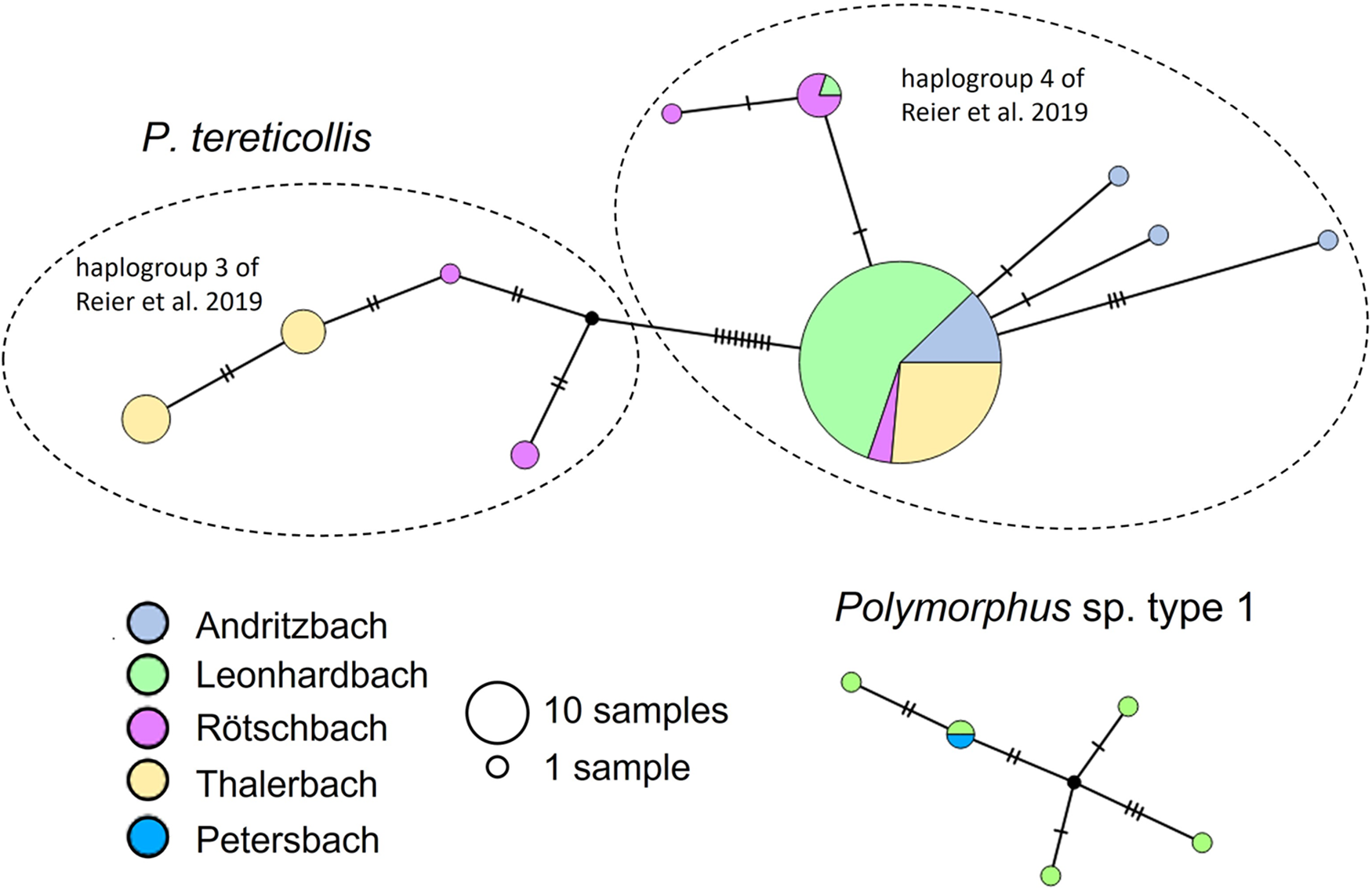
Figure 4. COI haplotype networks of P. tereticollis and Polymorphus sp. type 1. Colour coding and size of circles refer to stream systems and number of samples, respectively. The tick marks on the lines connecting the haplotypes indicate the number of nucleotide differences between haplotypes. In the network of P. tereticollis, dashed lines mark the haplogroups defined by Reier et al. (Reference Reier, Sattmann, Schwaha, Harl, Konecny and Haring2019).
