


INTRODUCTION
The hunter-gatherer-fisher Ertebølle culture of southern Scandinavia (EBK), c. 5400–4000 cal bc, is one of Europe’s best-known Mesolithic societies. Much of this owes to a long history of research (eg, Madsen et al. Reference Madsen, Müller, Neergård, Petersen, Rostrup, Steenstrup and Winge1900; Mathiassen Reference Mathiassen1943; Troels-Smith Reference Troels-Smith1967; Brinch Petersen Reference Brinch Petersen2015; Gron & Rowley-Conwy Reference Gron, Rowley-Conwy, Piskin, Marciniak and Barkowiak2018), which has uncovered a substantial and well-studied record of late Mesolithic life. Part and parcel to this are the substantial numbers of known burials (eg, Larsson Reference Larsson1988; Brinch Petersen Reference Brinch Petersen2015). As such, the EBK archaeological and bioarchaeological record represents one of the best opportunities for understanding Mesolithic society, its structure, variability, and whether functional social differentiation was present.
Conventional perspectives paint EBK groups as territorial and as delayed-return foragers (Woodburn Reference Woodburn and Gellner1980; Reference Woodburn1982). Several burial aggregations, possibly cemeteries, are known and, if understood in the framework of the Saxe-Goldstein hypothesis (Saxe Reference Saxe1970; Goldstein Reference Goldstein, Chapman, Kinnes and Randsborg1981), represent unilineal descent groups and ownership of estates. However, application of the model to the European Mesolithic may be questionable. In particular, it is primarily directed to the issue of formal cemeteries (Morris Reference Morris1991), a concept yet to be demonstrated in Scandinavia. Critical to defining formal cemeteries is spatial separation of the living and the dead, which does not appear to apply to either site central to this paper, Strøby Egede or Vedbæk-Bøgebakken (see Meiklejohn et al. Reference Meiklejohn, Brinch Petersen, Alexandersen, Zvelebil, Dennell and Domańska1998). In both cases, the burials lay within the primary occupation (see, for instance, Brinch Petersen Reference Brinch Petersen2015).
While lithic evidence (Vang Petersen Reference Vang Petersen1984) suggests territoriality or some similar form of land tenure, social hierarchy is far from clear and differentiation is probably horizontal. Recent work in boreal forest areas shows sites with multiple burials and rich burial goods are anything but restricted to groups with tightly defined territoriality (Wood et al. Reference Wood, Higham, Buzilhova, Suvorov, Heinemeier and Olsen2013). Therefore, there are no a priori reasons to assume that differences in burial practices reflect differences in station.
The pattern of Danish Mesolithic burials allowing the study of these themes is broad, ranging from classic inhumation burials to partial burials, bone scatters, and cremation deposits. Meiklejohn et al. (Reference Meiklejohn, Babb and Hiebert2016) list 26 sites with securely dated Mesolithic burials in Denmark. Footnote 1 With few exceptions, the burial units involve one or two persons, with only two containing more: burial 19 at Vedbæk-Bøgebakken, with two adults and a child, and Gøngehusvej 7, feature N, a multiple cremation deposit with five individuals. The Dyrholmen bone scatter has at least ten individuals but without current evidence that it was intentional. In looking at children’s skeletons, the clear majority accompany an adult. In aggregate, these provide little evidence regarding the living structure of EBK residential bands and, in particular, no information regarding the daily collective activities of these Stone Age foragers.
The Strøby Egede burial is an exceptional find, an inhumation with eight individuals ranging from a 35–45 year old adult to four children aged 6 or less, of which two may be newborns. Buried together, they almost certainly died at roughly the same time and potentially from the same event. The feature is therefore an apparent ‘snapshot’ of the past, ostensibly recording with whom the individuals were at death, their relationships with one another, if there was social differentiation in how they were memorialised within the grave, and whether sex and gender were recognised consistently or independently across the age range. Footnote 2
Unfortunately, poor organic preservation has, to date, precluded successful direct radiocarbon dating, light isotope analyses of bone and dentine collagen, or aDNA analysis. Previous conclusions regarding adolescent and sub-adult biological sex were therefore presumptive. In this paper, we apply a novel acid etch-based analysis of dimorphic sex chromosome-linked tooth enamel peptides to fully resolve the sex of individuals in the grave. This is the only available method, other than aDNA, which can determine accurately biological sex in non-adults. We also, for the first time, present bioarchaeological information gathered in the 1980s and 1990s by two of us (VA and CM). We neither attempt to unpick social hierarchies within the grave, nor identification of a ‘main’ person. Instead, we investigate the relationship between age, sex, and gender in this EBK group, the relationships between those interred and, ultimately, the circumstances of their deaths. We do so by contextualising the grave within the ethnographic, archaeological, and anthropological records. By revisiting this burial, we demonstrate that an integrated approach with interdisciplinary interpretive frameworks including archaeology, anthropology, and biology permits a deeper understanding of EBK society.
THE ERTEBØLLE SOCIAL SETTING
Land tenure and sedentism
The Ertebølle is the last phase of the southern Scandinavian Mesolithic (c. 5400–4000 cal bc). Starting in the early Atlantic period, the landscape had already changed dramatically from that of the early Mesolithic. Eustatic sea-level rise (Christensen Reference Christensen and Fischer1995; Bailey et al. Reference Bailey, Andersen, Maarleveld, Bailey, Galanidou, Peeters, Jöns and Mennenga2020) resulted in the submergence of Doggerland and formation of a landscape roughly approximating that seen today in the region. While not uniform, the resulting resource spaces created a highly productive habitat mix, with distribution of a large variety of resources in concentrated areas (Paludan-Müller Reference Paludan-Müller1978) available in a predictable sequence throughout the annual cycle (Rowley-Conwy Reference Rowley-Conwy and Bailey1983). These resources could be exploited from regions occupied year-round (Andersen Reference Andersen, Milner, Craig and Bailey2007). Such developments set the stage for what Rowley-Conwy (Reference Rowley-Conwy and Bailey1983) identified as probable sedentism but which more likely amounts to habitual regional estates. Associated developments include evidence of land tenure, seen in regional styles probably indicating group areas (Vang Petersen Reference Vang Petersen1984; Reference Vang Petersen, Jensen, Sørensen and Hansen2001), and logistic resource exploitation (Rowley-Conwy Reference Rowley-Conwy and Bailey1983; Price & Gebauer Reference Price and Gebauer2005) underpinned by storage technology including ceramics (Hallgren Reference Hallgren and Knutsson2004).
The arguments for year-round site occupation or sedentism (Andersen Reference Andersen, Milner, Craig and Bailey2007), or probable sedentism (Rowley-Conwy Reference Rowley-Conwy and Bailey1983), are not definitive and are, ultimately, a question of degree. In fact, Andersen, while discussing larger shell middens, primarily on Jutland, writes that ‘(t)he larger sites have been visited several times during the annual cycle, but we still lack convincing proof of permanent and continuous whole year occupation’ (Reference Andersen, Milner, Craig and Bailey2007, 37; italics in original). This ultimately means that one of the more problematic aspects of the Saxe-Goldstein model (Saxe Reference Saxe1970; Goldstein Reference Goldstein, Chapman, Kinnes and Randsborg1981), linked to sedentism and burial, is the striking absence of burial concentrations on larger shell midden sites.
