


Pulse pressure (PP) measures the change in blood pressure during a contraction of the heart. Elevated pulse pressure has been associated with cardiovascular disorders and mortality (Dart & Kingwell, Reference Dart and Kingwell2001). Several anthropometric factors have been shown to affect PP; for example, aging results in increased stiffness and loss of elasticity in the aorta and major artery leading to increased PP (Franklin et al., Reference Franklin, Gustin, Wong, Larson, Weber, Kannel and Levy1997), body-size (thus gender) affects PP through altering magnitude and timing of wave reflection (London et al., Reference London, Guerin, Pannier, Marchais and Stimpel1995; Smulyan et al., Reference Smulyan, Marchais, Pannier, Guerin, Safar and London1998), and recently age and gender effects have been shown to be cross-cultural (Skurnick et al., Reference Skurnick, Aladjem and Aviv2010).
Besides anthropometric factors, genetic epidemiology studies using family and twin data have provided verifiable evidence for a low to modest genetic contribution in pulse pressure with heritability estimates ranging from 0.13 using twins in the UK (Snieder et al., Reference Snieder, Hayward, Perks, Kelly, Kelly and Spector2000), 0.21 (Pilia et al., Reference Pilia, Chen, Scuteri, Orrú, Albai, Dei, Lai, Usala, Lai, Loi, Mameli, Vacca, Deiana, Olla, Masala, Cao, Najjar, Terracciano, Nedorezov, Sharov, Zonderman, Abecasis, Costa, Lakatta and Schlessinger2006) and 0.24 (van Rijn et al., Reference van Rijn, Schut, Aulchenko, Deinum, Sayed-Tabatabaei, Yazdanpanah, Isaacs, Axenovich, Zorkoltseva, Zillikens, Pols, Witteman, Oostra and van Duijn2007) in European pedigrees, to 0.35 and 0.54 in American Caucasian (Mitchell et al., Reference Mitchell, DeStefano, Larson, Benjamin, Chen, Vasan, Vita and Levy2005) and African families (Bochud et al., Reference Bochud, Bovet, Elston, Paccaud, Falconnet, Maillard, Shamlaye and Burnier2005). These results indicate that, similar to all other complex traits, the etiology of PP is complicated involving both genetic and environmental dissections (Turner & Boerwinkle, Reference Turner and Boerwinkle2003).
The existence of a genetic contribution has warranted efforts to elucidate genes affecting pulse pressure, especially in recent years. For example, genetic marker-based linkage studies have identified multiple loci harboring susceptibility genes for PP in different populations including Caucasians (Bielinski et al., Reference Bielinski, Lynch, Miller, Weder, Cooper, Oberman, Chen, Turner, Fornage, Province and Arnett2005; Camp et al., Reference Camp, Hopkins, Hasstedt, Coon, Malhotra, Cawthon and Hunt2003; DeStefano et al., Reference DeStefano, Larson, Mitchell, Benjamin, Vasan, Li, Corey and Levy2004; Mitchell et al., Reference Mitchell, DeStefano, Larson, Benjamin, Chen, Vasan, Vita and Levy2005), Mexicans (Atwood et al., Reference Atwood, Samollow, Hixson, Stern and MacCluer2001), American Indians (Franceschini et al., Reference Franceschini, MacCluer, Rose, Rutherford, Cole, Laston, Göring, Diego, Roman, Lee, Best, Howard, Fabsitz and North2008), and African Americans (Sherva et al., Reference Sherva, Miller, Lynch, Devereux, Rao, Oberman, Hopkins, Kitzman, Atwood and Arnett2007). The verification of susceptibility loci requires replication studies within and even across populations in order to consolidate linkage results. This is especially important given the fact that inconsistent linkage results have been reported by different studies (Turner & Boerwinkle, Reference Turner and Boerwinkle2003).
This paper reports results from the first twin-based genetic epidemiological study on pulse pressure in the Chinese population. Results from our study will be compared with those reported in the literature and the significance of our findings will be discussed.
Material & Methods
Twin Samples
All twins were taken from the most recent wave of a twin sample for a heritability study on multiple phenotypes associated with the metabolic syndrome, conducted by the Qingdao Twin Registry at Qingdao Center for Disease Control and Prevention in 2008. Twins were sampled through the local disease control network and residence registry. After informed consent was obtained, twins were invited to a clinical investigation if both co-twins were alive. Those who were pregnant, breastfeeding, had known diabetes and/or cardiovascular disease, or were taking weight-reducing medication within one month, were excluded, and incomplete twin pairs were discharged. Zygosity of like-sex twin pairs was determined by DNA testing using 16 short tandem repeat DNA markers. A total of 389 pairs of monozygotic (MZ) and 241 pairs of dizygotic (DZ) twins were sampled for heritability analysis, among which were 63 pairs of DZ twins randomly selected for genome-wide linkage analysis.
