


The East Flanders Prospective Twin Survey (EFPTS) was started in July 1964 at Ghent University, Department of Obstetrics, by Robert Derom and Michel Thiery, a twin himself. At present, it is hosted in ‘Twins’, a nonprofit Association for Scientific Research in Multiple Births and is now partly funded by the University of Ghent (Belgium), ‘Twins’ and the University of Maastricht (the Netherlands).
Fifty-five years later, the EFPTS is still active, with current research conducted in close collaboration with the Centre for Environmental Sciences of Hasselt University (Belgium) and the Departments of Psychiatry and Neuropsychology and Complex Genetics of Maastricht University (the Netherlands).
The main aims of EFPTS are:
(1) the determination of the prevalence of multiple births in a well-defined geographic area and the recording of the obstetrical (e.g., duration of pregnancy, pregnancy and birth complications, birth weight, induction of ovulation, placental weight and insertion of the cord) and obstetrico-pediatric outcomes (e.g., intrauterine growth, congenital malformations, perinatal and infant morbidity and mortality);
(2) the investigation of the causes of the multiple pregnancy and the origin of twinning;
(3) the influence of the prenatal environment on the investigated traits on later age, such as behavior, learning and school problems, intelligence, postnatal somatic growth, sexual maturation, blood pressure, stress (sub)clinical depression and psychotic symptom. Understanding how the perinatal period shapes the human epigenome and which genomic regions are sensitive to environmental influences will allow us to distinguish a healthy start of a new life from an unfavourable start;
(4) the determination of the genetic predisposition of the investigated phenotypes and the role of the environment (with special emphasis on the prenatal environment), both individual-specific and common environmental influences.
Sample Characteristics
The inclusion criteria are the following: all multiple births in the Province of East Flanders, Belgium (14,000 births per year) from whom at least one of the children, live- or stillborn, weighs 500 g or more or, when birth weights are unknown, the gestational age is at least 22 weeks. The specific methodology of this survey includes: the determination of the zygosity of each multiple birth with near certainty to certainty through examination of the placental membranes and vascular anastomoses, blood groups and DNA fingerprints (if necessary), the collection of medical data from gynecologists and neonatologists, and follow-up of the multiples through one of the studies and/or questionnaires.
Contact with the families of the multiples is maintained by an annual newsletter, a TWIN hotline for the public and the parents of twins with psychological, educational, medical or practical problems, and the organization of different evening meetings four times a year. These services for families with multiple births are a joint venture of EFPTS and the Association for Scientific Research in Multiple Births (Twins) and are mainly based on the work of volunteers. The newsletters are published on the EFPTS website (www.twins.be), and more recently a Facebook account was started to share information on a more informal level.
The EFPTS wants to respect the privacy of participants (parents and children) in accordance with the Belgian law of August 22, 2002 on patient rights and the General Data Protection Regulation (EU) 2016/679 of April 27, 2016. Therefore, the registry protocol will be submitted in 2019 to the ethical committee of all participating hospitals in East Flanders. Ghent University hospital will act as the coordinating center. This change means that an informed consent needs to be signed by all future mothers to confirm her acceptance of the registration of the obstetrico-pediatric data and the collection of a biopsy of the placenta.
Between 1964 and 2019, more than 10,000 twin-pairs and 250 triplet sets were registered and investigated (see Table 1). The EFPTS sample is representative of the total twin population of East Flanders with more than 95% registered multiple births since 1990 (Loos et al., Reference Loos, Derom, Vlietinck and Derom1998). Presently, more than 3000 twin-pairs have been enrolled in more than 10 major follow-up studies. Table 2 gives an overview of the most important follow-up cohorts of the registry.
