



No CrossRef data available.
Article contents
A QTL allele from wild soybean enhances protein content without reducing the oil content
Published online by Cambridge University Press: 29 November 2023
Abstract
Soybean is one of the chief crops producing protein and oil for human consumption. Wild soybean, the ancestor of cultivated soybean, possesses high seed protein content; therefore, it is a valuable genetic resource that could enhance protein content in the cultivated varieties. To identify the genes responsible for increasing protein content in wild soybean, a population comprising 113 BC4F6 chromosome segment substitution lines (CSSL) was developed from a cross between soybean cultivar ‘Jackson’ and wild soybean accession JWS156-1. The CSSL population was cultivated in the field conditions for 3 years (2018, 2019 and 2020), and the seeds harvested from each line were analysed for protein and oil contents by InfraTec Nova instrument. Quantitative trait locus (QTL) analysis with 243 SSR markers identified 12 QTLs associated with seed protein, oil and protein + oil contents. Among these QTLs, qPro8 and qPro19, two major and stable QTLs for protein content, were detected on chromosomes 8 and 19, respectively. No QTL for oil content was detected in the vicinity of qPro19, indicating that qPro19 did not influence the seed oil content. The effect of qPro19 was validated using near-isogenic lines (NILs) of qPro19. By introducing the qPro19 allele from wild soybean into another soybean variety, ‘Tachiyutaka’, a BC4 line, T-678, that showed enhanced seed protein content, without reducing the seed oil content. This study implied that the qPro19 allele from wild soybean could be a potential genetic resource for breeding programmes aimed to improve soybean seed quality.