Phenotypes
Blood pressure was taken by a standard procedure using mercurial table stand model sphygmomanometer. Systolic blood pressure (SBP) was calculated as Korotkof phase I (appearance of sound), and diastolic blood pressure (DBP) as Korotkof phase V (disappearance of sound). Pulse pressure was calculated as the difference between systolic and diastolic blood pressures. BMI (kg/m2) was obtained according to measured values of height and weight, by taking the subject's weight (in kilograms) and dividing by the subject's height (in meters) squared. All measurements above three standard deviations below or over the mean were assigned as missing values. In all subsequent analyses, we use the natural log transformation of PP to ensure normal or approximate normal distribution.
DNA Samples and Genotyping
Whole blood samples were taken for extracting leukocytes for DNA purification and genotyping using standard procedures. We used the Affymetrix Genome-Wide Human SNP Array 6.0 featuring 1.8 million genetic markers among which were more than 906,600 single nucleotide polymorphisms (SNPs) enabling high resolution genome-wide analysis. Detailed information about the array can be found at the manufacturer's website (www.affymetrix.com). Genotyping was performed by the Affymetrix service provider, Shanghai Biochip, in China using purified DNA samples. Genotyping was high quality with SNP calling rate ranges from 93.4% to 99%.
Generalized Estimating Equations for Twins
It has been shown that age, sex, and BMI are important factors affecting pulse pressure (Skurnick et al., Reference Skurnick, Aladjem and Aviv2010). With our data, we first want to test the effects of these factors in Chinese subjects and then adjust PP to remove the influences by these factors in the subsequent analyses. Since our twin samples were correlated samples, we introduced the generalized estimating equations (GEE) with an exchangeable working correlation matrix (Zeger & Liang, Reference Zeger and Liang1986). GEE was applied to MZ and DZ twins separately considering differences in their genetic correlations, which could result in different working correlation matrices. GEE models were fitted using the free R package gee (http://cran.r-project.org/web/packages/gee/index.html).
Intra-Class Correlation Coefficients (ICC)
After fitting the GEEs, residuals are kept for calculating ICC to show twin correlation on pulse pressure after adjusting for age, sex, and BMI. Following the definition (McGraw & Wong, Reference McGraw and Wong1996), the statistic is calculated as 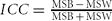 $ICC = \frac{{{\rm MSB\, -\, MSW}}}{{{\rm MSB\, +\, MSW}}}$. Here MSW is the within-pairs mean square and MSB the between-pairs mean square of the adjusted PP. ICC was estimated using the free R package psy (http://cran.r-project.org/web/packages/psy/index.html). Significance tests on statistical differences between MZ and DZ twin correlation were performed first by transforming an ICC (r) using
$ICC = \frac{{{\rm MSB\, -\, MSW}}}{{{\rm MSB\, +\, MSW}}}$. Here MSW is the within-pairs mean square and MSB the between-pairs mean square of the adjusted PP. ICC was estimated using the free R package psy (http://cran.r-project.org/web/packages/psy/index.html). Significance tests on statistical differences between MZ and DZ twin correlation were performed first by transforming an ICC (r) using 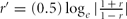 $r^\prime = (0.5)\log _e | {\frac{{1\, +\, r}}{{1\, -\, r}}} |$ and then computing the test statistic as
$r^\prime = (0.5)\log _e | {\frac{{1\, +\, r}}{{1\, -\, r}}} |$ and then computing the test statistic as 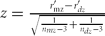 $z = \frac{{r^\prime _{{\rm mz}} - r^\prime _{dz} }}{{\sqrt {\frac{1}{{n_{mz} - 3}} + \frac{1}{{n_{dz} - 3}}} }},$ where rʹmz and rʹdz are the transformed coefficients, and n mz and n dz, the number of pairs for MZ and DZ twins. P values were obtained by referring the test statistic z to a standard normal distribution with one degree of freedom.
$z = \frac{{r^\prime _{{\rm mz}} - r^\prime _{dz} }}{{\sqrt {\frac{1}{{n_{mz} - 3}} + \frac{1}{{n_{dz} - 3}}} }},$ where rʹmz and rʹdz are the transformed coefficients, and n mz and n dz, the number of pairs for MZ and DZ twins. P values were obtained by referring the test statistic z to a standard normal distribution with one degree of freedom.