Table 1. The number of twin-pairs in the EFPTS born between 1964 and 2018 by zygosity, chorion type and sex

Table 2. Overview of most important follow-up cohorts of EFPTS
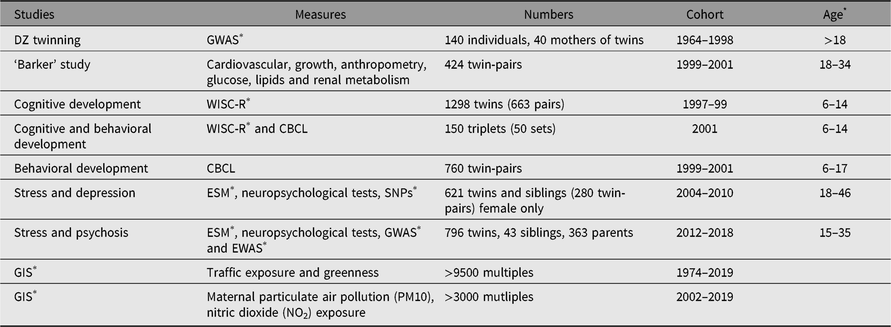
* Age: age at measurement; ESM = experience sampling method; EWAS = epigenome-wide association study; GIS = Geographic Information System; GWAS = genome-wide assocation study; SNP's = single nucleotide polymorphisms; WISC-R = Wechsler Intelligence Scale for Children–Revised
Types of Twins According to Zygosity and Chorionicity
Four groups of twins can be distinguished according to the zygosity and the number and structure of the placental membranes (see Figure 1).
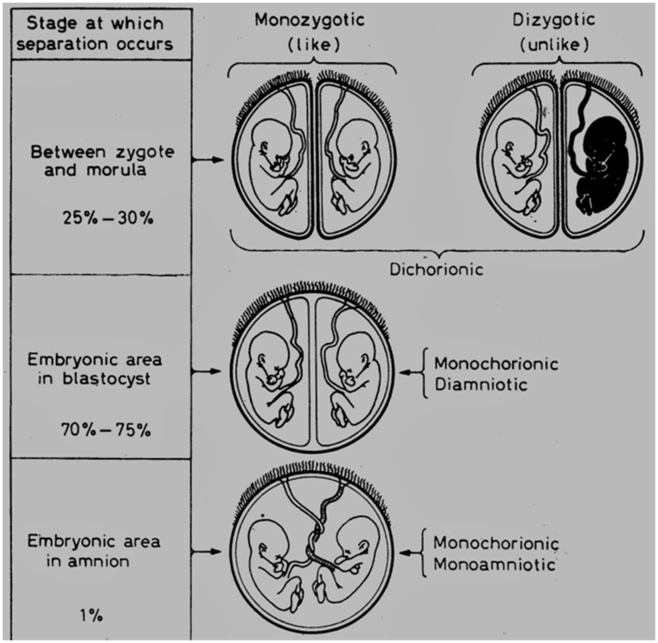
Fig. 1. Placentation in twins (Morison, Reference Morison1963).
In dizygotic (DZ) twins, who develop from the fertilization of two ova, each embryo develops within its own membranes: all DZ pairs are therefore dichorionic. In contrast, all monozygotic (MZ) twins arise from a single fertilized ovum. At some stage between the fertilization and the formation of the embryonic disk, the formative material divides into two parts, each giving rise to a complete embryo. Traditionally, it is thought that the timing of division of the fertilized egg determines chorionicity and amnionicity. If the division occurs at an early stage (before the fourth day after conception), each embryo of the resulting MZ pair will have a separate set of membranes. The two embryos are dichorionic and, in this respect, resemble DZ twins. If, however, the division of the ovum is delayed until the blastocyst has formed (between day 4 and day 8 after conception), the two embryos will share a single chorionic membrane but develop within two separate amniotic sacs. Such pairs are monochorionic–diamniotic. Exceptionally, the division of the formative material may be delayed until the embryonic disk separates from the cavities that subsequently form the amnionic and chorionic sacs (after implantation into the endometrium). When this happens, the embryos will both share a single chorionic and amnionic sac. They are monochorionic–monoamniotic (Bulmer, Reference Bulmer1970). Both members of a MZ monochorionic pair always share one placenta, which not only results in competition for a limited food supply but also enables vascular anastomoses between the circulations of the two fetuses.