- Type
- Research Article
- Information
- Copyright
- Copyright © The Author(s), 2023. Published by Cambridge University Press on behalf of National Institute of Agricultural Botany
References
Chapman, A, Pantalone, VR, Ustun, A, Allen, FL, Landau-Ellis, D, Trigiano, RN and Gresshoff, PM (2003) Quantitative trait loci for agronomic and seed quality traits in an F2 and F4: 6 soybean population. Euphytica 129, 387–393.CrossRefGoogle Scholar
Chung, J, Babka, HL, Graef, GL, Staswick, PE, Lee, DJ, Cregan, PB, Shoemaker, RC and Specht, JE (2003) The seed protein, oil, and yield QTL on soybean linkage group I. Crop Science 43, 1053–1067.CrossRefGoogle Scholar
Clemente, TE and Cahoon, EB (2009) Soybean oil: genetic approaches for modification of functionality and total content. Plant Physiology 151, 1030–1040.CrossRefGoogle ScholarPubMed
Concibido, V, La Vallee, B, Mclaird, P, Pineda, N, Meyer, J, Hummel, L, Yang, J, Wu, k and Delannay, X (2003) Introgression of a quantitative trait locus for yield from Glycine soja into commercial soybean cultivars. Theoretical and Applied Genetics 106, 575–582.CrossRefGoogle ScholarPubMed
Cregan, PB, Jarvik, T, Bush, AL, Shoemaker, RC, Lark, KG, Kahler, AL, Kaya, N, VanToai, TT, Lohnes, DG, Chung, J and Specht, JE (1999) An integrated genetic linkage map of the soybean genome. Crop Science 39, 1464–1490.CrossRefGoogle Scholar
Diers, BW, Keim, P, Fehr, WR and Shoemaker, RC (1992) RFLP analysis of soybean seed protein and oil content. Theoretical and Applied Genetics 83, 608–612.CrossRefGoogle ScholarPubMed