Johansen (Reference Johansen2006) and Brinch Petersen (Reference Brinch Petersen2015) both query the evidence and model of sedentism and, for Danish sites on the Øresund, the pattern of sites suggests overlying evidence of repeated short occupations while the presence of multiple sites around fjords and minimal evidence for permanent structures, as at Vedbæk and Nivå, supports a model of group movement. Additionally, burials are not spatially organised in obvious fashion. At Vedbæk-Bøgebakken and Gøngehusvej 7, both in the Vedbæk fjord, distribution of burials and hearths are highly similar (Meiklejohn et al. Reference Meiklejohn, Brinch Petersen, Alexandersen, Zvelebil, Dennell and Domańska1998). The pattern of sites could therefore fit the fission-fusion settlement pattern of hunter-gatherer groups (see, for instance, Turnbull Reference Turnbull, Harrison and Boyce1972); larger, more visible, sites represent group fusion of several smaller bands. Motivation for group fusion includes exploitation of periods of resource abundance, related feasting, and/or the opportunity for meeting potential marriage partners (see, for instance, Meiklejohn Reference Meiklejohn, Helmer, van Dyke and Kense1977; Reference Meiklejohn and Mellars1978).
While often discussed in relationship to low density hunter-gatherers, such scattered groups can produce some of the features central to the Saxe-Goldstein model, including large sites with associated burial concentrations of cemetery proportions. One example, Oleni Ostrov in Russian Karelia, provides one of the few European Mesolithic cases for a cemetery in the technical sense of a place set aside for burial, with little to no obvious evidence for the theoretically expected hierarchal social structure. The conundrum is clearly seen in the debate over the nature of this site between O’Shea and Zvelebil (Reference O’Shea and Zvelebil1984) and Jacobs (Reference Jacobs1995).
Finally, a seldom discussed issue is the population size needed to sustain a genetically boundaried group (or biological deme) and, in anthropological/demographic terms, a marriage universe. Simulation studies by Wobst (Reference Wobst1976; see also Dyke & MacCluer Reference Dyke and MacCluer1973; MacCluer & Dyke Reference MacCluer and Dyke1976; Meiklejohn Reference Meiklejohn and Mellars1978) see ‘self-sufficient mating networks’ requiring a population of at least 475 people. In practical terms, this means that, over any extended period, smaller groups tend to run out of potential mates of both opposite sex and appropriate age, in consideration of acceptable kinship categories (eg, not close relations Footnote 3 ). To date no model has been put forward that clearly encompasses both the archaeological evidence and the biological constraints stemming from the nature of extended kinship systems.
Social differentiation
Since the 1970s, the number of known Ertebølle burials has expanded substantially, particularly those variously referred to as cemeteries or burial grounds. The two largest, potentially best-suited for understanding Ertebølle social roles are Skateholm, in Skåne (Scania) southern Sweden, where work began in 1979/80 (Larsson Reference Larsson1988) and Vedbæk-Bøgebakken, Footnote 4 Sjælland (Zealand), Denmark, where work began in late 1974 (Brinch Petersen Reference Brinch Petersen2015). Vedbæk-Bøgebakken (Brinch Petersen Reference Brinch Petersen2015; Fig. 1), with 17 burial features and 22 individuals, is the best-studied Mesolithic site surrounding the Vedbæk fjord, with better organic preservation than Skateholm (eg, Eriksson & Lidén Reference Eriksson and Lidén2002). Of 12 excavated sites at Vedbæk eight have yielded human bone (Brinch Petersen Reference Brinch Petersen2015, figs 10:1 & 26:1).
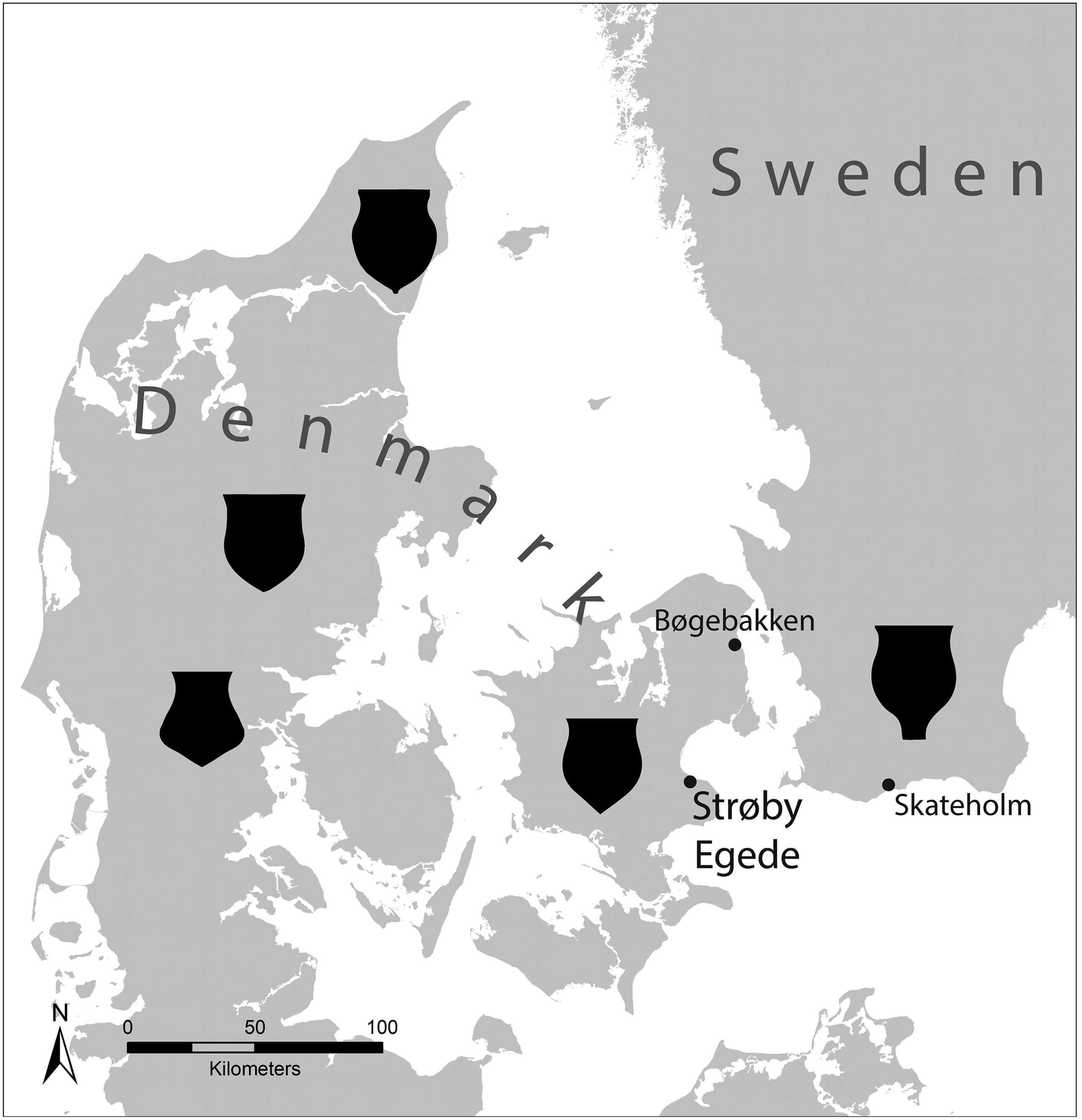
Fig. 1. Southern Scandinavia showing the location of key sites mentioned in the text, with approximate areas of Ertebølle ceramic style differences shown (after Sørensen Reference Sørensen, Kabaciński, Hartz, Raemaekers and Terberger2015 but see Jennbert Reference Jennbert2011 for more detail). Basemap data extracted from the GADM database (www.gadm.org), version 4.0.4
Known Ertebølle burials range in age from children and infants though mature adults and, among these, there are discernible differences in burial treatment. Fahlander (Reference Fahlander2012) identified differences between age classes from analysis of grave-goods and spatial distribution of burials at Skateholm. At c. 7 or 8 years children started to engage with the adult world and, by puberty, their graves were indistinguishable from those of adults. One element at Skateholm, but not consistent across Mesolithic Europe, was lack of evidence for infants buried individually; most are accompanied by an adult. To our knowledge, a full survey of Mesolithic infant burials is not available. Multiple examples are known at, for instance, Vlasac and Lepenski Vir in the Djerdap (Borić & Stefanović Reference Borić and Stefanović2004) and Moita do Sebastião in Portugal (Ferembach Reference Ferembach1974). However, in Denmark the number is very limited when compared to total number of burials. Besides sites with dated burials mentioned above (Vedbæk-Bøgebakken, Gøngehusvej 7), contextually dated cases are from Maglemosegård in the Vedbæk fjord (Alexandersen Reference Alexandersen1979), Nivågård in the Nivå fjord (Alexandersen et al. Reference Alexandersen, Meiklejohn and Brinch Petersen1999), Ertebølle on the Limfjord in northern Jylland (Madsen et al. Reference Madsen, Müller, Neergård, Petersen, Rostrup, Steenstrup and Winge1900), and Nederst on the now dry Kolindsund in east-central Jylland (Kannegaard Reference Kannegaard2016).