EFPTS Findings
The main aim of the Developmental Origin of Health and Disease hypothesis is to link early-life risk factors with specific chronic diseases in later life, including those of cardiovascular and neurodevelopmental nature. As early-life risk factors can originate during gestation, EFPTS is ideally designed to identify some of these risk factors. The storage of placental tissue for decades and the access to birth addresses of all registered twins and triplets give us the opportunity to retrospectively study factors such as birth weight, preterm birth and the maternal exposure to ambient air pollutants as causal risk factors for different (sub)clinical phenotypes. Furthermore, stored placental biopsies allow us to look at molecular markers such as telomere length, mitochondrial DNA content and genomewide methylation patterns in association with these different phenotypes.
To further illustrate the scope of our studies, some of the most important results are reported.
Perinatal Outcome and Timing of Twinning
The MZ monochorionic–monoamniotic twin-pairs are predominantly girls (Derom et al., Reference Derom, Vlietinck, Derom, Van den Berghe and Thiery1988). The MZ monochorionic–diamniotic twin-pairs have a higher mortality rate than dichorionic MZ and DZ twin-pairs. If the division takes place after the eighth day, the mortality risk increases even more dramatically (Loos et al., Reference Loos, Derom, Vlietinck and Derom1998).
After standardization for gestational age, the birth weight of twins is mainly determined by maternal factors, especially the mother’s genetic makeup, and to a lesser extent by the chorion type and the genes of the fetuses and their common environment (Vlietinck et al., Reference Vlietinck, Derom, Neale, Maes, van Loon, Derom and Thiery1989).
Since the mid-1980s, there has been a real explosion in the rate of multiple births, primarily caused by the increasing administration of ovulation stimulating agents and the use of other assisted reproduction techniques (ART). EFPTS was the first to draw attention to the importance of this epidemic. This increase in the rate of multiple pregnancies represents an important public health problem because of the rate of very preterm births and very low birthweight infants in the twin population (Derom et al., Reference Derom, Derom, Vlietinck, Maes and Van den Berghe1993). More recently, there has been a steep decrease in the rate of higher order multiple births and twins because of a selective feticide (selective abortion of one or more embryos or fetuses) policy and the single embryo transfer in in vitro fertilization (IVF; De Neubourg & Gerris, Reference De Neubourg and Gerris2003).
The frequency of zygotic splitting is higher after artificial induction of ovulation than after naturally occurring ovulation, which is of fundamental unexplained biological importance (Derom et al., Reference Derom, Vlietinck, Derom, Van den Berghe and Thiery1987). Among MZ twins, born after the use of assisted reproduction, the ratio of monochorionic to dichorionic pairs is higher compared with spontaneous twins. A greater proportion of MZ twins is observed with clomiphene citrate as the sole treatment compared with other ovulatory drugs (Derom et al., Reference Derom, Leroy, Vlietinck, Fryns and Derom2006).
The assumption that the division of the zygote occurs stepwise later in, respectively, dichorionic MZ, monochorionic–diamniotic and monochorionic–monoamniotic pairs, has been demonstrated to be highly probable by studying X-inactivation within MZ female pairs: X-inactivation is totally symmetrical in monochorionic–monoamniotic pairs, almost symmetrical in monochorionic–diamniotic pairs and asymmetrical in dichorionic MZ pairs (Chitnis et al., Reference Chitnis, Derom, Vlietinck, Derom, Monteiro and Gregersen1999; Monteiro et al., Reference Monteiro, Derom, Vlietinck, Kohn, Lesser and Gregersen1998).
The belief that discordant handedness in MZ twins represents mirror-imaging is mythical: the often observed higher frequency of left-handedness in twins is confirmed but seems to be independent of zygosity and chorion type (Derom et al., Reference Derom, Thiery, Vlietinck, Loos and Derom1996).
In opposite-sex twins, the length of gestation and the birth weight of the male co-twin are influenced by his female co-twin and not the other way around (Loos, Derom, Eeckels et al., Reference Loos, Derom, Eeckels, Derom and Vlietinck2001).
Twins conceived through ovulation induction, IVF or intracellular sperm injection (ICSI) are at significantly elevated risk of preterm birth and, to a lesser extent, of low birth weight. The observed risks associated with assisted reproduction are dampening down by predominantly DZ twinning following assisted reproduction and by differential maternal characteristics (Verstraelen et al., Reference Verstraelen, Goetgeluk, Derom, Vansteelandt, Derom, Goetghebeur and Temmerman2005).