Hamwieh, A and Xu, D (2008) Conserved salt tolerance quantitative locus (QTL) in wild and cultivated soybeans. Breeding Science 58, 355–359.CrossRefGoogle Scholar
He, Q, Yang, H, Xiang, S, Tian, D, Wang, W, Zhao, T and Gai, J (2015) Fine mapping of the genetic locus L1 conferring black pods using a chromosome segment substitution line population of soybean. Plant Breeding 134, 437–445.CrossRefGoogle Scholar
Kambhampati, S, Aznar-Moreno, JA, Hostetler, C, Caso, T, Bailey, SR, Hubbard, AH, Durrett, TP and Allen, DK (2019) On the inverse correlation of protein and oil: examining the effects of altered central carbon metabolism on seed composition using soybean fast neutron mutants. Metabolites 10, 18.CrossRefGoogle ScholarPubMed
Liu, D, Yan, Y, Fujita, Y and Xu, D (2018a) A major QTL (qFT12. 1) allele from wild soybean delays flowering time. Molecular Breeding 38, 45.CrossRefGoogle Scholar
Liu, D, Yan, Y, Fujita, Y and Xu, D (2018b) Identification and validation of QTLs for 100-seed weight using chromosome segment substitution lines in soybean. Breeding Science 68, 442–448.CrossRefGoogle ScholarPubMed
Liu, S, Liu, Z, Hou, X and Li, X (2023) Genetic mapping and functional genomics of soybean seed protein. Molecular Breeding 43, 29.CrossRefGoogle ScholarPubMed
Lozano-Isla, F (2021) inti: Tools and Statistical Procedures in Plant Science. R Package Version 0.1, 3, Vienna, Austria: R Foundation.Google Scholar
Lu, W, Wen, Z, Li, H, Yuan, D, Li, J, Zhang, H, Huang, Z, Cui, S and Du, W (2013) Identification of the quantitative trait loci (QTL) underlying water soluble protein content in soybean. Theoretical and Applied Genetics 126, 425–433.CrossRefGoogle ScholarPubMed
