



Introduction
Cystic echinococcosis (CE) is a zoonotic disease caused by metacestodes of the cestode Echinococcus granulosus sensu lato (s. l.). The World Health Organization (WHO) has listed echinococcosis among 20 neglected diseases targeted for control or elimination by 2050 (WHO, 2012). Although CE has a worldwide distribution, it has major public health and economic impact in areas of extensive livestock keeping (Deplazes et al., Reference Deplazes, Rinaldi, Alvarez Rojas, Torgerson, Harandi, Romig, Antolova, Schurer, Lahmar, Cringoli, Magambo, Thompson and Jenkins2017). The lifecycles of the various species within E. granulosus s. l. involve mainly canids as definitive hosts, and a range of herbivorous or omnivorous intermediate hosts in which the metacestode develops (Thompson, Reference Thompson2017).
The E. granulosus s. l. complex includes at least 5 cryptic species and some distinct genotypes (G), namely E. granulosus sensu stricto (s. s.) (G1, G3, GOmo), E. equinus (G4), E. ortleppi (G5), E. canadensis (G6–8, G10) and E. felidis (Nakao et al., Reference Nakao, Lavikainen, Yanagida and Ito2013a, Reference Nakao, Yanagida, Konyaev, Lavikainen, Odnokurtsev, Zaikov and Ito2013b; Wassermann et al., Reference Wassermann, Woldeyes, Gerbi, Ebi, Zeyhle, Mackenstedt, Petros, Tilahun, Kern and Romig2016; Vuitton et al., Reference Vuitton, McManus, Rogan, Romig, Gottstein, Naidich, Tuxun, Wen and Menezes da Silva2020). Epidemiological studies in eastern Africa have reported all these taxa except for E. canadensis G8 and G10 (which are wildlife parasites from the northern Holarctic) and E. equinus. The latter mainly infects intermediate hosts of the horse family (Equidae), is rarely zoonotic and occurs around the globe mainly in domestic lifecycles involving dogs and horses or donkeys (Romig et al., Reference Romig, Deplazes, Jenkins, Giraudoux, Massolo, Craig, Wassermann, Takahashi and de la Rue2017).
In Africa, E. equinus is known to infect donkeys north of the Sahara at considerable frequency (Azlaf and Dakkak, Reference Azlaf and Dakkak2006; Haridy et al., Reference Haridy, Gawad, Ibrahim, Hassan, El-Sherbi, El Shazly and Morsy2008; Taha, Reference Taha2012; Aboelhadid et al., Reference Aboelhadid, El-Dakhly, Yanai, Fukushi and Hassanin2013; Boufana et al., Reference Boufana, Lahmar, Rebai, Ben Safta, Jebabli, Ammar, Kachti, Aouadi and Craig2014; Lahmar et al., Reference Lahmar, Boufana, Jebabli, Craig, Ayari, Basti, Dhibi and Torgerson2014; Mahdy et al., Reference Mahdy, Abdel-Maogood, Abdel-Wahab and El-Bahy2014a, Reference Mahdy, Maogood, Wahab and El-Bahy2014b; Barghash et al., Reference Barghash, El Sayed, El-Alfy, Abou-Elnour, El-Kattan and Sadek2017; Desouky et al., Reference Desouky, Helmy, Sorour and Amer2017), and in southern Africa, it has been reported from horses, zebras and rhinos (Kumaratilake et al., Reference Kumaratilake, Thompson and Eckert1986; Wassermann et al., Reference Wassermann, Aschenborn, Aschenborn, Mackenstedt and Romig2015; Romig et al., Reference Romig, Deplazes, Jenkins, Giraudoux, Massolo, Craig, Wassermann, Takahashi and de la Rue2017; Zaffarano et al., Reference Zaffarano, de Klerk-Lorist, Junker, Mitchell, Bhoora, Poglayen and Govender2021). However, there are no records from the west, central and east of the continent. At least for eastern Africa, the absence of E. equinus can be explained by sampling bias. While numerous studies in the last decade contributed to our understanding of CE epidemiology, mainly in Kenya, by screening livestock, wildlife and humans (Kagendo et al., Reference Kagendo, Magambo, Agola, Njenga, Zeyhle, Mulinge, Gitonga, Mbae, Muchiri, Wassermann, Kern and Romig2014; Mbaya et al., Reference Mbaya, Magambo, Njenga, Zeyhle, Mbae, Mulinge, Wassermann, Kern and Romig2014; Addy et al., Reference Addy, Wassermann, Banda, Mbaya, Aschenborn, Aschenborn, Koskei, Umhang, De La Rue, Elmahdi, Mackenstedt, Kern and Romig2017a; Romig et al., Reference Romig, Deplazes, Jenkins, Giraudoux, Massolo, Craig, Wassermann, Takahashi and de la Rue2017; Mulinge et al., Reference Mulinge, Magambo, Odongo, Njenga, Zeyhle, Mbae, Kagendo, Addy, Ebi, Wassermann, Kern and Romig2018; Odongo et al., Reference Odongo, Tiampati, Mulinge, Mbae, Bishop, Zeyhle, Magambo, Wasserman, Kern and Romig2018; Kere et al., Reference Kere, Joseph, Jessika and Maina2019; Nungari et al., Reference Nungari, Mbae, Gikunju, Mulinge, Kaburu, Zeyhle and Magambo2019; Omondi et al., Reference Omondi, Gitau, Gathura, Mulinge, Zeyhle, Kimeli and Bett2020), none of these surveys involved donkeys, horses or zebras. This was mainly due to the absence of slaughter facilities for donkeys, as they were not slaughtered for human consumption even though the government of Kenya had gazetted donkeys as food animals in 1999 (Legal Act Notice No146, 1999). Only recently, several export slaughterhouses for donkeys were licensed to operate in Kenya, which provided a research opportunity to close the gap of knowledge on the presence, frequency and causative species of CE in donkeys and to determine their role in the transmission cycles.