Using this material, some initial discussion of EBK sex, gender, and social differentiation has occurred (Meiklejohn et al. Reference Meiklejohn, Brinch Petersen, Alexandersen, Donald and Hurcombe2000) resulting in attempts to define male and female burial characteristics. Early work at Vedbæk-Bøgebakken saw the suggestion that females were buried with belts, sometimes termed girdles, adorned with teeth from multiple taxa (Albrethsen & Brinch Petersen Reference Albrethsen and Brinch Petersen1977; Brinch Petersen Reference Brinch Petersen1979; see also Price et al. Reference Price, Ambrose, Bennike, Heinemeier, Noe-Nygaard, Brinch Petersen, Vang Petersen and Richards2007). However, more broad-scale studies of EBK sex and gender based on burial goods demonstrate that exceptions to perceived patterns (Schmidt Reference Schmidt, Casella and Fowler2005) are very common and probably belie far more complex cultural attitudes than are easily characterised. Not the least of the questions is whether some burial goods may mimic those normally identified as related to sex/gender. As a clear example, Meiklejohn et al. (Reference Meiklejohn, Brinch Petersen, Alexandersen, Donald and Hurcombe2000) raised the issue of whether the linkage could reflect lineage which, of course, manifests itself through sex/gender in both matrilineal and patrilineal societies, though not intrinsically based on the sex/gender separation.
In a cross-cutting analysis of burial goods and attempting to identify social differentiation on the basis of gender, investigations found almost certain horizontal (age, sex, achievement-based) differentiation among EBK individuals (Clark & Neeley Reference Clark, Neeley, Rowley-Conwy, Zvelebil and Blankholm1987; Meiklejohn et al. Reference Meiklejohn, Brinch Petersen, Alexandersen, Donald and Hurcombe2000). If anything, this complicates things further because if achievement-based differentiation in grave-goods was common in the EBK, then differences in grave furnishing may reflect neither gender nor sex. Furthermore, Meiklejohn et al. (Reference Meiklejohn, Brinch Petersen, Alexandersen, Donald and Hurcombe2000) discuss explicitly difficulties in determining whether burial equipment marks ascribed or prescribed status. The potential for prescribed status is certainly present, as the delayed-return character of the EBK makes establishment of social hierarchies likely despite the relative dearth of evidence to its effect (Layton & Rowley-Conwy Reference Layton and Rowley-Conwy2013). Therefore, evidence of particular artefact types, artefact categories, and their abundance, even with young children including infants, cannot be definitively linked, in whole or in part, to sex and/or gender.
Additionally, sex and gender are not the same; Deaux (Reference Deaux1985, 51) provides useful definitions. Sex is biologically defined, with male and female categories based on chromosomal X and Y allele combinations, while gender is psychological and usually, but not exclusively, associated with the biological categories. The two are correlated but not mutually dependent. Gender is, at least in part, culturally constructed, being the sum total of cultural, social, and psychological human interpretations of the meaning of biological sex (Shapiro Reference Shapiro1981). Society constructs a degree of implicit and explicit, intentional and unintentional, and cultural normative attitudes towards gender. Modern North American populations publicly engender even foetuses in utero (Guignard Reference Guignard2015) and, in so doing, intrinsically confuse the gender/sex boundary by assuming that cultural construction occurs in utero. Therefore, while children are not born with gender, they must learn these attitudes, which may start at birth. Moreover, no societies are genderless. All languages include words separating boys/men and girls/women (Maccoby Reference Maccoby1988); gender is a cultural universal.
Children will self-segregate by gender (or by sex?) by age three (Maccoby Reference Maccoby1988) in modern industrialised societies, although there is no reason to suspect this will be the case everywhere. The more settled a hunting and gathering people the earlier in life gender roles develop and become entrenched, at least in part due to the fact that, with increasing sedentism, gender roles are learned more through explicit teaching by adults than through imitation of adults (Lew-Levy et al. Reference Lew-Levy, Lavi, Reckin, Cristóbal-Azkarate and Ellis-Davies2018). Among Kipsigis pastoralists of Kenya for example, despite strong gender differentiation among adults no clear preferences for gender segregation occur in children’s peer groups until middle childhood, around the age of six (Harkness & Super Reference Harkness and Super1985). Given this base, no a priori assumptions can be made about the age when gender would have been reified among EBK foragers.
A central theme in the number of studies aimed at understanding the intersection between age, sex, and social status during the Ertebølle, is the fundamental persistent difficulty with geography; comparisons are separated widely in space (Fahlander Reference Fahlander2012). This is problematic because of land tenure (see above) and strong indications that the Ertebølle was characterised by estates of varying scales (Vang Petersen Reference Vang Petersen1984; Sørensen Reference Sørensen, Hårdh, Jennbert and Olausson2007; Grøn Reference Grøn2020). Their presence is implied by the wealth of data clearly suggesting a degree of geographic variation in material culture across the landscape but the situation is complex and not straightforward to interpret. One indicator of differences between adjacent estates is seen in flake axe styles, which can vary by location in eastern Sjælland (Vang Petersen Reference Vang Petersen1984). Another is the eastern–western divide across the Storebælt over the presence and absence of particular artefact types, especially t-shaped antler axes which are rare in the east (Vang Petersen Reference Vang Petersen1984). Similarly, Limhamn axes seem not to have been characteristic of the west (Sørensen Reference Sørensen, Hårdh, Jennbert and Olausson2007). There is also some suggestion that anthropomorphic figure styles show regional variation (Płonka Reference Płonka2021). However, perhaps the clearest indicator is the variation in ceramic styles across the landscape (Jennbert Reference Jennbert2011; Sørensen Reference Sørensen, Kabaciński, Hartz, Raemaekers and Terberger2015; Fig. 1) which can be best described as variations on a common theme.
It therefore stands to reason that if there are regional differences in artefact types and styles, then there may also be regional differences in treatment of the dead (see, for instance, Drucker Reference Drucker1950), borne out by the EBK record. Grave-goods vary by location and composition and cannot be reliably correlated with specific gender in a broad sense (Meiklejohn et al. Reference Meiklejohn, Brinch Petersen, Alexandersen, Donald and Hurcombe2000). We also might expect different populations. As an example, Meiklejohn et al. (Reference Meiklejohn, Brinch Petersen, Alexandersen, Zvelebil, Dennell and Domańska1998; Meiklejohn et al. in prep.) postulated, based on statistical analysis of cranial measurements, that flake axe style zones might imply biologically separated groups on Sjælland, though population size would play a role, as noted above. A possible solution to the dilemma of how style zones and distinct populations might interact is seen in ethnographic evidence showing hunter-gatherer marriage exchange networks linking across strong cultural and linguistic boundaries; a classic example (Burch & Correll Reference Burch and Correll1972) showed long lasting marriage networks linking Athapaskan and Inuit groups in northern Alaska.