Nozawa, S, Hakoda, A, Sakaida, K, Suzuki, T and Yasui, A (2005) Method performance study of the determination of total nitrogen in soy sauce by the Kjeldahl method. Analytical Sciences 21, 1129–1132.CrossRefGoogle ScholarPubMed
Orf, JH, Chase, K, Jarvik, T, Mansur, LM, Cregan, PB, Adler, FR and Lark, KG (1999) Genetics of soybean agronomic traits: I. Comparison of three related recombinant inbred populations. Crop Science 39, 1642–1651.CrossRefGoogle Scholar
Pathan, SM, Vuong, TD, Clark, K, Lee, JD, Shannon, JG, Roberts, CA, Ellersieck, MR, Burton, JW, Cregan, PB, Hyten, DL, Nguyen, HT and Sleper, DA (2013) Genetic mapping and confirmation of quantitative trait loci for seed protein and oil contents and seed weight in soybean. Crop Science 53, 765–774.CrossRefGoogle Scholar
Patil, G, Vuong, TD, Kale, S, Valliyodan, B, Deshmukh, R, Zhu, C, Wu, X, Bai, Y, Yungbluth, D, Lu, F, Kumpatla, S, Varshney, R and Nguyen, HT (2018) Dissecting genomic hotspots underlying seed protein, oil, and sucrose content in an interspecific mapping population of soybean using high-density linkage mapping. Plant Biotechnology Journal 16, 1939–1953.CrossRefGoogle Scholar
Peng, L, Qian, L, Wang, M, Liu, W, Song, X, Cheng, H, Yuan, F and Zhao, M (2021) Comparative transcriptome analysis during seeds development between two soybean cultivars. PeerJ 9, e10772.CrossRefGoogle ScholarPubMed
Qin, P, Wang, T and Luo, Y (2022) A review on plant-based proteins from soybean: health benefits and soy product development. Journal of Agriculture and Food Research 7, 100265.CrossRefGoogle Scholar
Reinprecht, Y, Poysa, VW, Yu, K, Rajcan, I, Ablett, GR and Pauls, KP (2006) Seed and agronomic QTL in low linolenic acid, lipoxygenase-free soybean (Glycine max (L.) Merrill) germplasm. Genome 49, 1510–1527.CrossRefGoogle ScholarPubMed