Heritability Estimation
We introduced the structural equation model to fit univariate genetic models to our twin data. The univariate model included additive genetic (A), common environmental (C), dominant genetic (D), and unique environmental (E) effects. As the effects of C and D are confounded in twin data (Rijsdijk & Sham, Reference Rijsdijk and Sham2002), two full models, i.e., the ACE and ADE models, were fitted separately to the data. Discrimination of the two models was made using Akaike's Information Criterion (AIC) with the lowest AIC for the best model. The best fitting and most parsimonious model from the nested models of either ACE or ADE was selected based on the likelihood ratio test for nested models and AIC for competing models not nested. The fitting of twin models was all performed using the free software package Mx (Neale et al., Reference Neale, Boker, Xie and Maes2003).
Genome-Wide Linkage Analysis
We applied the variance components method for non-parametric linkage analysis (Marlow, Reference Marlow2002) to our genome-wide SNP marker data on DZ twins. Data were analyzed with the free linkage software package, Merlin (Abecasis et al., Reference Abecasis, Cherny, Cookson and Cardon2002). Before linkage analysis, our SNP genotype data were first processed for genotyping errors using the error-detection procedure in Merlin (–error). The detected unlikely genotypes were removed from the data using the Merlin command pedwipe. The variance components (Amos, Reference Amos1994) (–vc) procedure was used to test for linkage for pulse pressure with age, sex, and BMI incorporated as covariates.
Although our high density SNP map increases linkage information content which is important in non-parametric linkage analysis (Evans & Cardon, Reference Evans and Cardon2004), on the other hand, it could also result in linkage disequilibrium (LD) among adjacent SNPs. When parental phase information is missing, LD can lead to overestimation of the number of alleles shared by identical-by-descent (IBD) and thus inflate the lod score estimates (Cho & Dupuis, Reference Cho and Dupuis2009). To accommodate LD, Merlin organizes correlated SNPs into clusters and assumes linkage equilibrium between the clusters and no recombination within clusters. This is done using the Merlin procedure –cluster and defining a threshold for pairwise marker-marker correlation r 2. In our analysis, we introduce a stringent threshold of 0.1. According to a simulation study by Cho and Dupuis (Cho & Dupuis, Reference Cho and Dupuis2009), such a threshold can efficiently eliminate the lod score inflation due to LD.
Genome-wide significance of the identified linkage peaks were assessed by applying the threshold criterion provided by Lander and Kruglyak (Lander & Kruglyak, Reference Lander and Kruglyak1995) with a score of 2.2 for suggestive and 3.6 for genome-wide significant linkages.
Results
Basic Statistics
The basic statistics for all subjects are shown in Table 1 for MZ and DZ twins separately. The mean age for all twins was approximately 37 years with a mean BMI around 24 kg/m2, mean SBP around 119 mmHg, mean DBP around 81 mmHg, and mean PP around 39 mmHg. In both GEE models for MZ and DZ twins, age showed a highly significant (p < .002) positive correlation with PP indicating a slow but steady increase of PP with increasing age, when sex and BMI are balanced. Sex has very highly significant negative correlations consistent in both MZ and DZ twins meaning that, for given age and BMI, females tend to have lower PP than males. The effect for BMI is also highly significant with positive correlation in both MZ and DZ twins suggesting that when age and sex are fixed, PP increases with increasing BMI. Figure 1 plots the residuals (i. e., PP adjusted for age, sex, and BMI) from the GEE models for MZ and DZ twins. As can be seen in the figure, MZ twins exhibit an obvious higher correlation compared to DZ twins. The intra-pair correlation for the adjusted PP is shown in the bottom of Table 1 with ICCs of 0.42 for MZ (p = 5e-07) and 0.19 for DZ (p = 3.06e-03) twins, both are statistically highly significant. ICCs in MZ and DZ twins are significantly different with a p value of .002 (z score = 3.10, df = 1).
TABLE 1 Descriptive, Regression, and Correlation Analyses

*ICCs in MZ and DZ twins are statistically significantly different with a p value of 1.90e-03 (Z = 3.10).

FIGURE 1 Scatter plot for the residuals of log PP from GEE models fitted to MZ (left) and DZ (right) twins. MZ twins are more closely correlated than the DZ twin suggesting the existence of genetic contributions to PP.
