



1. Introduction
The Cambrian Stage 4 Guanshan Biota is one of the famous Burgess-Shale-type Lagerstätten found in Yunnan, South China, that continues to yield exquisitely preserved fossils with soft tissues and organs, and greatly attracts the interests of palaeontologists and evolutionary biologists (Yang et al. Reference Yang, Hou, Cong, Dong, Zhang and Luo2010; Hu et al. Reference Hu, Zhu, Luo, Steiner, Zhao, Li, Liu and Zhang2013, Reference Hu, Zhu, Zhao and Steiner2017; Hopkins et al. Reference Hopkins, Chen, Hu and Zhang2017; Chen et al. Reference Chen, Zhang, Betts, Zhang and Liu2019 a, Reference Chen, Legg, Zhai, Liu and Hou2020). While the type localities (i.e. Gaoloufang and Gangtoucun) have been irrevocably lost as a result of urban development, new sites yielding soft-bodied fossils have been discovered in Yunnan, South China, such as Yiliang County (Jiao et al. Reference Jiao, Yang and Zhang2016), Malong County (Chen et al. Reference Chen, Zhang, Betts, Zhang and Liu2019 a), Wuding County (Liu et al. Reference Liu, Han, Li, Wu, Peng, Qi, Yang and Li2016), Baimei village (south of Kunming, Liu et al. Reference Liu, Han, Li, Wu, Peng, Qi, Yang and Li2016) and Huize County (Liu et al. Reference Liu, Ou, Han, Zhang, He, Yao, Fu and Shu2012). The fossils of non-trilobite artiopods are extremely rare at all the Guanshang Biota localities, and the most recent discovery of a new artiopod species was made 12 years ago (Luo et al. Reference Luo, Li, Hu, Fu, Hou, Liu, Chen, Li, Pang and Liu2008; Hu et al. Reference Hu, Zhu, Luo, Steiner, Zhao, Li, Liu and Zhang2013). In this contribution, we describe a small-sized but well-preserved non-trilobite artiopod from the Cambrian Guanshan Biota, and investigate its phylogenetic relationships among early euarthropods with the phylogeny programme TNT. Parsimony analyses based on morphological characters support a position for the new taxon among the non-trilobite Artiopoda.
2. Materials and methods
The single articulated and completely preserved specimen (RCP 001) was collected from Yiliang County, and is housed in the Research Center of Paleobiology, Yuxi Normal University (RCP).
The specimen was photographed with a LEICA DFC 550 digital camera mounted to a Stereoscope LEICA M205 C, and a LEICA DFC7000 T monochrome digital camera attached to a LEICA M205 FA fluorescence stereomicroscope. This specimen was also scanned with a Zeiss Xradia 520 Versa X-ray Microscope, but the result did not provide more useful information.
The character matrix (online Supplementary Material, available at http://journals.cambridge.org/geo), including 65 taxa and 89 characters, is an updated version of the dataset used by Chen et al. (Reference Chen, Ortega-Hernández, Wolfe, Zhai, Hou, Chen, Mai and Liu2019 b; see also Ortega-Hernández et al. Reference Ortega-Hernández, Legg and Braddy2013; Lerosey-Aubril et al. Reference Lerosey-Aubril, Zhu and Ortega-Hernández2017; Du et al. Reference Du, Ortega-Hernández, Yang and Zhang2019) (the tailspine of Retifacies is shorter than half of its trunk, and the lateral notches of the head shield are present on Acanthomeridion; K.S. Du, personal observation). The parsimony analyses were run in TNT version 1.5 under New Technology Search, using Driven Search with Sectorial Search, Ratchet, Drift and Tree fusing options activated with standard settings (Goloboff & Catalano, Reference Goloboff and Catalano2016; Du et al. Reference Du, Ortega-Hernández, Yang and Zhang2019).
3. Systematic palaeontology
Euarthropoda Lankester, Reference Lankester1904
Artiopoda Hou & Bergström, Reference Hou and Bergström1997
Bailongia gen. nov. (Figs 1–2)

Fig. 1. Completely articulated specimen of Bailongia longicaudata gen. et sp. nov. from the Cambrian Stage 4 Guanshan Biota. (a) RCP 0001b preserving the semi-elliptical cephalon with right genal spine, small antennae, possible scales on the right antenna (arrows), cephalic limbs, eyes, nine tapering tergites, lamellae of exopod and the long tailspine. (b) RCP 0001a preserved the semi-elliptical head shield, cephalic limbs, eyes, nine tapering tergites, a trunk endopod and the long tailspine. (c) Interpretive drawing of RCP 0001a. (d, e) Close-up of cephalon, genal spine, antennae, possible scales on the right antenna (arrows), post-antennal cephalic limbs and eyes. (f) Details of the eyes, the only exposed trunk endopod. (g) Close-up of the lamellae of trunk exopod. (h) Details of the tergites two to six. (i) Close-up of tergites six to nine, photographed with fluorescent microscopy. Abbreviations: ant – antenna; en – endopod; ey – eye; gs – genal spine; hs – head shield; hy – hypostome; lam – lamellae; lim – limb; Tn – tergite n; ts – tailspine.