Rodrigues, GM, Cardozo-Filho, L and Silva, C (2017) Pressurized liquid extraction of oil from soybean seeds. The Canadian Journal of Chemical Engineering 95, 2383–2389.CrossRefGoogle Scholar
Sebolt, AM, Shoemaker, RC and Diers, BW (2000) Analysis of a quantitative trait locus allele from wild soybean that increases seed protein concentration in soybean. Crop Science 40, 1438–1444.CrossRefGoogle Scholar
Shook, JM, Zhang, J, Jones, SE, Singh, A, Diers, BW and Singh, AK (2021) Meta-GWAS for quantitative trait loci identification in soybean. Genes Genomes Genetics 11, jkab117.CrossRefGoogle ScholarPubMed
Song, QJ, Marek, LF, Shoemaker, RC, Lark, KG, Concibido, VC, Delannay, X, Specht, JE and Cregan, PB (2004) A new integrated genetic linkage map of the soybean. Theoretical and Applied Genetics 109, 122–128.CrossRefGoogle ScholarPubMed
Song, Q, Jia, G, Zhu, Y, Grant, D, Nelson, RT, Hwang, EY, Hyten, DL and Cregan, PB (2010) Abundance of SSR motifs and development of candidate polymorphic SSR markers (BARCSOYSSR_1. 0) in soybean. Crop Science 50, 1950–1960.CrossRefGoogle Scholar
Tajuddin, T, Watanabe, S, Yamanaka, N and Harada, K (2003) Analysis of quantitative trait loci for protein and lipid contents in soybean seeds using recombinant inbred lines. Breeding Science 53, 133–140.CrossRefGoogle Scholar
Wang, S, Kanamaru, K, Li, W, Abe, J, Yamada, T and Kitamura, K (2007) Simultaneous accumulation of high contents of α-tocopherol and lutein is possible in seeds of soybean (Glycine max (L.) Merr.). Breeding Science 57, 297–304.CrossRefGoogle Scholar