Heritability Estimates
As the ICC for MZ twins is more than double the ICC for DZ twins, both ACE and ADE models were fitted to PP with age, sex, and BMI incorporated as covariates. Table 2 shows the results for the two models together with their corresponding nested models. Between the two competing full models, the ADE model slightly outperformed the ACE model according to their AICs (AIC for ADE model slightly more negative than AIC for ACE model). However, based on the likelihood ratio test and AICs, the nested parsimonious AE model for both ACE and ADE models was suggested as the best model with an estimate for additive genetic effect that accounts for 45% of the total variance in PP (95% CI 0.37–0.52). Performances for all other nested models were very significantly worse than that of their full models.
TABLE 2 Estimates (95% Confidence Interval) and Model Performances for ACE, ADE, and Their Nested Models

Genome-Wide Linkage Analysis
With age, sex, and BMI incorporated as covariates, our whole genome linkage scan (Table 3) identified three significant linkage peaks at chromosome 11 (lod score of 4.06, linkage region from 26–33 cM), chromosome 12 (lod score of 3.97, from 93–104 cM) and chromosome 18 (lod score of 4.01, from 68–72 cM) (Figures 2 and 3), together with multiple suggestive linkage peaks (Figure 2) with lod scores over 3 on chromosome 1 (3.05 from 81–100 cM, 3.54 from 170–242 cM), chromosome 3 (3.6 from 141–170 cM), chromosome 4 (3.08 from 72–128 cM), chromosome 6 (3.19 from 129–163cM), chromosome 7 (3.24 from 55–64 cM, 3.41 from 68–88 cM, 3.18 from 95–116), chromosome 9 (3.49 from 154–167 cM), chromosome 17 (3.32 from 61–103 cM), and chromosome 22 (3.04 from 1–31 cM).
TABLE 3 Significant and Suggestive Linkage Peaks for Pulse Pressure


FIGURE 2 Linkage results for genome-wide scan by chromosomes. Multiple suggestive linkage peaks were found on different chromosomes except chromosomes 2, 13, 14, 19, and 21.

FIGURE 3 The significant linkage peaks identified on chromosomes 11, 12, and 18 with lod score estimates of 4.06, 3.97, and 4.01.
Discussion
We have conducted the first extensive genetic study on pulse pressure in the Chinese population that covered both heritability estimation and gene mapping using a linkage approach. The significant heritability estimate confirms the genetic dissection on pulse pressure in the Chinese population, which is supported by our subsequent linkage analyses that identified multiple genomic regions harboring susceptibility loci for pulse pressure. Although each of the two approaches focused on different aspects of the genetic architecture of PP, overall, our significant results emphasize the genetic control over hemodynamic conditions.
By applying the GEE model to MZ and DZ twins separately, we were able to examine the effects of age, sex, and BMI on pulse pressure in Chinese twins. Highly significant effects of age, sex, and BMI were found in both MZ and DZ twins. Zhao et al. (Reference Zhao, Stamler, Yan, Zhou, Wu, Liu, Daviglus, Dennis, Elliott, Ueshima, Yang, Zhu and Guo2004) reported significant effects of dietary factors including salt intake on blood pressure differences between Chinese in the north (higher salt intake) and the south (lower salt intake). As a limitation of the study, salt intake was not measured and thus cannot be adjusted although individual adjustment on a population factor can only have limited effect within the same population. On the other hand, our models for gene mapping are based on intra-pair differences that match the effects of shared environmental factors (for example, family diet habits). It is encouraging to see that our results based on Chinese data are consistent with that from studies on other ethnic populations and give new support to the conclusion that the effects of these factors are cross-sectional (Skurnick et al., Reference Skurnick, Aladjem and Aviv2010).
Although low to modest heritabilities have been estimated in many different ethnic populations, to our knowledge, our twin-based study is the first to focus on PP in the Chinese population. Our heritability estimate of 0.45 is in agreement with recent studies reporting modest genetic effects in American Caucasians (DeStefano et al., Reference DeStefano, Larson, Mitchell, Benjamin, Vasan, Li, Corey and Levy2004) and Africans (Bochud et al., Reference Bochud, Bovet, Elston, Paccaud, Falconnet, Maillard, Shamlaye and Burnier2005). Comparison of model performances suggested that the parsimonious AE model as the best model indicating the existence of additive genetic regulation on PP.