Polymorphus sp. type 1 (n = 6 individuals from three sites in two stream systems) contained five different haplotypes (He = 0.78), one of which was shared between the two stream systems in which the species was found (Figure 4).
Discussion
Parasite species diversity in G. fossarum
The prevalence of Acanthocephala at the study sites was in the single digit range. The majority of parasites extracted from the amphipod hosts were P. tereticollis, whose prevalence per site reached up to 8% of examined amphipods. This species has previously been shown to have a high prevalence in cyprinid and salmonid fish in the River Mur, to which the studied streams are tributaries (Reier et al., Reference Reier, Sattmann, Schwaha, Harl, Konecny and Haring2019). Polymorphus sp. type 1 (a member of the Polymorphus minutus species complex, Zittel et al., Reference Zittel, Grabner, Wlecklik, Sures, Leese, Taraschewski and Weigand2018) was found in two adjacent sites in one stream at a very low prevalence (<1%), and a single infection with P. laevis was detected in another stream. P. laevis had previously been detected in fish near the area where we conducted our study (Reier et al., Reference Reier, Sattmann, Schwaha, Harl, Konecny and Haring2019). The specimen collected in the course of this study shared one of the two haplotypes previously identified in the Mur (Reier et al., Reference Reier, Sattmann, Schwaha, Harl, Konecny and Haring2019), which fits into broader patterns of genetically structured lineages across Europe (Supplementary Fig. S1) as described by Perrot‐Minnot et al. (Reference Perrot‐Minnot, Špakulová, Wattier, Kotlik, Düşen, Aydoğdu and Tougard2018), who highlighted a strong east–west differentiation and several distinct mitochondrial clades. The lineage found in this study corresponds to the Eastern lineage, which was also detected in the Sava basin (Smrzlić et al., Reference Smrzlić, Valić, Kapetanović, Marijić, Gjurčević and Teskeredžić2015, Reference Smrzlić, Čolić, Kapetanović, Šariri, Mijošek and Filipović Marijić2024). Genetic similarity between samples from Mur and Sava drainages (both connected to the Danube) was also observed in P. tereticollis (haplogroup 3; Supplementary Fig. S3).
In contrast, the present identification of Polymorphus sp. type 1 is the first evidence for the occurrence of this cryptic species in Austria, which was previously found in western Germany (Zittel et al., Reference Zittel, Grabner, Wlecklik, Sures, Leese, Taraschewski and Weigand2018). Polymorphus sp. type 1 has so far been shown to infect one particular genetic lineage of G. fossarum (type B, Zittel et al., Reference Zittel, Grabner, Wlecklik, Sures, Leese, Taraschewski and Weigand2018; Grabner et al., Reference Grabner, Doliwa, Bulantová, Horák and Sures2020). The G. fossarum lineage infected by Polymorphus sp. type 1 in the present study belongs to a different G. fossarum clade separated from type B by over 20 million years of divergence (Wattier et al., Reference Wattier, Mamos, Copilaş-Ciocianu, Jelić, Ollivier, Chaumot, Danger, Felten, Piscart and Žganec2020), which suggests that Polymorphus sp. type 1 may be able to parasitize a broad range of G. fossarum types. Previous reports of Polymorphus cf. minutus in Austria refer to Polymorphus sp. type 3 (Jirsa et al., Reference Jirsa, Reier and Smales2021) collected from mallards in north-eastern Austria. This cryptic species uses G. roeselii as intermediate host (Zittel et al., Reference Zittel, Grabner, Wlecklik, Sures, Leese, Taraschewski and Weigand2018) and could therefore not have been detected in the present study, in which only G. fossarum were collected and dissected. However, both Gammarus species occur in the River Mur and it remains to be tested whether Polymorphus sp. type 3 also occurs in south-eastern Austria.