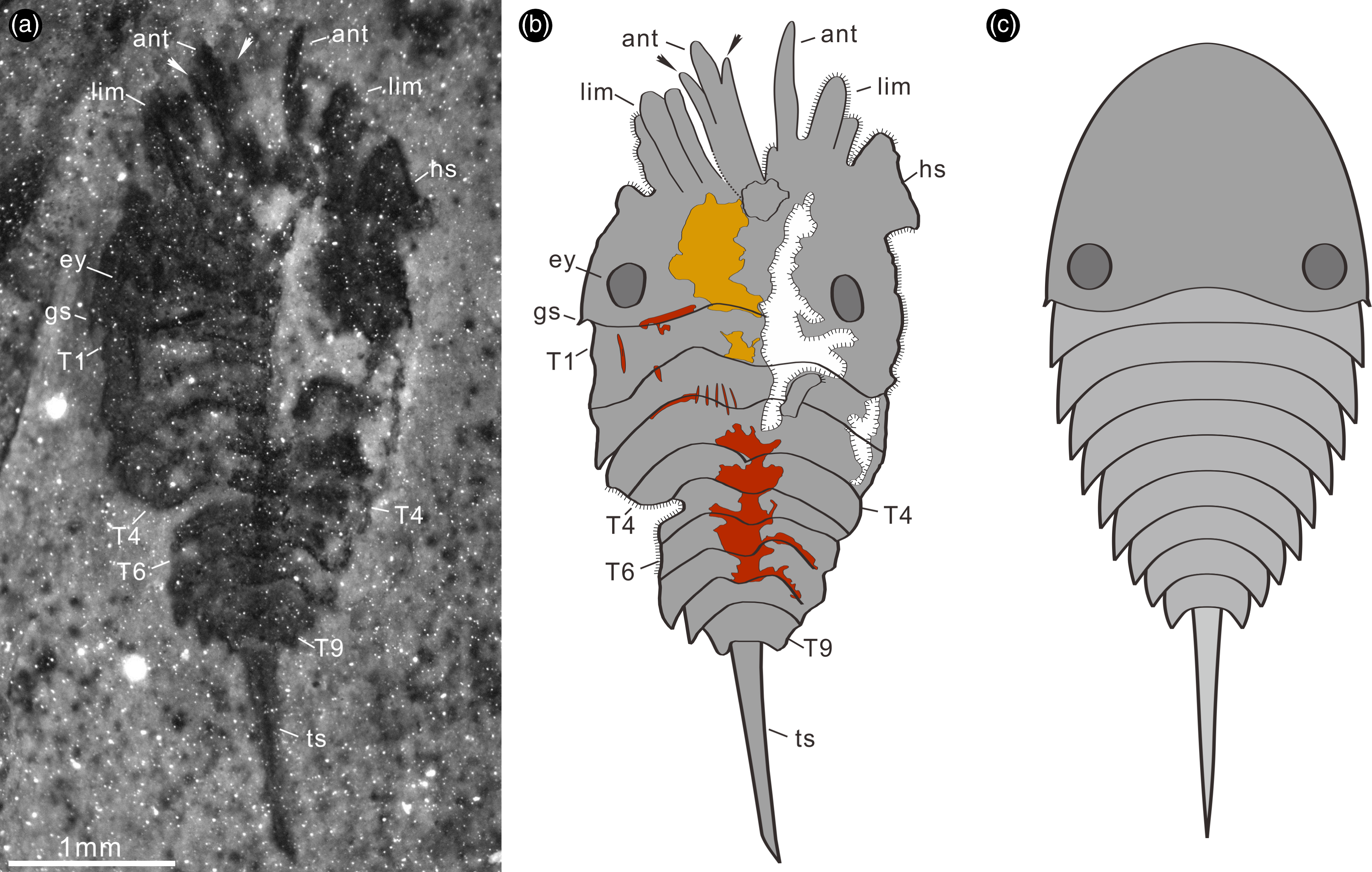
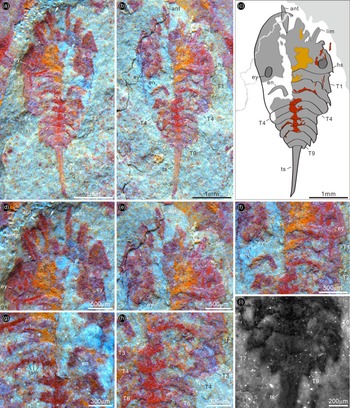
Fig. 2. Bailongia longicaudata gen. et sp. nov. from the Guanshan Biota. (a) Fluorescent photograph of RCP 0001b. (b) Interpretive drawing of RCP 0001b showing the cephalon with genal spine, small antennae, possible scales on the right antenna (arrows), cephalic limbs, eyes, nine tapering tergites and the long tailspine. (c) Morphological reconstruction. Abbreviations as for Figure 1.