In recent decades, gestational age decreased in a linear fashion from 1964 to 2007 in a similar way for MZ and DZ twins. Changes in birth weight depended on gestational age. The decrease in gestational age and change in birth weight in twins are sources of concern, especially for very preterm twins, for whom birth weight decreased (Gielen et al., Reference Gielen, van Beijsterveldt, Derom, Vlietinck, Nijhuis, Zeegers and Boomsma2010).
Under the assumption that the spontaneous DZ twinning rate is a sensor of fecundity, our population-based data after age-adjustment indicate a stable ‘high’ fecundity for the East Flanders population (Derom et al., Reference Derom, Gielen, Peeters, Frijns and Zeegers2011).
A monochorionic placenta remains an excellent predictor of monozygosity and monozygosity diagnosis based on the structure of the fetal membranes can be considered accurate in almost all instances. In a consecutive series of 2006 monochorionic pairs and 3156 opposite-sex twins, all but one of the monochorionic twins were of the same sex and all but one of the opposite-sex twins were dichorionic (Table 1). Birth weight discordancy within MZ twin-pairs, however, cannot be used as an indicator of chorionicity (Gielen et al., Reference Gielen, Derom, Derom, Vlietinck and Zeegers2009).
Placental telomere length decreases during the third trimester of gestation of live-born twins by approximately 25% and is influenced by parity, indicating that telomere shortening may play a role in aging of the placenta (Gielen et al., Reference Gielen, Hageman, Pachen, Derom, Vlietinck and Zeegers2014).
In twins, a risk group for low birth weight and fetal growth restriction, a significant proportion of fetal growth restriction could be attributed to in utero exposure to particulate air pollution. Air pollution is associated with small for gestational age and birth weight in moderate-to-late preterm born twins. The within-pair difference in birth weight increases with higher air pollution exposure during the last month of pregnancy (Bijnens et al., Reference Bijnens, Derom, Gielen, Winckelmans, Fierens, Vlietinck and Nawrot2016).
Cognitive and Behavioral Development
A significant effect of chorion type on the heritability of two IQ subtests was found: the monochorionic twins resembled each other more than the dichorionic MZ twins on the subscales of Arithmetic and Vocabulary. The effect accounts for, respectively, 14% and 10% of the total variance (Jacobs et al., Reference Jacobs, Van Gestel, Derom, Thiery, Vernon, Derom and Vlietinck2001). X-inactivation, however, is not likely involved as no significant interaction was found between sex and chorionicity (Peerbooms et al., Reference Peerbooms, Wichers, Jacobs, Kenis, Derom, Vlietinck and Rutten2010).
Lower birth weight is associated with greater child problem behavior. The greater the degree of birth weight discordance within twin-pairs, the greater the degree of discordance for child problem behavior. This finding held for both MZ and DZ pairs. Therefore, it is unlikely that a shared environmental or genetic variable contributes to both birth weight and child problem behavior. These results suggest that lower birth weight represents a causal risk factor for child problem behavior (van Os et al., Reference van Os, Wichers, Danckaerts, Van Gestel, Derom and Vlietinck2001; Wichers et al., Reference Wichers, van Os, Danckaerts, Van Gestel, Derom and Vlietinck2001).
Twins with lower birth weight and cord knots had lower IQ scores. The etiology of IQ is largely distinct from that of birth weight and cord knots, and nonshared environment may influence the observed relationships (Antoniou et al., Reference Antoniou, Fowler, Thiery, Southwood, van Gestel, Jacobs and Zeegers2013).