Wang, W, He, Q, Yang, H, Xiang, S, Zhao, T and Gai, J (2013) Development of a chromosome segment substitution line population with wild soybean (Glycine soja Sieb. et Zucc.) as donor parent. Euphytica 189, 293–307.CrossRefGoogle Scholar
Wang, J, Li, H, Zhang, L and Meng, L (2016) Users’ manual of QTL IciMapping. The Quantitative Genetics Group, Institute of Crop Science, Chinese Academy of Agricultural Sciences (CAAS) Beijing, China, and Genetic Resources Program. CIMMYT, Mexico City, Mexico.Google Scholar
Wang, S, Liu, S, Wang, J, Yokosho, K, Zhou, B, Yu, YC, Liu, Z, Frommer, WB, Ma, JF, Chen, LQ, Guan, Y, Shou, H and Tian, Z (2020) Simultaneous changes in seed size, oil content and protein content driven by selection of SWEET homologues during soybean domestication. National Science Review 7, 1776–1786.CrossRefGoogle ScholarPubMed
Warrington, CV, Abdel-Haleem, H, Hyten, DL, Cregan, PB, Orf, JH, Killam, AS, Bajjalieh, N and Boerma, HR (2015) QTL for seed protein and amino acids in the Benning × Danbaekkong soybean population. Theoretical and Applied Genetics 128, 839–850.CrossRefGoogle ScholarPubMed
Wilcox, JR (1998) Increasing seed protein in soybean with eight cycles of recurrent selection. Crop Science 38, 1536–1540.CrossRefGoogle Scholar
Xu, W, Wang, Q, Zhang, W, Zhang, H, Liu, X, Song, Q, Zhu, Y, Cui, X, Chen, X and Chen, H (2022) Using transcriptomic and metabolomic data to investigate the molecular mechanisms that determine protein and oil contents during seed development in soybean. Frontiers in Plant Science 13, 1012394.CrossRefGoogle ScholarPubMed