Diagnosis. Small arthropod, c. 5 mm in length, with a long and wide semi-oblong head shield possessing a pair of lateral posterior eyes, an elongated thorax consisting of nine imbricated tapering tergites and a long slender tailspine. The cephalon, thorax and tailspine are approximately the same longitudinal length.
Etymology. ‘Bailong’ is the name of a geographical area near the fossil site; longicaudata refers to its long tailspine.
Bailongia longicaudata sp. nov. (Figs 1–2)
Type material, locality and stratigraphy. Holotype, RCP 0001, a complete specimen (Fig. 1). Bailongia longicaudata gen. et sp. nov. was discovered in the outcrop near Lihuazhuang village (see Jiao et al. Reference Jiao, Pates, Lerosey-Aubril, Ortega-Hernández, Yang, Lan and Zhang2021, Fig. 1) in Yiliang, Kunming, Yunnan; Cambrian Stage 4 Wulongqing Formation, Palaeolenus Biozone.
Diagnosis. As for genus by monotypy.
Description. The only available specimen is complete and fully articulated. It reaches a maximum length (sagittal) of 5 mm including antenna and tailspine, and a maximum width (transversal) of 2.1 mm (Fig. 1a–c). The dorsal exoskeleton consists of a long and wide semi-elliptical cephalon with a pair of lateral posterior eyes (Fig. 1a–e), a tapering thorax possessing nine imbricated tergites (Fig. 1a–c, f, h, i) and a long slender tailspine (Fig. 1a–c). The enormous head shield has a posterior bay-like margin that curves forwards (Fig. 1a–f), and is about one-third of the total body length (sag.) (Fig. 1a–c). The cephalon bears a pair of genal spines, a pair of small antennae, at least three pairs of post-antennal limbs and a pair of lateral posterior elliptical eyes (Fig. 1a–f); the left antenna seems to have paired antennal scales (Figs 1a, d; 2a, b, arrows); the cephalic appendages are recognized, but their segments are unclear (Figs 1a–e; 2a) even with the help of the fluorescence stereomicroscope. Following the cephalon, there are nine imbricated tapering tergites lacking pleural spines, which maintain a subequal length (sag.) and possess extended tergopleurae (Figs 1a–c, f, h, i; 2a, b); the left pediform endopod under tergite two is preserved although its segments are unclear (Figs 1a–c, f, g; 2a); the lamellae of exopod are well exposed (Fig. 1a, g). The long preserved extent of the tailspine accounts for approximately one-third of the total body length (sag.) (Figs 1a–c; 2a–c).
4. Discussion and conclusions
The first antenniform limbs, exopod with lamellae, and homonomous dorsal exoskeleton with expanded tergopleurae of Bailongia all support its assignment to Artiopoda Hou & Bergström, Reference Hou and Bergström1997, but its detailed morphology does not readily fit within any of the major artiopod clades. The features identified as diagnostic of the proposed non-trilobite artiopod clades (namely Protosutura, Nektaspida, Conciliterga, Petalopleura, Xenopoda, Cheloniellida or Aglaspidida) do not readily fit the new taxon for its long and wide semi-elliptical head shield with lateral posterior eyes, nine tapering imbricated tergites, a long slender tailspine that is longer than half of its trunk, and the small exoskeleton.