Cardiovascular and Metabolic Phenotypes
An adverse prenatal environment during twin pregnancies has small but permanent effects on health in adult life: adult body composition, blood pressure, glucose metabolism and renal function have part of their origins in utero, but they are programmed through different prenatal environmental influences. Furthermore, the prenatal environment seems to program men and women in a different way (Gielen et al., Reference Gielen, Pinto-Sietsma, Zeegers, Loos, Fagard, de Leeuw and Vlietinck2005; Loos, Beunen, Fagard et al., Reference Loos, Beunen, Fagard, Derom and Vlietinck2001; Loos, Fagard, Beunen et al., Reference Loos, Fagard, Beunen, Derom and Vlietinck2001). The satiety signal leptin, which regulates food intake and energy expenditure, may act as a growth-promoting signal during fetal development, and the leptin receptor could have a possible role in explaining the inverse relationship between birth weight and the development of metabolic diseases in adulthood (Souren et al., Reference Souren, Paulussen, Steyls, Loos, Stassen, Gielen and Zeegers2008).
Growth during infancy is associated with birth weight and gestational age. From 0 to 1 month, environmental factors are most important for growth, whereas genetic factors become more important over time. This is a first step in identifying age windows for future counseling and interventions on the effects of accelerated growth (Touwslager et al., Reference Touwslager, Gielen, Mulder, Gerver, Zimmermann, Fowler and Zeegers2011).
DNA methylation variability at growth-related imprints does not contribute to overweight in MZ twins discordant for body mass index (Souren et al., Reference Souren, Tierling, Fryns, Derom, Walter and Zeegers2011).
Severe intrauterine growth differences observed within MZ twins are not associated with long-lasting DNA methylation differences in cells composing saliva. Additionally, our results indicate that uneven cell-type composition can lead to spurious results and should be addressed in epigenomic studies (Souren et al., Reference Souren, Lutsik, Gasparoni, Tierling, Gries, Riemenschneider and Walter2013).
Longitudinal evidence of telomere length tracking from birth to adulthood shows inverse associations of residential traffic exposure in association with telomere length at birth as well as accelerated telomere shortening in the first two decades of life (Bijnens, Zeegers, Derom et al., Reference Bijnens, Zeegers, Derom, Martens, Gielen, Hageman and Nawrot2017). Telomere length in placental tissue and in buccal cells in young adulthood, and the geocoding of residential addresses both at birth and in young adulthood, as well as residential traffic and greenness exposure were used to study this association.
Residential greenness has persistent effects on blood pressure: lower residential greenness in the early-life environment was independently associated with a higher adult blood pressure (Bijnens, Nawrot, Loos et al., Reference Bijnens, Nawrot, Loos, Gielen, Vlietinck, Derom and Zeegers2017).
Stress and Mental Health
Stress-induced increase in negative effect that regulates the individual sensitivity to small daily stress is associated with elevated cortisol levels, (sub)clinical depression and (sub)clinical psychotic symptoms, and is influenced by genes, which is indicative of gene–environment interaction. Neuroticism, as measured by the Eysenck questionnaire, may index an environmental risk for decreased daily life positive effect levels and a genetic as well as an environmental risk for increased negative effect variability (Jacobs et al., Reference Jacobs, Van Os, Derom, Thiery, Delespaul and Wichers2011). The findings are also consistent with the hypothesis that adult daily life stress sensitivity is the result of sensitization processes initiated by developmental stress exposures. Genes associated with depression may act by accelerating the process of stress-induced sensitization (Wichers et al., Reference Wichers, Geschwind, Jacobs, Kenis, Peeters, Derom and van Os2009).