Ertebølle land tenure therefore imbues a wrinkle of doubt into discussions of larger scale patterns or commonality in cultural practice, including how gender was recognised in life and death and discussions of burial practice. This relates to clear differences in material culture and economic practices at nested scales within the Ertebølle culture area. While both are, at least in part, tied to factors over which foragers have no control (eg, availability of raw material; Vang Petersen Reference Vang Petersen1984) and environmental factors such as presence or absence of a particular species (eg, Aaris-Sørensen Reference Aaris-Sørensen1999), many are explainable only through the lens of human decision-making within the limits of culturally-constructed frameworks of propriety and habitus. Furthermore, we would expect to see more group insularity in highly productive environments, as clearly provided in many areas of southern Scandinavia, and in particular Sjælland (Paludan-Müller Reference Paludan-Müller1978; Fitzhugh et al. Reference Fitzhugh, Phillips, Gjesfjeld, Whallon, Lovis and Hitchcock2011). An argument can therefore be made that the only scale at which gender, group structure, and cultural practice should be investigated is at the most insular, within-group, scale. Cultural attitudes about these practices differ between populations, even in infancy (Drucker Reference Drucker1950).
The southern Scandinavian archaeological record therefore presents a plethora of possible avenues for analysis of burials in the context of studies regarding sex and gender (Schmidt Reference Schmidt, Casella and Fowler2005) and should theoretically represent an opportunity for understanding cultural patterns relating to these. However, evidence for regional group land tenure, and the likely fluid dynamics of any social or perimeter defense mechanisms for their maintenance, means broad-scale comparisons may, at their core, be faulty, with different cultural groups having different traditions. Added to the variation will be leakage of individuals across identified group boundaries resulting from marriage exchange, producing what Lee (Reference Lee1972) called the half-life of a population. A more insular approach would focus on individual graves, providing snapshot views of the largest number of individuals possible. Strøby Egede has, arguably, one of the best opportunities to do so in a Mesolithic setting.
MATERIALS AND METHODS
Strøby Egede
In the summer of 1986 a mass grave was discovered during construction of a carp pond along the bank of the river Tryggevælde Å, c. 2 km south-east of where it currently empties into Køge Bugt, the embayment south of modern Copenhagen (Brinch Petersen Reference Brinch Petersen1987; Reference Brinch Petersen1988; Reference Brinch Petersen1990a; Reference Brinch Petersen1990b; Fig. 1). Ultimately, eight individuals were identified, four on each side of the grave when oriented north to south, placed head to foot. Initial analyses, centred on burial arrangement and grave-goods (Brinch Petersen Reference Brinch Petersen1988; Reference Brinch Petersen1990b) interpreted the burial as with four females arranged on the southern end, four males to the north. The number of individuals, their age range, and what must have been the uncommon circumstance(s) under which they died, make this an exceptional find in Mesolithic Europe. Unfortunately, as noted above, collagen preservation issues prevented direct radiocarbon dating. Some years later, a single grave containing a male individual was discovered only a few metres from the mass grave, confirming rumours about more graves in the area. Footnote 5 The associated settlement has characteristic middle EBK artefact types (Stationsvej projectile points), and an archaeological date estimate of c. 4700 bc (Brinch Petersen Reference Brinch Petersen1990a; Reference Brinch Petersen1990b). To address sex, gender, relationships, and cause of death it is necessary to know the ages and biological sexes of those interred.
Skeletal assessment
To unpack various aspects of the Strøby Egede burial, it is pivotal to estimate the age at death of the individuals. A primary issue for analysis was the lifting of the feature en bloc, with a view towards museum display, following initial in situ excavation (Brinch Petersen Reference Brinch Petersen1990b). Other than the crania and mandibles of the older individuals the remains could neither be manipulated nor removed from the block for close examination. This prevented full analysis, including inspection of bone surfaces, especially of the pelvic and vertebral regions, and prevented a full biometric study. In retrospect, Strøby Egede epitomises a dilemma of modern archaeology, pitting the scientific against the public side.
Four individuals were initially identified, a number that expanded to eight following additional work at the site and after transportation to the Preservation Laboratory, Nationalmuseet, Copenhagen, where further work included examining the burial unit from below by inverting it. Following this the burial, in toto, was moved to Køge Museum for permanent display (Brinch Petersen Reference Brinch Petersen1987). Initial descriptions were based on observations by Verner Alexandersen and J. Balslev Jørgensen during excavation and cleaning of the material (Brinch Petersen Reference Brinch Petersen1988; Reference Brinch Petersen1990a; Reference Brinch Petersen1990b), when the bones, other than crania and mandibles, were uncovered but not removed from their matrix. The discussion below centres on later work, in 1992, by one of us (CM), assisted by Jeffrey M. Wyman (University of Winnipeg), made within the confines of the exhibit case at Køge Museum (further recent input was by VA; see supplementary information SI). Given limitations of working in the exhibit case, the inventories in the SI are incomplete; much of the material could not be fully observed.
Peptide-based sex determination
To fully confirm the biological sex of the individuals we applied a tooth-enamel acid etch-based sex determination procedure (Stewart et al. Reference Stewart, Gerlach, Gowland, Gron and Montgomery2017). This involves a less-destructive acid-based etch to recover chromosome-linked dimorphic peptides from tooth enamel useful in determining the biological sex of human skeletal remains.
RESULTS
The Strøby Egede individuals
The burial, enclosed within a c. 1 × 2 m pit, has bodies aligned in two directions. The southern group, A, B, C, and F, comprise an adult, an adolescent, and two children, while the northern group, D, E, G, and H, are an adult and three children (Fig. 2). The adults and the adolescent were better preserved than the children. The information below has four sources, the initial papers written after excavation (Brinch Petersen Reference Brinch Petersen1987; Reference Brinch Petersen1988; Reference Brinch Petersen1990a; Reference Brinch Petersen1990b), work by CM (with J.M. Wyman) at the Køge Museum in 1992, an unpublished paper on the dentition written by VA in 1997, and further analysis by VA for this paper, based on photos taken by staff at the Køge Museum. We stress again the limited possibilities for analysis that followed lifting of the burial en bloc and its installation at the Køge Museum following preservation.

Fig. 2. The Strøby Egede burial. Throughout the text individuals are referred to by their letter designation. North to the right. See Brinch Petersen (Reference Brinch Petersen1990b)
The summaries below, given in order of letter identification (see Fig. 2), are to provide context for the tooth-etching described below, the primary focus of this paper. Fuller descriptions of the material and associated data are found in the supplementary Appendix (SI) to this paper.
Strøby Egede A
The only adult and most complete individual in the southern group had a single associated flint blade. Skull and postcranial skeleton are largely complete with most bones preserved. Initial identification as female is confirmed by dental etching. Age evaluation, including dental markers, suggests a range of 35–45 years. Estimated stature from humerus, femur, and tibia is 146–160 cm, consistent with the sex diagnosis. The thick cranial vault, reaching 10 mm in places, is noted in other Danish Mesolithic samples, including Bøgebakken. The dentition shows features associated with heavy attrition and age (for further information here and for further individuals see supplementary Appendix SI).
Strøby Egede B
The only older child, also from the southern group, was associated with ten red deer tooth pendants. It includes a partial skull, mandible, and largely complete postcranial skeleton. Given the age, no skeletal sex assessment was possible, though identified as female from grave-goods (Brinch Petersen Reference Brinch Petersen1987), confirmed by dental etching. Dental eruption and long bone length suggest an age of 7–8 years.
Strøby Egede C
The only adolescent, and from the southern group, was associated with a single bone hairpin and ten red deer tooth pendants. Parts of the vault and face of a partial skull, a mandible, and most of the postcranial skeleton were identified. Though identified as female by Brinch Petersen (Reference Brinch Petersen1987), age made that diagnosis suspect, though now confirmed by dental etching. Initially assessed as c. 18 years old (ibid.), work in 1992, focused on dental eruption and other markers, gives a range of 14–17 years. Stature, consistent with age and sex, is estimated at c. 150 cm.