Bailongia bears a resemblance to Retifacies from the Cambrian Stage 3 Chengjiang Biota in the presence of paired genal spines on its head shield and a long tailspine (Hou & Bergström, Reference Hou and Bergström1997). The new taxon differs from Retifacies by bearing a huge semi-elliptical cephalon, lateral eyes, nine tapering tergites and a long tailspine that is longer than half of its trunk. There is a difficulty to determine whether the eyes of Bailongia are sessile or stalked, or accommodated in dorsal exoskeletal bulges based on this single specimen. By considering those expressed in Sinoburius, Xandarella spectaculum and Phytophilaspis (Ivantsov, Reference Ivantsov1999; Chen et al. Reference Chen, Ortega-Hernández, Wolfe, Zhai, Hou, Chen, Mai and Liu2019 b), we suggest that the eyes of Bailongia may be stalked and accommodated in bulges. The possible two scales on the right antenna of Bailongia resemble those on the antennae of Sinoburius, although there is a single scale on each antenna of the latter (Chen et al. Reference Chen, Ortega-Hernández, Wolfe, Zhai, Hou, Chen, Mai and Liu2019 b). If this is correct, the antennae of Bailongia are uniramous as same as those of Sinoburius and Misszhouia longicaudata (Zhang et al. Reference Zhang, Shu and Erwin2007; Chen et al. Reference Chen, Ortega-Hernández, Wolfe, Zhai, Hou, Chen, Mai and Liu2019 b). Bailongia resembles Xandarella spectaculum (Hou & Bergström, Reference Hou and Bergström1997) from the Chengjiang Biota in the presence of a long and wide semi-elliptical head, but differs from it in the posteriorly located eyes, nine tapering tergites without the reduction of the first tergite, and a long styliform tailspine.
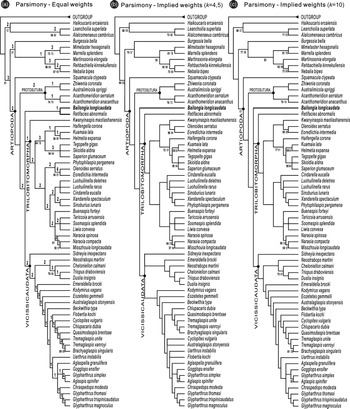
Parsimony under equal weights (EW) and implied weights (IW) are congruent in the phylogenetic position of Bailongia within Artiopoda, recovering Squamacula as the earliest diverging representative of Artiopoda, and resolving artiopods consisting of three main clades, i.e. Protosutura + Trilobitomorpha + Vicissicaudata (Fig. 3), which are similar to previous studies (e.g. Du et al. Reference Du, Ortega-Hernández, Yang and Zhang2019; Chen et al. Reference Chen, Ortega-Hernández, Wolfe, Zhai, Hou, Chen, Mai and Liu2019 b). IW-parsimony analyses (k = 4, 5, 10) support Bailongia at the position that has a diverse assemblage composed of Trilobitomorpha and Vicissicaudata as its sister taxon (Fig. 3b, c). Although the head shield with genal spines (Char 36) supports Bailongia and Retifacies as a sister-group under EW-parsimony (Fig. 3a), we regard this result as suspicious given that it is inconsistent with the outcomes under IW-parsimony (Fig. 3b, c), the two genera share few characters, and genal spines as an extremely weak character are highly convergent.

Fig. 3. Results of parsimony analyses. All trees depict a strict consensus. (a) Equal weight parsimony (two most-parsimonious trees (MPTs), 258 steps, consistency index (CI) = 0.415, retention index (RI) = 0.742). (b) Implied weight parsimony, k = 4, 5 (two MPTs, CI = 0.408, RI = 0.735). (c) Implied weight parsimony, k = 10 (one MPT, CI = 0.413, RI = 0.741). Bremer support values are shown in bold above each node in (a). Jack-knife resampling values > 50% are shown in bold and italics under each node. Bootstrap resampling values > 50% are shown under each node.
In conclusion, while Bailongia can be placed confidently in Artiopoda, its detailed morphology cannot readily be accommodated within any of the major artiopod clades. B. longicaudata provides new insights into the community structure of the Guanshan Biota in Yiliang County, and further helps to advance our understanding of the diversity and evolution of the non-trilobite artiopods. The autoecology of this new taxon will require further investigation. We hope that further field work in Yiliang County will find more specimens of non-trilobite artiopods.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/S0016756821001254
Acknowledgements
We thank Gregory D. Edgecombe (The Natural History Museum) and Peter Van Roy (Ghent University) for their constructive reviews. Special acknowledgements are given to Tian Lan, Jin-Bo Hou and Jia-Lin Du (Yunnan University) for their help with fieldwork. We sincerely thank Javier Ortega-Hernández (Harvard University) for his valuable suggestions. This work was supported by the National Natural Science Foundation of China (X.G. Zhang, grant no. 41730318; J. Yang, grant no. 42162002) and an Innovative Research Fund of Yuxi Normal University (K.S. Du, grant no. 202111390020).
Conflict of interest
None.



 Open access
Open access