Three linkage peaks that reside separately on chromosomes 11, 12, and 18 have been found to show genome-wide significance (lod score > 3.6) with their lod score estimates of around 4. It is surprising that among the three significant linkage peaks, two have been confirmed to show linkage in other populations. In a multipoint linkage scan performed on large and extended Utah pedigrees, Camp et al. (Reference Camp, Hopkins, Hasstedt, Coon, Malhotra, Cawthon and Hunt2003) reported linkage evidence on chromosome 12q at 109 cM which is covered by our linkage region on chromosome 12 in Figure 3. In another study based on American families, Bielinski et al. (Reference Bielinski, Lynch, Miller, Weder, Cooper, Oberman, Chen, Turner, Fornage, Province and Arnett2005) detected a significant linkage for PP on chromosome 18 at 71 cM, the most significant linkage in their study. It is interesting that this is exactly the location for our linkage peak on chromosome 18 (at 70.75 cM, Figure 3). Considering that there have been only a limited number of whole genome linkage scan on PP conducted in the literature, it is very unlikely that these results are coincidence.
Multiple suggestive linkages were detected by our linkage scan (Table 3). We assume that their trustworthiness increases with their lod score magnitudes. This seems to be reasonable considering the fact that four suggestive peaks with lod scores over 3 overlap with reported linkages in previous studies. The highest suggestive linkage peak on chromosome 1 lies in the same region where Sherva et al. (Reference Sherva, Miller, Lynch, Devereux, Rao, Oberman, Hopkins, Kitzman, Atwood and Arnett2007) reported a suggestive linkage at 215 cM. This region resides the G protein-coupled receptor gene, which is involved in the signaling cascades that regulate blood pressure during changes in cardiac output (Sherva et al., Reference Sherva, Miller, Lynch, Devereux, Rao, Oberman, Hopkins, Kitzman, Atwood and Arnett2007). The highest suggestive linkage peak on chromosome 7 at 74.37 cM coincides with a linkage peak at 75cM reported by Bielinski et al. (Reference Bielinski, Lynch, Miller, Weder, Cooper, Oberman, Chen, Turner, Fornage, Province and Arnett2005), and is in the vicinity of a peak at 71 cM in the same region reported by DeStefano et al. (Reference DeStefano, Larson, Mitchell, Benjamin, Vasan, Li, Corey and Levy2004). The peak regions on chromosomes 17 and 22 cover linkage peaks for pulse pressure reported by Bielinski et al. (Reference Bielinski, Lynch, Miller, Weder, Cooper, Oberman, Chen, Turner, Fornage, Province and Arnett2005) (at 89 cM on chromosome 17 and 11 cM on chromosome 22). In addition, their study also detected linkages on chromosomes 1 (106 cM), 3 (172 cM), and 7 (75 cM) that are located in the vicinity of our suggestive regions. These results emphasize the importance of our suggestive linkages.
The performances of SNP and traditional microsatellite markers in linkage analysis have been compared by Schaid et al. (Reference Schaid, Guenther, Christensen, Hebbring, Rosenow, Hilker, McDonnell, Cunningham, Slager, Blute and Thibodeau2004) and Evans and Cardon (Reference Evans and Cardon2004) who consistently reported a higher information content from the dense map of SNP markers than the from the microsatellite markers spaced at 1 marker per ~10 cM. Moreover, it was reported that linkage mapping using dense SNP markers identified more linkage peaks with more narrow widths than did traditional markers (Schaid et al., Reference Schaid, Guenther, Christensen, Hebbring, Rosenow, Hilker, McDonnell, Cunningham, Slager, Blute and Thibodeau2004). This conclusion coincides with our multiple suggestive scores with many of them overlapping with linkage peaks detected by published large studies.
Our identified significant and suggestive linkage peaks especially those overlapping closely with previous studies encourage further investigations with aim at identifying genetic variations that affect pulse pressure using more focused linkage and association approaches on large samples. Meanwhile, interpretation of the multiple suggestive linkages should be made with caution since the validity of these loci requires additional replication studies to be conducted particularly in the Chinese and Asian populations.
List of Abbreviations
- AIC:
Akaike information criterion
- BMI:
body mass index
- DBP:
diastolic blood pressure
- DZ:
dizygotic
- GEE:
generalized estimating equation
- IBD:
identical-by-descent
- ICC:
intra-class correlation coefficient
- LD:
linkage disequilibrium
- MZ:
monozygotic
- PP:
pulse pressure
- SBP:
systolic blood pressure
- SNP:
single nucleotide polymorphism
Acknowledgments
This project was funded by the European Foundation for the Study of Diabetes, 2007 Research Project ‘A genome-wide linkage analysis on metabolic/intermediate phenotypes predisposing to type II diabetes in the Chinese population’ and by the National Natural Science Foundation of China (grant #30872170). The authors are grateful to Dr. Gu Zhu at the Genetic Epidemiology Unit, Queensland Institute of Medical Research, Australia, for excellent technical help.