Strøby Egede D
The only northern group adult and best-preserved individual had seven associated flint blades, one red deer antler axe, and a bone knife. Visible are a largely complete skull and postcranial skeleton (parts of the left side are obscured in the display). Initially identified as male (Brinch Petersen Reference Brinch Petersen1987; Reference Brinch Petersen1990a), this is confirmed by dental etching. Initially assessed as c. 30 years old (Brinch Petersen Reference Brinch Petersen1987; Reference Brinch Petersen1990a), more recent work suggests a range of 20–25 years. Long bone stature is estimated at 165–175 cm, similar to taller males at Bøgebakken.
Strøby Egede E
The oldest of three children in the northern group was accompanied by a flint knife in two pieces. Though identified in Brinch Petersen (Reference Brinch Petersen1988), the cranium was not visible in 1992. The 1988 figure shows this as the largest and, by extension, oldest of the younger children (see Figs 2 & 5). The postcranial skeleton was reasonably complete though partially obscured in the display.
Strøby Egede F
The only younger child in the southern group, accompanied by a pendant from the anterior tooth of a boar and a roe deer foot. Few remains were recovered. Sex identification of this and the next two individuals was inferred from position; no tooth etching was possible from the three.
Strøby Egede G
One of two infants in the northern group and the least complete individual from the site, with two truncated flint blades in association. A limited set of postcranial remains were identified.
Strøby Egede H
The second infant from the northern group had an associated boar tooth pendant and seven red deer tooth pendants. In slightly better condition than G, there are postcranial remains but only tooth germs were found from the cranium.
Tooth etch-based sex determination
Enamel from five teeth was etched (Table 1), for individuals A–E. We were unable to etch the infants, F–H; no tooth remains were recovered from G and the tooth germs from F and H could not be etched.
TABLE 1: details of teeth etched and results

The etch procedure followed a slightly modified version of Stewart et al. (Reference Stewart, Gerlach, Gowland, Gron and Montgomery2017), with two etches per tooth, at the same location. However, unlike Stewart et al. (Reference Stewart, Gerlach, Gowland, Gron and Montgomery2017), the first etch was not discarded but processed as below, in case the enamel thickness was breached, reaching the dentine. This proved unnecessary and ultimately the first etches were not analysed. Teeth were first cleaned of obvious surface contamination using a dental tool, to identify a surface suitable for etching (eg, free of cracks, fully mineralised, etc). Approximately 60μL of 0.5M HCl was placed in the open cap of a 0.2 mL Eppendorf tube, forming a convex meniscus protruding above the cap surface. The first etch was performed by lowering the enamel surface onto the acid, maintaining contact for two minutes. A C18 resin ziptip pipette tip (EMD Millipore ZTC18S096) attached to a 4–10μL pipette set to 10μL was first conditioned 3× with 100% acetonitrile, and 3× with 0.1% formic acid. The etch solution was pipetted 10× through the conditioned ZipTip binding the enamel proteins to the resin. The ZipTip was then washed six times in 0.1% formic acid. The peptides were then eluted by setting the adjustable pipette to 4μL and drawing a 4μl 60% acetonitrile/0.1% formic acid buffer through the ZipTip 10×, leaving the last draw in a 0.2 mL Eppendorf tube, which was lyophilised.
The second etch from each tooth was analysed, with the exception of individual A, etched three times with only the third giving a result (Fig. 3). The nanoLC-MS method is as described in Stewart et al. (Reference Stewart, Gerlach, Gowland, Gron and Montgomery2017), with addition of an inclusion list for masses 440.2233 m/z and 540.2796 m/z; corresponding to Ser-Met(oxidised)-Ile-Arg-Pro-Pro-Tyr (from AMELY) and Ser-Ile-Arg-Pro-Pro-Tyr-Pro-Ser-Tyr (from AMELX), respectively. Sex was determined by visualising the reconstructed ion chromatogram at 1 PPM mass accuracy for these two peptides and confirmed with correct retention time, a correct charge state of 2 in the full MS and an accompanying MS/MS spectrum matching predicted fragment ions. All samples were anonymised and analysed blind.

Fig. 3. Reconstructed ion chromatograms at 2 ppm mass accuracy for the m/z of 440.2231 and 540.2796 for the peptides SM(ox)IRPPY ([M+2H]+2, AmelY) and SIRPPYPSY ([M+2H]+2, AmelX), respectively from Individual A. The presence of only the AmelX isoform confirms this individual as female
Given the presence of only the X chromosome-linked isoform of amelogenin, individuals A, B, and C are identified as female (Figs 3 & 4), and those with both X and Y chromosome-linked isoforms of amelogenin, D and E, as male (Fig. 4). Summary results are given in Table 1.

Fig. 4. Tooth etch-based sex determination for individuals B, C, D, E. Italicised letters indicate the individual from which the data derive
LESSONS FROM A SINGLE GRAVE
How do we tie these results and the archaeological record to anthropological theory, particularly in a Stone Age context? Because gender, status, and relationships are culturally constructed we can only access these aspects of culture through decision-making, found in evidence such as what to include with a buried child, what to feed the child, and with whom to bury the child in death. The key is in recognising when and how society perceives gender, status, and social relationships and the age when these become reified in material culture. This requires determination of the age categories that EBK groups and, in particular, this EBK group, recognised as significant. The Strøby Egede multiple inhumation is remarkable in its ability to provide information, given the number of individuals, their spread of ages at death, their biological sex, their grave-goods (Table 2), and the likelihood they were bound by a particular set of cultural norms through a discrete time-period (or the norms of whoever buried them). In so doing, its analysis minimises potential misinterpretation arising from ongoing changes in human culture (see Eerkens & Lipo Reference Eerkens and Lipo2005).
TABLE 2: summary data regarding the strøby egede burial (partially from brinch petersen Reference Brinch Petersen1988). * columns reflect work undertaken in 1992

The Strøby Egede burial contains eight individuals, oriented roughly on a north–south axis (Brinch Petersen Reference Brinch Petersen1988; Fig. 5). Based on tooth etches and bioarchaeological assessment, the males were placed head to toe from north to south in the grave, the females south to north (Fig. 5). All had at least one type of grave-good or tool (Table 2). Individuals of both sexes had associated blade knives. Two females had tooth belts, males did not. Two infants had a tooth at their head, probably attached to a hood, one did not. Infant G, without the tooth bead, was buried with two knives. Both adults had blade knives, individual A with one, individual D with seven (Brinch Petersen Reference Brinch Petersen1988).
The three infants have no sex diagnosis derived from direct analysis. Those with tooth pendants at their heads (F & H) that were probably affixed to a hood, are presumptively male and female, based on burial arrangement. No clearly engendered goods were with them. But were they arranged with those of the same sex? Which was more important, burial with close kin or with one’s own sex? Placement of individual H, not being held, suggests association with individual A, placed between her legs. Therefore, not holding them is an argument for individual H being male given the burial arrangement. Or is the relationship with individual E, with their heads closely together? The other two infants were clearly being held by individuals D and B/C respectively.
Therefore, in this EBK band, burial with a knife was not restricted to a single sex and burial with knives was not related to gender. Burying an infant with a tooth at their head related to neither sex nor gender; the burial arrangement suggests this was done for both sexes. Given the presence of only one adult male, his burial with a stippled antler axe could easily be related to other types of horizontal differentiation, with no other male buried with this type of artefact. Individuals B and C, both female, were buried with a tooth belt (or girdle), seen in other female burials on Sjælland (Albrethsen & Brinch Petersen Reference Albrethsen and Brinch Petersen1977; Brinch Petersen et al. Reference Brinch Petersen, Alexandersen, Vang Petersen and Christensen1979; Price et al. Reference Price, Ambrose, Bennike, Heinemeier, Noe-Nygaard, Brinch Petersen, Vang Petersen and Richards2007). No Strøby Egede males were buried with similar adornment, nor was the infant in the south, found with the other females. Tooth belts were likely engendered in this instance with individual F not having one, though both B and C did. None of the sex-determined males nor those arranged as male had this adornment. Given her determined sex, possession of a tooth belt by a 6–9 year old (B) shows enculturation of gender-identity by as early as 6 years old in girls of this EBK band.