Currently, more than 400 twin-pairs and their siblings in the age range of 15–35 years are taking part in a project (TwinssCan) that aims to examine genetic and environmental underpinnings of behavioral and cognitive processes (e.g., aberrant salience attribution and disturbed neurocognition) and their relation with multidimensional psychopathology using novel experimental tasks such as the white noise speech illusion task (Pries et al., Reference Pries, Guloksuz, Menne-Lothmann, Decoster, van Winkel, Collip and van Os2017), the digital social peer evaluation experiment (Menne-Lothmann et al., Reference Menne-Lothmann, Decoster, van Winkel, Collip, Rutten, Delespaul and Wichers2017) and the experience sampling method (ESM) to assess the moment-to-moment dynamic changes of mental states. The TwinssCan project aims to shed light on biological mechanisms underlying mental disorders by analyzing a wide collection of biological materials that include placenta samples, cortisol levels, and genome- and epigenomewide molecular data (Pries, Snijders, Menne-Lothmann et al., Reference Pries, Snijders, Menne-Lothmann, Decoster, van Winkel, Collip and Rutten2019). In this direction, recent ESM studies of the first wave of the TwinssCan cohort investigated the influence of childhood adversity and genetic liability on momentary mental states. The network analysis of the ESM data revealed that proxy genetic liability for psychopathology was associated with more densely connected network structure of the emotions, which include ‘cheerful’, ‘insecure’, ‘relaxed’, ‘anxious’, ‘irritated’ and ‘down’ items (Hasmi et al., Reference Hasmi, Drukker, Guloksuz, Menne-Lothmann, Decoster, van Winkel and van Os2017). A recent gene–environment interaction study using the ESM data has demonstrated that polygenic risk score for schizophrenia interacts with childhood adversities to influence negative effect, positive effect and subtle psychosis expression (Pries, Klingenberg, Menne-Lothmann et al., Reference Pries, Klingenberg, Menne-Lothmann, Decoster, van Winkel, Collip and Guloksuz2019).
Discussion
Criticism has been addressed to the classical twin studies with regard to the assumption of equal intrauterine environments of MZ twins. The most important examples of potentially unequal intrauterine environments are undoubtedly chorionicity, zygosity and the origin of the pregnancy, spontaneous or ART. Other factors, however, must also be considered: gestational age, birth sequence, birth weight, insertion of the umbilical cord, the mode of delivery and presentation of both twins.
With a sizable group of more than 1000 dichorionic MZ, 2000 monochorionic MZ and 3000 ART twin-pairs, the EFPTS can perform twin studies with fewer biases, that is, studies that are based on the crucial assumption that MZ and DZ twins, and spontaneous and ART twins have a similar intrauterine environment. Previous results have indicated that this could not be the case (Delbaere et al., Reference Delbaere, Goetgeluk, Derom, De Bacquer, De Sutter and Temmerman2007; Verstraelen et al., Reference Verstraelen, Goetgeluk, Derom, Vansteelandt, Derom, Goetghebeur and Temmerman2005). Comparison of dichorionic MZ, monochorionic MZ and DZ twins will allow better evaluation of the degree of gene–environment interactions as the prenatal environment is concerned. In contrast to all dichorionic MZ and DZ twins, monochorionic MZ twins share their chorion: most of them share their blood supply and therefore their immune system during intrauterine life. This could have long-term effects on phenotypes in later life (Craig & Saffery, Reference Craig and Saffery2015).
There is further evidence that ART can induce epigenetic variation that might be transmitted to the next generation and that subfertility itself is a risk factor for imprinting diseases (Huntriss et al., Reference Huntriss, Balen, Sinclair, Brison and Picton2018). With more than 3000 twin-pairs and 200 triplet sets being born after various techniques of ART, whether born after fertility treatment without further intervention (ovulation induction only) or after IVF/ICSI or related techniques, EFPTS is the ideal resource for the long-term follow-up study of these multiples on a population-based manner.
After 55 years, EFPTS still has the capacity to continue to collect prospectively perinatal and biological data at birth, and phenotypes in later life, and link both of them. As retrospective analysis of the placentation is still impossible to carry out and multiples born after ovulation induction only are hard to detect retrospectively, the effect of chorion type, origin of the pregnancy and the epigenetic phenomena on phenotypes in later life will remain one of the primary goals of the research for the EFPTS. As such, EFPTS shows that twin research remains a reliable tool to study not only the genetic determination of different phenotypes, but also it provides an unparalleled opportunity to study links between the (epi)genome, the intrauterine environment, air pollution exposure and (sub)clinical outcomes.
Acknowledgments
We thank all families of twins and triplets who took part in different studies. These research projects were aided by grants coming mainly from the Fund for Scientific Research — Flanders, the Dutch Foundation for Scientific Research, and the Marguerite-Marie Delacroix Foundation. The TwinssCan project is part of the European Community’s Seventh Framework Program under grant agreement No. HEALTH-F2-2009-241909 (Project EU-GEI).