Parasite prevalence
The prevalence estimates in the present study are in the range of those reported from elsewhere in Europe, such as the prevalence of P. tereticollis and Polymorphus minutus in G. fossarum in Swiss streams (Westram et al., Reference Westram, Baumgartner, Keller and Jokela2011), the prevalence of P. tereticollis in unidentified gammarids in French rivers (Fayard et al., Reference Fayard, Cezilly and Perrot-Minnot2019), and the prevalence of Polymorphus minutus in G. fossarum in two small streams in Germany (Ladewig et al., Reference Ladewig, Jungmann, Köhler, Schirling, Triebskorn and Nagel2006). Other studies, however, have found substantially higher acanthocephalan prevalence at individual study sites (Bauer and Rigaud, Reference Bauer and Rigaud2015; Galipaud et al., Reference Galipaud, Bollache and Lagrue2019; Fanton et al., Reference Fanton, Franquet, Logez, Cavalli and Kaldonski2022). In Galipaud et al., Reference Galipaud, Bollache and Lagrue2019, G. fossarum were infected with P. tereticollis, P. laevis and Polymorphus minutus, and similar to our study, the highest prevalence was achieved by P. tereticollis.
The rarity of P. laevis in the studied streams is somewhat unexpected, given its presence in fish collected at Styrian sites in the Mur and its tributary, the Sulm (Reier et al., Reference Reier, Sattmann, Schwaha, Harl, Konecny and Haring2019). In that study, however, P. laevis was also detected at very low prevalence compared to P. tereticollis, suggesting that it plays only a minor role in the local parasite communities of the Mur and its tributaries. Similar findings have been reported elsewhere: In headwater systems of southern Germany, P. tereticollis was the dominant species found in salmonids, whereas the Western lineage of P. laevis (e.g. Perrot‐Minnot et al., Reference Perrot‐Minnot, Špakulová, Wattier, Kotlik, Düşen, Aydoğdu and Tougard2018) occurred only in small numbers (Ros et al., Reference Ros, Basen, Teschner and Brinker2020). The predominance of P. tereticollis in streams suggests that this species is more ecologically suited to colonizing rhithral environments than P. laevis. Support for this view also comes from intermediate host range studies: in the Weser river system (Germany), P. laevis was found exclusively in the native amphipod Gammarus pulex (which does not occur in Eastern Austria), while P. tereticollis showed greater flexibility in host use (Vogel and Taraschewski, Reference Vogel and Taraschewski2023). Its broader host range may enable P. tereticollis to maintain populations and become the dominant species across a wider array of environmental and ecological conditions.
Covariation between parasite abundance and urbanization has been observed in some fish parasite systems (Hernandez et al., Reference Hernandez, Bunnell and Sukhdeo2007; Taglioretti et al., Reference Taglioretti, Rossin and Timi2018; Shah Esmaeili et al., Reference Shah Esmaeili, Checon, Corte and Turra2021). In the present study, parasite prevalence did not differ significantly between sites in urban and residential areas on the one hand and sites surrounded by agricultural land and forests on the other hand. Instead, prevalence estimates increased towards downstream regions independent of surrounding land use. An exception was site #3, which was in the lower reaches of a stream close to the confluence with the river Mur, but had no infected amphipods. The stream in question is only a little more than 4 km long and likely too small to host a sufficient number of definitive hosts to establish a local parasite population. In general, the small streams in the study area are inhabited by trout, and fish also migrate from the Mur River into the lower reaches of the streams. Fish in the Mur in the study area include various cyprinids, salmonids, perch and grayling (Woschitz and Parthl, Reference Woschitz and Parthl2013). Waterfowl in the area are mainly mallards, which are common hosts of Polymorphus minutus (Jirsa et al., Reference Jirsa, Reier and Smales2021) and, in smaller numbers, mergansers. Both species can occasionally be found in the small streams, including their middle and upper reaches (FG and KMS, personal observation). Therefore, possible explanations for the downstream increase in prevalence include a downstream gradient of parasite eggs due to transport in the stream current, as well as an increasing density of definitive hosts, especially fish (Blasco-Costa et al., Reference Blasco-Costa, Koehler, Martin and Poulin2013). Upstream-downstream gradients in parasite infection levels, as observed here and elsewhere (e.g. Blasco-Costa et al., Reference Blasco-Costa, Koehler, Martin and Poulin2013), are plausible but not ubiquitous. Chub (Squalius cephalus) sampled along a Czech river experienced lower parasite (including acanthocephalan) infections in downstream compared to upstream sites (Wenger et al., Reference Wenger, Ondračková, Machala, Neča, Hyršl, Šimková, Jurajda, von der and Segner2010), and variation in the prevalence of Acanthocephala in gammarids along a French river did not follow a longitudinal gradient but covaried with levels of environmental pollution (Fanton et al., Reference Fanton, Franquet, Logez, Cavalli and Kaldonski2022).