So, at what age was gender recognised in this Ertebølle group? The evidence suggests that, at birth, gender was probably not recognised though sex was, and that, by 6 years of age, a gender associated primarily with female biological sex was recognised. With biological sex clearly known and important since birth, and despite it not being possible to determine the sex of the infants, juveniles were arranged with adults of the same sex. But sex is not gender. Individual E, a 4–6 year old boy, was arranged with the males, but not associated with apparently engendered grave-goods, making it unclear whether this individual was perceived as a boy, a male, both, or neither. These subtle differences reflect the intersection of culture and biology. Given the clear evidence of engendered grave-goods associated with individual B and lack of clear gender difference among infants, gender enculturation in this EBK band therefore appears to have taken place between birth and c. 6 years of age.
WHAT HAPPENED?
Did everyone die at the same time for the same reason? In modern hunter-gatherer societies, childhood mortality rates are around 50% (Volk & Atkinson Reference Volk and Atkinson2013), similar to that of archaeological hunter-gatherers (Johnston & Snow Reference Johnston and Snow1961). Childhood deaths are therefore not exceptional. But it is not strictly necessary that the infant deaths were for the same reasons as the adults. The adults may have died together with the infants killed or dying as a result. For example, after death of a mother in Greenlandic Inuit society, infants would often be killed and buried with them (Ammitzbøll et al. Reference Ammitzbøll, Ry Andersen, Andersson, Bodenhoff, Eiken, Eriksen, Foged, Ghisler, Gotfredsen, Hansen, Hart Hansen, Jakobsen, Jørgensen, Kobayasi, Kromann, Lyberth, Lyneborg, Mikkelsen, Møhl, Møller, Myhre, Pedersen, Prause, Sebbesen, Svejgaard, Thompson, Frølund Thomsen and Vanggaard1991). This would still place infant deaths soon after the older individuals but means that the aetiology need not be identical. However, this does not explain the deaths of the non-adults older than infancy, possibly suggesting a cause of death similar to the adults.
Comment is needed here on Terberger et al. (Reference Terberger, Kotula, Jungklaus and Piezonka2021, 685), arising from discussion of the positioning of grave 1 at Groß Fredenwalde in Brandenburg, discovered in 1962 and referred to by Brinch Petersen (Reference Brinch Petersen1988). They ask whether Strøby Egede ‘might be the result of more than one event’, comparing it to work since 2012 on the Groß Fredenwalde material. Given that our discussion below accepts the Strøby Egede burial as a single event, some justification is necessary.
While recognising the issue raised above, we feel that its direction to Strøby Egede is incorrect. Similarities exist between the sites, most clearly that both were discovered accidentally, during construction. However, the work immediately after differs substantially in the two. Neither the work at Groß Fredenwalde in 1962, nor that at Neuwied-Irlich discovered in 1953 and also noted by Terberger, resemble that undertaken at Strøby Egede in 1986. Though a first-order resemblance exists between figures 7a and 7b of Terberger et al. (Reference Terberger, Kotula, Jungklaus and Piezonka2021, 682, 683) and portions of the Strøby Egede find, resemblance of the cleaned version of the latter, after work at the Nationalmuseet, is not to the work in 1962 at Groß Fredenwalde, but to that after 2012, when the true nature of the site became clear. The situation at Strøby Egede shown in Brinch Petersen (Reference Brinch Petersen1988) differs profoundly from the Groß Fredenwalde pattern in Terberger et al. (Reference Terberger, Kotula, Jungklaus and Piezonka2021), figure 8. The intricate interlaying of individuals and lithics at Strøby Egede may not represent burial of all eight individuals in a single day but is certainly consistent with placing them into an open pit over a period measured in days rather than weeks or months, let alone the years as now understood for Groß Fredenwalde. Three items from Strøby Egede provide clear support for a single, even if extended, event. First, no evidence for more than one pit being dug was found during the work prior to lifting the structure en bloc. Secondly, and supporting this, individual D, apparently the last interred individual, was pressed up on the side of the pit rather than on top of the others, as would be the case in a later burial. In other words, the individual was fit into an existing pit with the others. Thirdly, individual E, the 4–6 year old child, lies with parts both overlying and underlying its neighbours, A, B, C, and H, a complex interlayering of individuals rather than the layered ‘pack of cards’ pattern seen in separate but overlapping burials in one location.
Finally, we emphasise that this is not an isolated burial but lies within an extended site including other burials, not unlike the pattern at Vedbæk and other inlets on the Øresund. A second, clearly unrelated, Ertebølle burial with grave-goods was later found within a few metres during clearing of land prior to building a neighbouring house. It remains unstudied. The overall site lies below the current suburban development.
Depending on the society, the most likely cause of death in modern hunter-gatherers is illness or, more rarely, violence (Gurven & Kaplan Reference Gurven and Kaplan2007; Hill et al. Reference Hill, Hurtado and Walker2007). The age and sex composition at Strøby Egede, if due to violence, is in line with raiding (Keeley Reference Keeley1996). Such attacks, often while victims are vulnerable as while sleeping, result in the death of mixed sex and age groups. In contrast, deaths are disproportionately male in warfare (Keeley Reference Keeley1996). However, no evidence of fatal trauma was found at Strøby Egede at any stage of analysis. The cranial lesion of Individual D is clearly healed and from an earlier event, comparable with the healed cranial injury on the Gøngehusvej 7 adult female, feature CÆ (Brinch Petersen et al. Reference Brinch Petersen, Alexandersen and Meiklejohn1993). The evidence is not similar to that seen at, for example, LBK Herxheim (Orschiedt et al. Reference Orschiedt, Häußer, Haidle, Alt and Buitrago-Téllez2003) or Globular Amphora Koszyce (Schroeder et al. Reference Schroeder, Margaryan, Szmyt, Theulot, Włodarczak, Rasmussen, Gopalakrishnan, Szczepanek, Knopka, Jensen, Witkowska, Wilk, Przybyła, Pospieszny, Sjögren, Belka, Olsen, Kristiansen, Willerslev, Frei, Sikora, Johannsen and Allentoft2019). What other possibilities are there? Prehistoric hunter-gatherer population numbers were likely too low to sustain endemic infectious pathogens (Inhorn & Brown Reference Inhorn and Brown1990). Diseases persisting in small modern hunter-gatherer populations were introduced recently (Wirsing et al. Reference Wirsing, Logan, Micozzi, Nyamwaya, Pearce, Renshaw and Schaefer1985). This point, reinforced by biomedical and ethnographic literature (Weiss Reference Weiss2001), attests to the linkage of epidemic diseases to agriculture and especially animal domestication. The earliest evidence for the plague in Scandinavia, for example, falls in the late Neolithic (Rascovan et al. Reference Rascovan, Sjögren, Kristiansen, Nielsen, Willerslev, Desnues and Rasmussen2019).
So, could disease have come from adjacent groups? Contact between Ertebølle groups and LBK farmers to the south is clear (eg, Fischer 1982), and Strøby Egede, dated to c. 4700 cal bc, clearly post-dates the earliest LBK in north-central Germany. Moreover, where widespread deaths from infectious disease occur in modern hunter-gatherers they tend to involve frequent and habitual interactions with much larger populations (Gurven & Kaplan Reference Gurven and Kaplan2007), an unlikely scenario. Nevertheless, there is the possibility of a virgin soil epidemic, exposure to a previously unknown infectious disease. Historical examples include depopulation of North America and Australia after first contact (Crosby Reference Crosby1976; Campbell Reference Campbell1983). However, our understanding of EBK/LBK contact limits this possibility at Strøby Egede.