Yang, Y, La, TC, Gillman, JD, Lyu, Z, Joshi, T, Usovsky, M, Song, Q and Scaboo, A (2022) Linkage analysis and residual heterozygotes derived near isogenic lines reveals a novel protein quantitative trait loci from a Glycine soja accession. Frontiers in Plant Science 13, 938100.CrossRefGoogle ScholarPubMed
Zhang, H, Li, C, Davis, EL, Wang, J, Griffin, JD, Kofsky, J and Song, BH (2016) Genome-wide association study of resistance to soybean cyst nematode (Heterodera glycines) HG Type 2.5.7 in wild soybean (Glycine soja). Frontiers in Plant Science 7, 1214.Google ScholarPubMed
Zhang, D, Lü, H, Chu, S, Zhang, H, Zhang, H, Yang, Y, Li, H and Yu, D (2017) The genetic architecture of water-soluble protein content and its genetic relationship to total protein content in soybean. Scientific Reports 7, 5053.CrossRefGoogle ScholarPubMed
Zhang, S, Hao, D, Zhang, S, Zhang, D, Wang, H, Du, H, Kan, G and Yu, D (2021) Genome-wide association mapping for protein, oil and water-soluble protein contents in soybean. Molecular Genetics and Genomics 296, 91–102.CrossRefGoogle ScholarPubMed
Zhou, Z, Jiang, Y, Wang, Z, Gou, Z, Lyu, J, Li, W, Yu, Y, Shu, L, Zhao, Y, Ma, Y, Fang, C, Shen, Y, Liu, T, Li, C, Li, Q, Wu, M, Wang, M, Wu, Y, Dong, Y, Wan, W, Wang, X, Ding, Z, Gao, Y, Xiang, H and Tian, Z (2015) Resequencing 302 wild and cultivated accessions identifies genes related to domestication and improvement in soybean. Nature Biotechnology 33, 408–414.CrossRefGoogle ScholarPubMed

Park et al. supplementary material 1
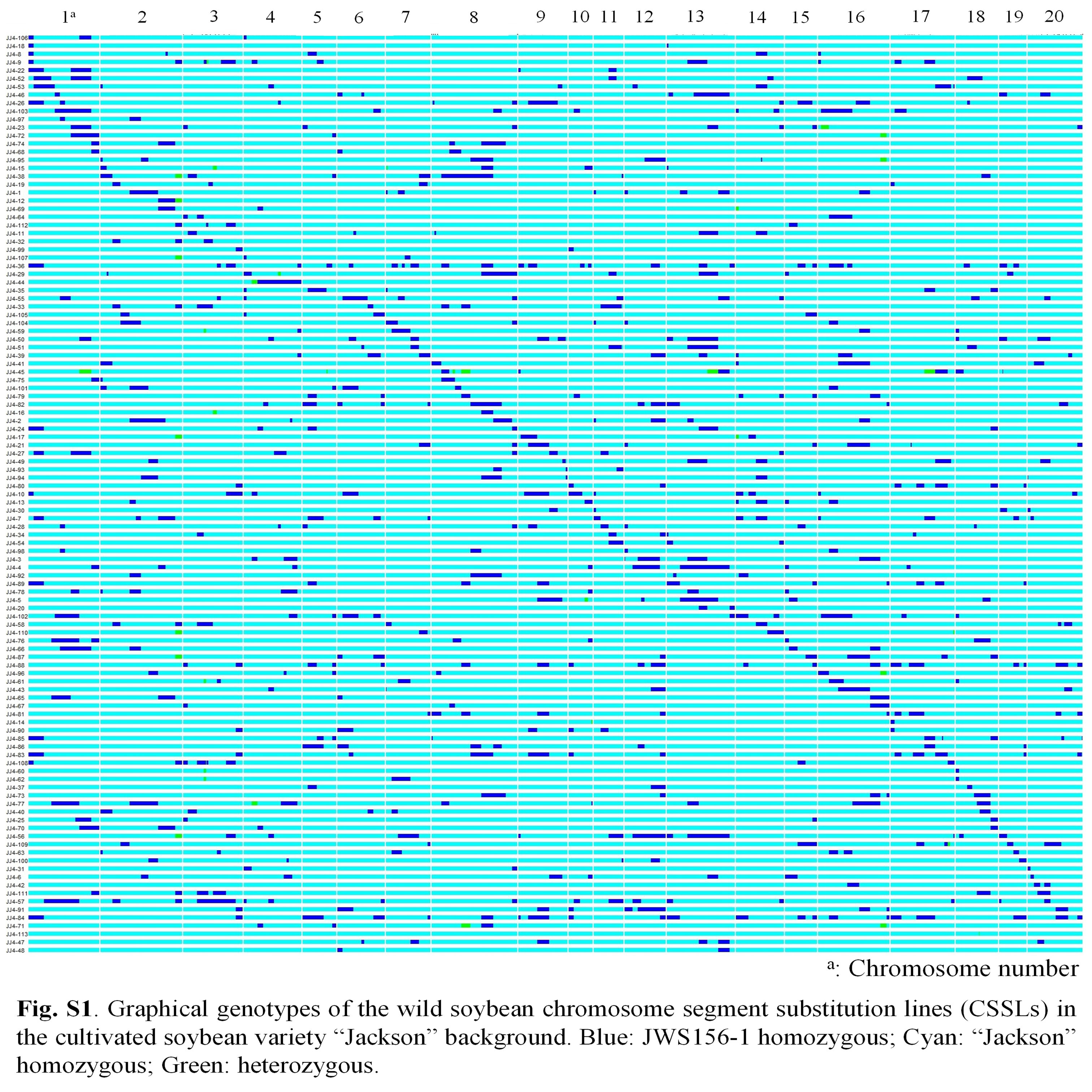
File
3.3 MB
Park et al. supplementary material 2
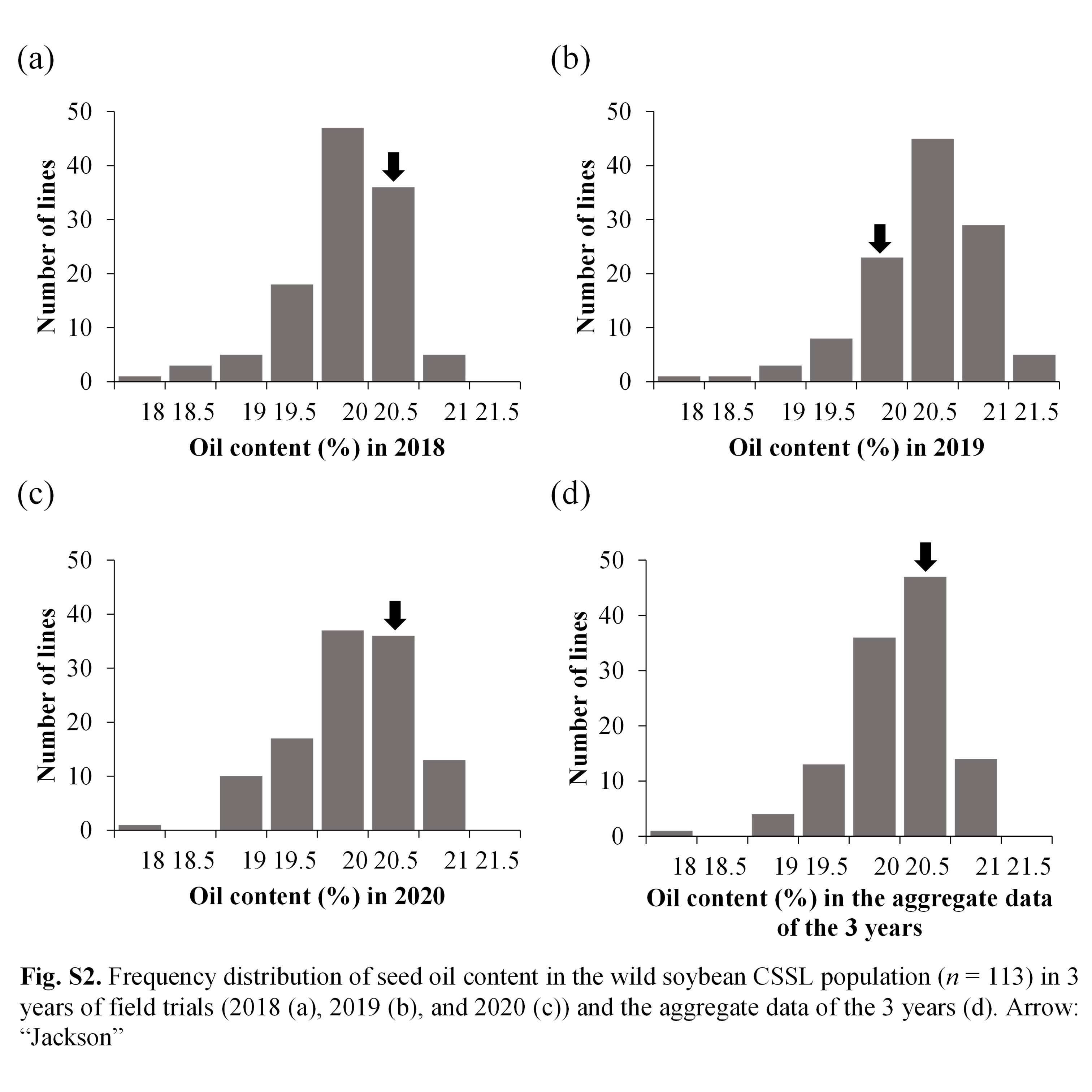
File
701.8 KB
Park et al. supplementary material 3
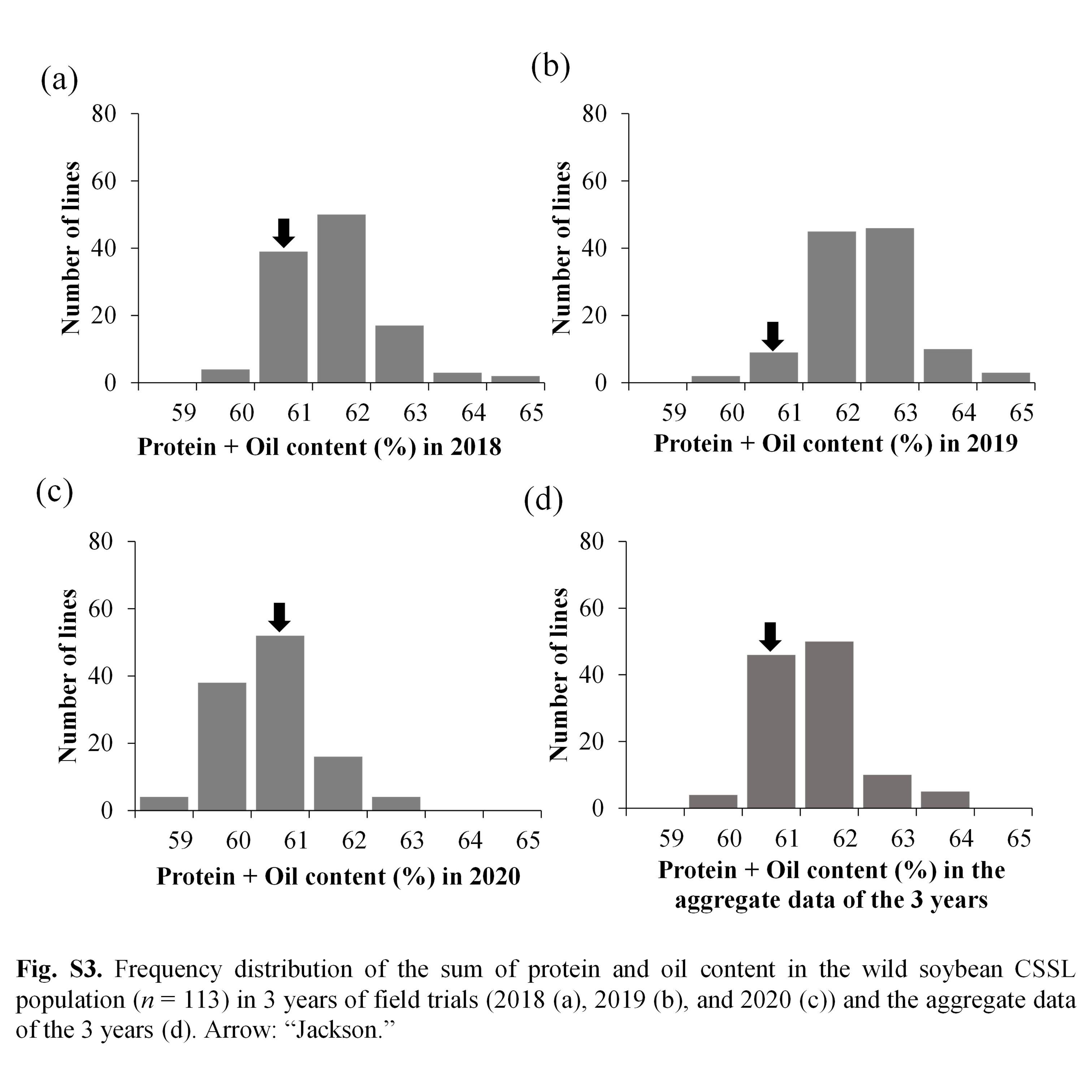
File
694.2 KB
Park et al. supplementary material 4
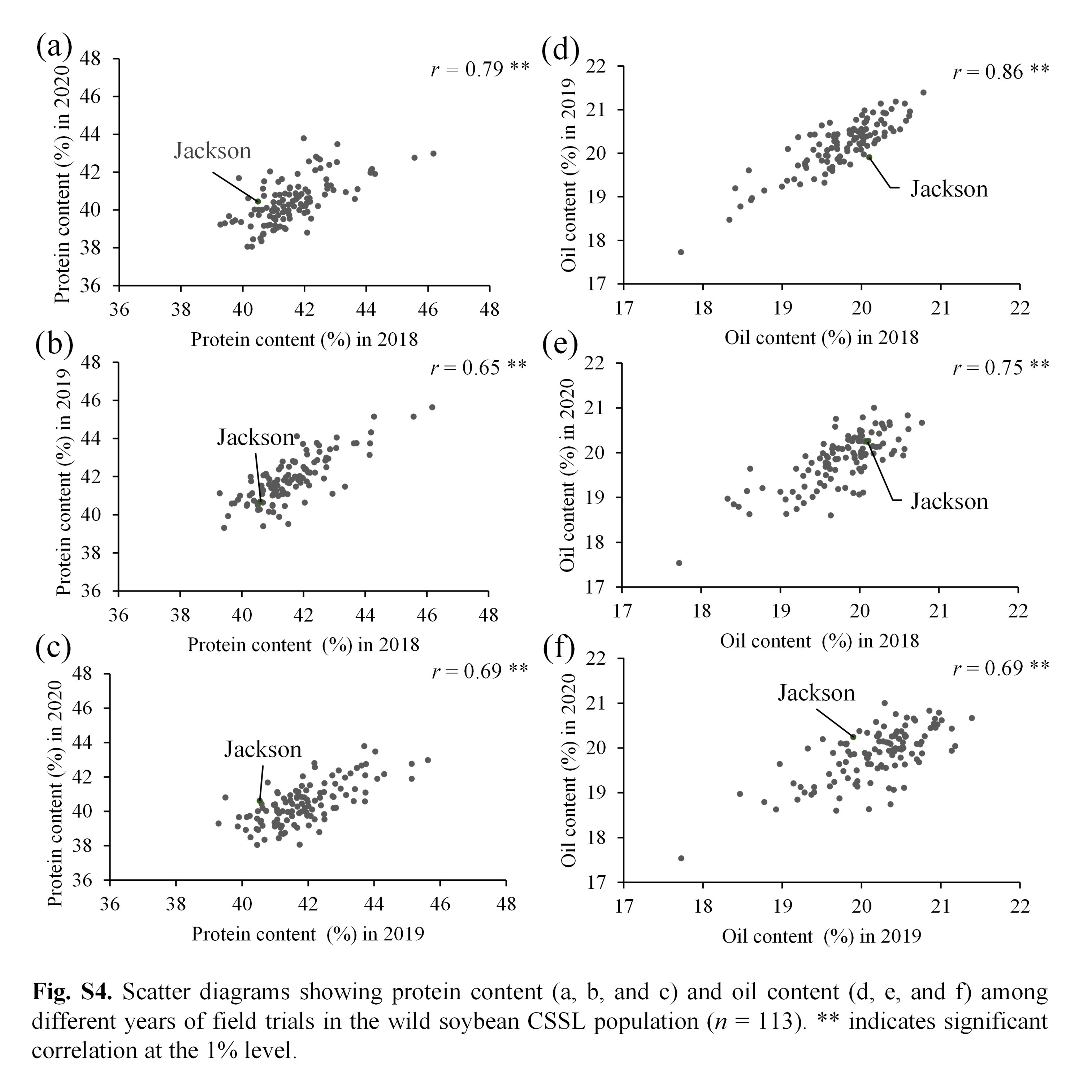
File
854.3 KB