In addition, artificial ponds and reservoirs stocked with fish that are connected to the study streams may have contributed to elevated parasite prevalence at downstream sites in our study. For example, site #5 was located downstream of a dammed reservoir stocked with various, mostly cypriniform, fish and had the highest prevalence of P. tereticollis in this study. In fish ponds upstream of our sampling site #7, stocked rainbow trout, that were collected three years prior to the current study, were found to be infected with P. tereticollis and P. bosniacus (Reier et al., Reference Reier, Sattmann, Schwaha, Fuehrer and Haring2020). The two P. tereticollis haplotypes in the fish ponds (Reier et al., Reference Reier, Sattmann, Schwaha, Fuehrer and Haring2020) are identical to ones detected in our study, which is consistent with connectivity between pond and stream populations of P. tereticollis. In contrast, we did not detect P. bosniacus in the amphipods despite its presence in the fish pond (Reier et al., Reference Reier, Sattmann, Schwaha, Fuehrer and Haring2020). This is unlikely to be a technical artefact of DNA-based species identification, as the COI primers used in our PCR correspond to the primers used for sequencing the COI of P. bosniacus in Reier et al. (Reference Reier, Sattmann, Schwaha, Fuehrer and Haring2020). Rather, it raises the question of whether this parasite can complete its life cycle using the local G. fossarum lineages as intermediate hosts. P. bosniacus is thought to have colonized the Danube and Rhine river systems together with its Ponto-Caspian intermediate host Dikerogammarus villosus (David et al., Reference David, Staentzel, Schlumberger, Perrot-Minnot, Beisel and Hardion2018; Hohenadler et al., Reference Hohenadler, Nachev, Thielen, Taraschewski, Grabner and Sures2018; Vogel and Taraschewski, Reference Vogel and Taraschewski2023; note that initially, no distinction was made between P. laevis and P. bosniacus). This aligns with findings from the Weser river system, where P. bosniacus was found exclusively in D. villosus, despite extensive sampling of native gammarid species (Vogel and Taraschewski, Reference Vogel and Taraschewski2023). Such host specificity likely prevents P. bosniacus from establishing in regions lacking D. villosus, like our study area.
Variability in parasitism rates suffered by amphipods may generally be related to species-specific parasite–host relationships. In gammarids, acanthocephalan prevalence has been shown to differ not only among recognized species but also among morphologically cryptic, yet genetically highly divergent lineages (molecular operation taxonomic units, MOTUs) within, for example, G. fossarum and G. pulex (Westram et al., Reference Westram, Baumgartner, Keller and Jokela2011; Bauer and Rigaud, Reference Bauer and Rigaud2015; Zittel et al., Reference Zittel, Grabner, Wlecklik, Sures, Leese, Taraschewski and Weigand2018; Galipaud et al., Reference Galipaud, Bollache and Lagrue2019). The study sites in the present study are inhabited by five different MOTUs of G. fossarum, three of which were common and infected by Acanthocephala. None of the G. fossarum MOTUs found to be infected in our analyses were included in previous studies, and our data therefore extend the known host range of P. tereticollis and Polymorphus sp. type 1 to include these MOTUs. Unfortunately, the low parasite prevalence at sites with more than one G. fossarum MOTU precluded statistical tests for MOTU-specific infection patterns.
Temporal variation in parasite prevalence
Temporal fluctuations in acanthocephalan prevalence levels have been documented in several Acanthocephala-gammarid systems, such as P. laevis in G. balcanicus (Dudiňák and Špakulová, Reference Dudiňák and Špakulová2003) and G. fossarum (van Maren, Reference van Maren1979) and Polymorphus minutus in G. roeselii (Médoc and Beisel, Reference Médoc and Beisel2009) and G. lacustris (Spencer, Reference Spencer1974), but the patterns vary. Spencer (Reference Spencer1974) described a decrease of the prevalence of Polymorphus minutus coinciding with the emergence of juvenile amphipods (G. lacustris) in the summer months, and suggested that the reduction of prevalence estimates was due to a dilution of the population with young, not visibly infected amphipods. This explanation may also apply to what we observed at site #12 in our study. The mean weight of the amphipods collected there ranged from 18 to 27 mg (max. 57 mg for the heaviest individual) in the months when the prevalence was between 7% and 9%, but was only 12–13 mg (max. 54 mg) in the two summer months when the prevalence was below 2%, indicating that the summer samples contained more small amphipods than those collected in the spring months. Long-term monitoring in several streams is required to determine whether the observed drop in visible infections during the summer months is a general, seasonal pattern in the study region.