We might also imagine sporadic zoonotic infection from close interactions with hunted animals (Weiss Reference Weiss2001) or implicate microbial pathogens that evolved with humans. However, these are typically low-morbidity illnesses where individuals live for some time and infection rates are limited. Here, we have eight dead individuals of all ages who apparently died at roughly the same time. Furthermore, many communicable diseases disproportionally affect the very young and/or very old (Hamborsky et al. Reference Hamborsky, Kroger and Wolfe2015), which, despite some exceptions, again makes disease unlikely. Residential groups of hunter-gatherers dependent on aquatic resources usually involve c. 18 individuals (Binford Reference Binford2001; Kelly Reference Kelly2013). If correct, a disease killing nearly half of the group would exceed the morbidity of smallpox in historical periods (Blake Reference Blake1953) and be simultaneous. Infectious disease is therefore unlikely.
Other possibilities include the debilitating effect of intractable or habitual diarrhoea. Non-exclusive breastfeeding can cause gastrointestinal distress in very young children (GBD 2018), and allo-mothering is known to represent a significant proportion of early infant nursing ethnographically (Hewlett Reference Hewlett and Hewlett2014). Also possible is Salmonella or Giardia infection, known in hunter-gatherers who had dogs and hunted wild game (Gracey Reference Gracey1992). However, presence of several adults plus the infants renders this unlikely. Mortality in these cases is usually substantially higher in adults than in children (ibid.).
Could poisoning have occurred? Many foods can poison, including shellfish, but ethnographically recorded populations were usually well aware of this and avoided dangerous taxa (Moss Reference Moss1993). However rare, paralytic shellfish poisoning can happen with regularity in northern waters, including Europe (McCollum et al. Reference McCollum, Pearson, Ingham, Wood and Dewar1968; Prakash et al. Reference Prakash, Medcof and Tennant1971), and ‘red tide’ has been mentioned in the Mesolithic literature (eg, Fontanals-Coll et al. Reference Fontanals-Coll, Subirá, Marín-Moratalla, Ruiz Ventura and Gibaja Bao2014). There is no reason to suspect species commonly consumed in the Mesolithic (eg, Mytilus sp.) (Andersen Reference Andersen, Milner, Craig and Bailey2007) and present in the Øresund (Strelkov et al. Reference Strelkov, Katolikova and Väinolä2017). Vectors of toxins (Burrell et al. Reference Burrell, Gunnarsson, Gunnarsson, Clarke and Turner2013) might not be to blame. Poisoning is rapid in onset (Backer et al. Reference Backer, Fleming, Rowan, Baden, Hallegraeff, Anderson and Cembella2003) and can be fatal (2–14% with no medical care; Kao Reference Kao and Falconer1993). Even higher morbidity (up to 50%) can occur in children (Rodrigue et al. Reference Rodrigue, Etzel, Hall, De Porras, Velasquez, Tauxe, Kilbourne and Blake1990). Toxins causing this type of poisoning are concentrated most often in bivalve shellfish and not affected by food preparation methods including cooking (Backer et al. Reference Backer, Fleming, Rowan, Baden, Hallegraeff, Anderson and Cembella2003). This is a possibility, particularly if the Strøby Egede burial represents a small proportion of attendees at, for example, a large feast.
Could a toxin be to blame? Botulism is profoundly deadly (Dolman Reference Dolman1960), and well known ethnographically, ethnohistorically, and historically in the northern hemisphere. The British Columbia coast, whose pre-contact coastal populations have been compared to the Scandinavian late Mesolithic (eg, Layton & Rowley-Conwy Reference Layton and Rowley-Conwy2013) is a North American hotspot. Caused by an anaerobic bacterium normally present in many basic food sources exploited during the late Mesolithic, including seals and salmon, its growth to toxic levels is found in various traditional foods, including smoked salmon, other types of preserved fish, and preserved seal flippers. Most relevantly, fatal outbreaks from fish consumption are recorded from early 20th century Denmark (Jensen & Hahnemann Reference Jensen and Hahnemann1959), usually from consuming fish and marine mammal products stored incorrectly in anaerobic conditions. Without modern medical treatment it is one of the most lethal toxins known, with a morbidity rate of 50–60% (Dembek et al. Reference Dembek, Smith and Rusnak2007). In addition, being a toxin, it is unaffected by cooking or processing. Ethnographically, these types of poisoning (albeit unconfirmed as botulism) have occurred and have killed groups of eight individuals (Stefansson Reference Stefánsson1913, 32) and whole families (Parnell Reference Parnell1934).
An accident is also possible and, most obviously, a boating accident. Other examples exist for Strøby Egede type demographic profiles including ethnohistoric cases. A 1909 case from western Greenland recorded a probable umiaq (skin boat) accident with seven dead: two adult males, an adult female, and four female children (Hansen Reference Hansen2008). A better-known example involves the eight Thule-Inuit buried in 15th century ad rock tombs at Qilakitsoq in western Greenland (Andreasen et al. Reference Andreasen, Gulløv, Hart Hansen, Lyberth and Tauber1991; Hart Hansen et al. Reference Hart Hansen, Meldgaard and Nordqvist1991); aDNA analyses revealed six female adults and two male infants representing three maternally unrelated lineages (Gilbert et al. Reference Gilbert, Djurhuus, Melchior, Lynnerup, Worobey, Wilson, Andreasen and Dissing2007). EBK boats were certainly large enough to carry eight, especially with several infants. The larger canoe at Tybrind Vig is estimated to have accommodated 6–8 individuals (Andersen Reference Andersen1987).
Falling through ice on a frozen waterway is another possibility. In modern-day Alaska falling-through-the-ice accidents are common and, in about a third of occurrences, are fatal. Essentially all ages are affected by this type of accident, with up to a half-dozen people recorded in single incidents. The most common location for these sorts of incidents was while crossing rivers (Fleischer et al. Reference Fleischer, Melstrom, Yard, Brubaker and Thomas2013).
From the above, the most likely scenarios appear to be accident or poisoning, with violence apparently excluded and infectious disease unlikely. No matter what killed those interred, it appears to have happened quickly and been unrelated to age, health, or sex. The above presents an opportunity for understanding those that died, allowing the application of anthropological models of with whom hunter-gatherers spend their time, live and/or travel, and the relationships they share with those around them.
HYPOTHETICAL RELATIONSHIPS
Unpacking Strøby Egede group relationships in the absence of aDNA data is a hypothetical exercise. However, even with good preservation (Gilbert et al. Reference Gilbert, Djurhuus, Melchior, Lynnerup, Worobey, Wilson, Andreasen and Dissing2007), aDNA cannot fully rectify the relationships between non-related individuals. This is where anthropology and the ethnographic record can help. Interpretation also relies on thorough consideration of what might have befallen those interred (see above). If, for example, the burial resulted from an accident, we may be looking at a travelling family group. A poisoning event, however, may contain members of a larger residential group or even the larger local group if poisoning took place during, for example, a feast.
Were the individuals related, and how? Bio-anthropology, cultural anthropology, and archaeology can offer possibilities. Question one is whether and how they could have been related. Ethnographic evidence shows that any given hunter-gatherer will be genetically related to c. one quarter of those in their band, by marriage to c. one half, and will have no relationship to the remaining quarter (Hill et al. Reference Hill, Walker, Božičević, Eder, Headland, Hewlett, Hurtado, Marlowe, Wiessner and Wood2011).
