




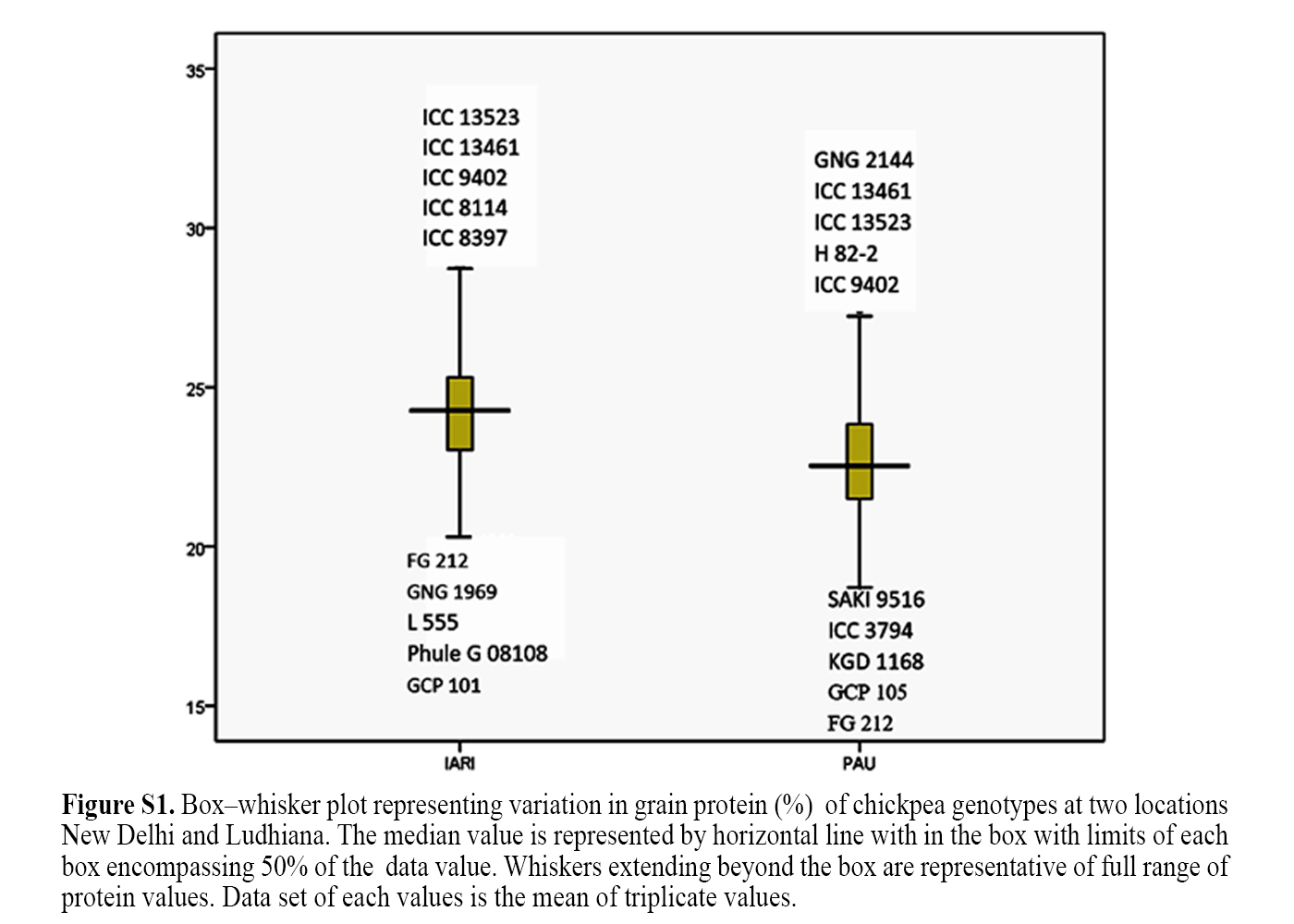
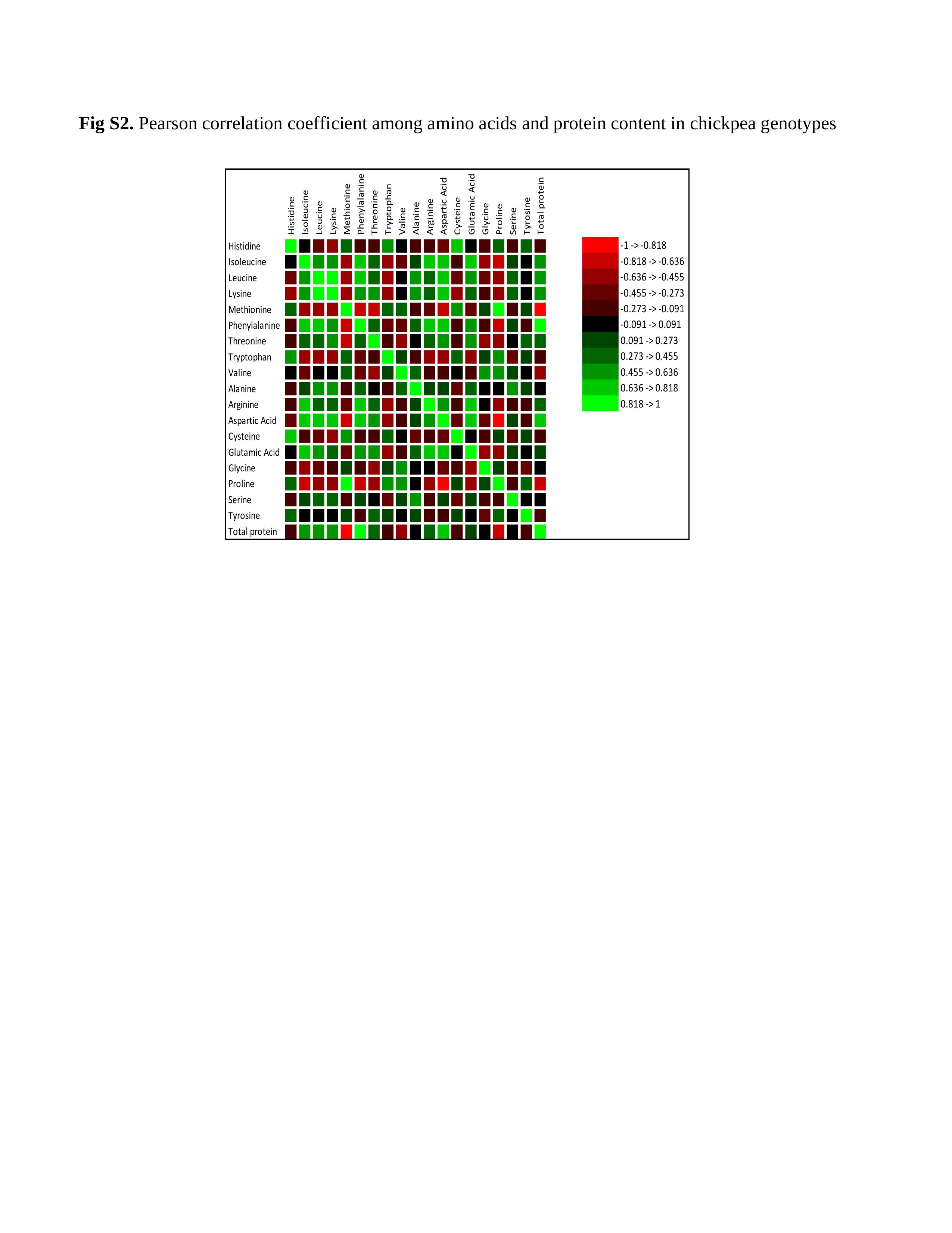
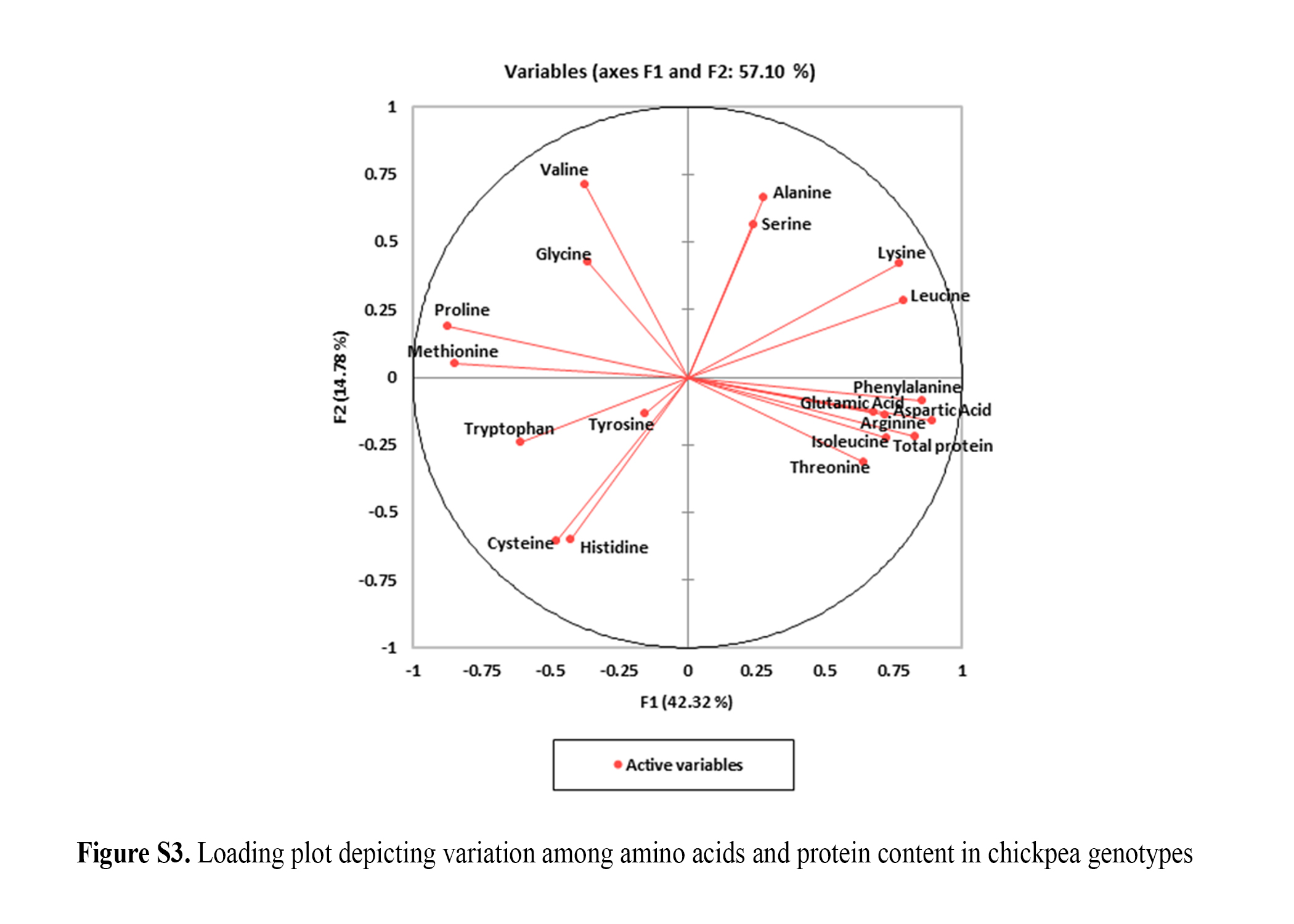

Crossref Citations
This article has been cited by the following publications. This list is generated based on data provided by Crossref.
Jindal, Aprajita
Patil, Nikhil
Bains, Aarti
Sridhar, Kandi
Stephen Inbaraj, Baskaran
Tripathi, Manikant
Chawla, Prince
and
Sharma, Minaxi
2023.
Recent Trends in Cereal- and Legume-Based Protein-Mineral Complexes: Formulation Methods, Toxicity, and Food Applications.
Foods,
Vol. 12,
Issue. 21,
p.
3898.
Abou Chehade, Lara
Tavarini, Silvia
Bozzini, Maria Francesca
Koskey, Gilbert
Caturegli, Lisa
Antichi, Daniele
and
Angelini, Luciana G.
2024.
Agronomic and Phytochemical Characterization of Chickpea Local Genetic Resources for the Agroecological Transition and Sustainable Food Systems.
Agronomy,
Vol. 14,
Issue. 10,
p.
2229.
Jha, Uday Chand
Nayyar, Harsh
Thudi, Mahender
Beena, Radha
Vara Prasad, P. V.
and
Siddique, Kadambot H. M.
2024.
Unlocking the nutritional potential of chickpea: strategies for biofortification and enhanced multinutrient quality.
Frontiers in Plant Science,
Vol. 15,
Issue. ,