Intraspecific genetic diversity
We detected high genetic diversity among the small number of Polymorphus sp. type 1 individuals that were collected in this study. In one stream reach (sites # 10 and 11), each of five individuals had a different haplotype. This suggests that, despite low local abundances, a large effective population size might be maintained by the transport of parasites by their mobile avian hosts. In contrast, haplotype diversity in P. tereticollis was lower, especially in relation to sample size. Nucleotide diversity within stream systems, driven by the presence of less common haplotypes in addition to the most common one, varied independently of the level of parasite prevalence and may be related to gene flow mediated by the host fish. Most individuals of P. tereticollis shared a common haplotype that was present at all sampling sites. The predominance of a single haplotype at all sampling sites may indicate a recent, perhaps postglacial, population expansion, as has also been inferred on a larger geographic scale for P. tereticollis throughout the western Palearctic (Perrot‐Minnot et al., Reference Perrot‐Minnot, Špakulová, Wattier, Kotlik, Düşen, Aydoğdu and Tougard2018). Although some sampling sites contained private haplotypes, no geographic structure was apparent within the study region. At a European scale, the restriction of haplogroup 4 to the Styrian part of the Mur and its tributaries indicates phylogeographic structure within P. tereticollis, which is, however, diluted by the widespread occurrence of haplogroup 3 across Europe (from the United Kingdom to Slovakia, and including the present study region; Supplementary Fig. S3).
Conclusions
In conclusion, we report species-specific abundance and intraspecific genetic diversity of three species of Acanthocephala in gammarid hosts in and around an urban centre, including the first record of a recently identified cryptic species, Polymorphus sp. type 1 (Zittel et al., Reference Zittel, Grabner, Wlecklik, Sures, Leese, Taraschewski and Weigand2018), in Austria. Our data did not support a correlation between acanthocephalan infections in amphipods and urban land use. The overall low prevalence of parasites in the amphipods, along with higher values at more downstream sites, suggests that the abundance of the definitive hosts (fish and waterfowl) may be a limiting factor for Acanthocephala populations in the small streams, which may attenuate potential impacts of urbanization. While within-stream comparisons are desirable to allow some control of among-stream variation, disentangling natural and anthropogenic causes of the downstream accumulation of parasites may prove difficult in short streams, where anthropogenic disturbances are often concentrated in the downstream reaches.
Our study demonstrates the usefulness of collecting infected amphipods to obtain Acanthocephala samples suitable for, e.g. genetic analysis. Importantly, given the limited mobility of amphipods (Alther et al., Reference Alther, Fronhofer and Altermatt2021; Švara et al., Reference Švara, Michalski, Krauss, Schulze, Geuchen, Brack and Luckenbach2022; Weiss et al., Reference Weiss, Weigand and Leese2022), the Acanthocephala collected in this way represent the natal population, i.e., individuals that developed from eggs that were released in the same stream. This allows for a more spatially explicit analysis of parasite population structure than sampling from the definitive hosts, which may include individuals migrating through the study area.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0031182025100449.
Data availability statement
DNA sequences generated in this study are available at GenBank (accession numbers PV792653-PV792788). All other data used in the study are included in the submission.
Acknowledgements
The authors thank Špela Di Batista Borko for calculating the distance-from-source variable, and Stephan Koblmüller as well as two anonymous reviewers for valuable comments on an earlier version of the manuscript.
Author contributions
FG and KMS designed the study; FG, JG and KMS collected the data; FG, KMS and SR analyzed the data; FG and KMS wrote the manuscript with input from all authors.
Financial support
DNA sequencing was supported by the Biodiversitätsfonds of the Federal Ministry of Agriculture and Forestry, Climate and Environmental Protection, Regions and Water Management of the Republic of Austria, and the University of Graz. The authors acknowledge the financial support by the University of Graz for Open Access publishing.
Competing interests
None.
Ethical standards
Not applicable.








 Open access
Open access