The Strøby Egede individuals have a broad age spread, from infants to c. 40. Of the infants, two appear to be newborns, G and H, interestingly both in the northern burial group. The third, F in the southern group, is apparently c. six months older, ruling out triplets but not twins. There are two possible maternal combinations: of three mothers with one child each or one with twins G and H, the other with the older F. In modern populations, age at menarche is c. 12–13 years (Parent et al. Reference Parent, Teilmann, Juul, Skakkebaek, Toppari and Bourguignon2003), so individuals A, C, and/or D could have been parents of the infants. The children could have been unrelated, cousins, or half-siblings.
Question two is how these individuals could have been related. Even without aDNA it may be possible to infer relationships based on age, sex, and expected group composition, based on the ethnographic record. Should we expect a single family to have the observed age distribution? Ethnographic-based hunter-gatherer demographic data need viewing with caution because of biases introduced by short-term observation of long-term and dynamic social processes (Kelly Reference Kelly2013). A residential group size of c. 18 individuals for hunter-gatherers dependent on aquatic resources has been suggested (Binford Reference Binford2001; Kelly Reference Kelly2013), meaning 3–4 families of 4–5 individuals (Kelly Reference Kelly2013). It is therefore reasonable to posit that the Strøby Egede group could fit more than one pattern. While possibly members of the same immediate family, they could also represent individuals from three unrelated families of the same residential group. Under this framework, however, at least some individuals must be kin-related.
Age uncertainty in the adults makes understanding relationships harder. It matters greatly if individual A was 35 years of age, 45, or somewhere in-between. The situation for individual D seems different, since the range appears to be only half a decade. However, and regardless of age, individuals A and D could have been partners, though an older female raises significant questions. However, if A was 45, she could easily have been D’s mother. The question of whether she could also be the mother of one of the infants depends on her actual age within the posited range. With the birth of a last child in natural human populations occurring at c. 40 years old in available ethnographic hunter-gatherer records (te Velde & Pearson Reference te Velde and Pearson2002; Kelly Reference Kelly2013), the probability is much higher at 35 than at 45.
Figure 6 synthesises the above and attempts to unravel possible relationships. Polygamy is possible as, ethnographically, the practice ranges from rare to common in hunters and gatherers (Layton et al. Reference Layton, O’Hara and Bilsborough2012). Such a potential situation is illustrated by (1). It is also possible that three families from a residential group are represented, illustrated by (2). We must also consider how many generations are represented if the burial represents an extended family. Relationships (1) and (3) posit two generations, as does (2). Relationship (4) posits three.

Fig. 5. The biological sex of the Strøby Egede interment. Hatching: female; crosses: male; grey: unknown. North to the right. See Brinch Petersen (Reference Brinch Petersen1990b)

Fig. 6. Kinship diagrams illustrating possible relationships between the Strøby Egede individuals. Individual scenarios are numbered. Grey individuals represent absent mating partners. The infant Individual F is c. 6 months older than the other two infants and therefore cannot be their sibling
There are no easy answers but understanding the relationships between individuals is key to unlocking what befell them. Throughout life, hunter-gatherers share most social interactions with kin, though this decreases with age (Migliano et al. Reference Migliano, Page, Gómez-Gardeñes, Viguier, Dyble, Thompson, Chaudhary, Smith, Strods, Mace, Thomas, Latora and Vinicius2017). Only in adulthood could interactions with non-kin exceed those with kin and, even then, would focus on a few non-kin individuals or friends. Infants are held for most of the day in hunter-gatherer groups (Hewlett et al. Reference Hewlett, Lamb, Leyendecker and Schölmerich2000) so, whatever occurred, if accidental, happened when the infants were probably being held. In some ethnographic instances a variety of carers may look after an infant but in most hunter-gatherers infants are carried by a parent or close relative (Kelly Reference Kelly2013). However, available data indicate that individuals other than an infant’s mother only carry them 20–50% of the time (Hewlett Reference Hewlett1991). If the higher end was typical for Strøby Egede, then it is statistically possible that none of the infants was being held by their mother when they died.
Ethnographic time allocation studies also provide indication of when mixed-sex and mixed-age groups might be together (Johnson Reference Johnson1975; Munroe et al. Reference Munroe, Munroe, Michelson, Koel, Bolton and Bolton1983) but underscore the fact that the proportion of time spent on a specific activity may or may not differ between sexes, depending on the activity. The age of individuals also affects time spent in particular activities. Answering questions regarding whether there were engendered differences in these activities in EBK society is in its early stages but evidence from dentition, for example, shows some tasks primarily undertaken by women, evidenced in tooth wear, with others undertaken by men (Alexandersen Reference Alexandersen1988). Given the age and sex spread, it is unlikely that the Strøby Egede individuals died during a gender-specific activity.
So why were the Strøby Egede individuals apparently arranged by sex? The obvious question relates to the infants, with no assigned sex independent of associated grave-goods. Binary arrangements are not uncommon in prehistory. Corded Ware and early Bronze Age cemeteries in central Europe typically placed individuals on their sides facing south, with females oriented to the east and males to the west (Strouhal Reference Strouhal1978; Turek Reference Turek2017). However, such arrangements are not typical of the Scandinavian Mesolithic (eg, Larsson Reference Larsson1988; Brinch Petersen Reference Brinch Petersen2015). Ethnographic evidence also shows clear-cut distinctions are not always the rule, even where some differences occur. For example, Inuit burials, though not clearly binary in burial orientation, show some male/female differences (Crass Reference Crass and Rautman2000). These examples illustrate that differentiation between sexes, and likely between genders, lies on a spectrum and a potentially fluid spectrum at that. The fact that the individuals were apparently arranged by sex, despite dying most probably in a non-gender specific cultural context, underscores that sex and gender were meaningful categories of social differentiation within this group, even in childhood.
CONCLUSION
The Strøby Egede burial is exceptional in Mesolithic Europe and has not been afforded the comprehensive treatment deserved. This paper centres on applying one of the latest techniques of sex determination for understanding the burial, while working within the preservation limitations at the site. If anything, the aggregate data underscore the complexity of the human culture at Strøby Egede. In this case knowing more shows how little we actually know. The very presence of conflicting lines of information shows that there are both concrete differences in treatment of individuals based on sex but lack of engendered grave- goods for all cases, showing that the relationship between gender, sex, status, kin relationships, and age is not clear-cut. This is to be expected because these relationships largely encompass the breadth of inter-group social differentiation in human cultures (Sorokin Reference Sorokin and Stills1968).
If anything, the eight individuals placed within the grave show the diversity of EBK methods of intra-group social differentiation and extend the diversity of burial practice found in the EBK and, to a degree, in the preceding Kongemose (Meiklejohn et al. Reference Meiklejohn, Brinch Petersen, Alexandersen, Zvelebil, Dennell and Domańska1998). They also show the nested hierarchy of social differentiation seen in other forms of material culture across the Ertebølle (eg, Vang Petersen Reference Vang Petersen1984; Jennbert Reference Jennbert2011) extended to the day-to-day social interactions within individual residential groups. This underscores the fact that simple bifurcation of social roles, as evidenced by grave-goods, does little to advance our understanding of what is an alien social milieu. It is only through interdisciplinary considerations of the possibilities that human cultural complexity can be recognised.
Acknowledgements
Funding for this research was provided by the Leverhulme Trust (Grant RPG RPG-2019-253). Work on the Strøby Egede material by Christopher Meiklejohn occurred during work with the Vedbæk Project, directed by Erik Brinch Petersen, with funding from 1987 to 1999 supported by a series of research grants from the Social Sciences and Humanities Research Council of Canada (SSHRCC). Support for Jeffrey Wyman fell within these grants to CM. CM would like to thank Erik Brinch Petersen and Niels Lynnerup for assistance in checking details during the writing of this paper. Lastly, thanks are owed to Kristen Hopper for help with the map.
SUPPLEMENTARY MATERIAL
To view supplementary material for this article, please visit https://doi.org/10.1017/ppr.2022.7










 Open access
Open access