Materials and methods
Study sites
The study was done in 3 private-owned slaughterhouses, 1 in Turkana County (Lodwar: Silzha Ltd.) and 2 in central Kenya (Naivasha: Star Brilliant Ltd. and Mogotio: Goldox Kenya Ltd.) (Fig. 1). The origin of the donkeys was identified to county level in Kenya; donkeys from Uganda were from the Karamoja area, donkeys from Tanzania were from the Maasai area in northern Tanzania, and donkeys from Ethiopia entering Kenya through the border town of Moyale originated from the adjacent Borena zone of southern Ethiopia (Fig. 1).

Fig. 1. A map of eastern Africa showing the location of the 3 slaughterhouse, the origin of the donkeys in Kenya and the neighbouring countries, and the 4 sub-regions listed in Table 2.
Collection of CE cysts and microscopic examination
The study was done at 2 intervals; the first between January and September 2017 which involved all the 3 slaughterhouses (3197 donkeys). The second phase was carried out only in Naivasha slaughterhouse between February and August 2019 (2764 donkeys). Carcasses were examined for cysts by palpation and subsequent incision of the major organs. The cysts/lesions were excised in toto, cleaned and stored individually in 70% ethanol. The cysts were examined microscopically for the presence of protoscoleces, and with sterile cysts (not containing protoscoleces), the integrity of the germinal layer was visually assessed to discriminate between viable and degenerated cysts.
DNA extraction
Cyst material or a single protoscolex were lysed in 0.02 M NaOH at 99°C for 10 min (Nakao et al., Reference Nakao, Sako and Ito2003). The lysate was used as template immediately following polymerase chain reaction (PCR). Cyst samples that failed to yield a PCR product following the lysis procedure were subjected to DNA extraction using DNeasy Blood and Tissue kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions.
Polymerase chain reaction-restriction fragment-length polymorphism, PCR purification and sequencing
Two nested PCR assays based on NADH dehydrogenase subunit 1 gene (nad1) were used for genotyping of cysts depending on their conditions. The first PCR targeted the entire nad1 gene (894 bp) and was carried out according to Hüttner et al. (Reference Hüttner, Nakao, Wassermann, Siefert, Boomker, Dinkel, Sako, Mackenstedt, Romig and Ito2008). The second PCR amplified part of the nad1 gene (550–552 bp) as described by Mulinge et al. (Reference Mulinge, Magambo, Odongo, Njenga, Zeyhle, Mbae, Kagendo, Addy, Ebi, Wassermann, Kern and Romig2018) and was carried out on samples that failed to yield an amplicon with the first PCR. In both PCR assays, the reaction mixture contained 2 μL of the DNA, 1 × DreamTaq Green Buffer [20 mm Tris-HCl (pH 8.0), 1 mm Dithiothreitol (DTT), 0.1 mm Ethylenediamine tetraacetic acid (EDTA), 100 mm KCl, 0.5% (v/v) Nonidet P40, 0.5% (v/v) Tween 20] (Thermo Scientific, Waltham, MA, USA), 0.2 mm deoxynucleotide triphosphate (New England Biolabs, Ipswich, MA, USA), 0.25 μ m of forward and reverse primers each, 2 mm MgCl2 and 0.625 units of DreamTaq Green DNA Polymerase (Thermo Scientific) in 25 μL final volume. The PCR cycling conditions were 5 min for initial denaturation at 94°C, 40 cycles of 94°C for 30 s, 55°C for 30 s and 72°C for 60 s, and a final extension at 72°C for 5 min (Hüttner et al., Reference Hüttner, Nakao, Wassermann, Siefert, Boomker, Dinkel, Sako, Mackenstedt, Romig and Ito2008).
The nad1 PCR products were digested using the restriction enzyme, HphI (New England Biolabs), either the entire or partial nad1 gene (Hüttner et al., Reference Hüttner, Siefert, Mackenstedt and Romig2009; Mulinge et al., Reference Mulinge, Magambo, Odongo, Njenga, Zeyhle, Mbae, Kagendo, Addy, Ebi, Wassermann, Kern and Romig2018). The total reaction mixture was 20 μL including 7.5 μL nuclease-free water, 2.0 μL of 10 × CutSmart buffer, 0.5 μL HphI (5 units) and 10 μL PCR product. The restriction digests were incubated overnight at 37°C and separated on 3% agarose gel alongside positive controls for E. granulosus s. s., E. equinus, E. ortleppi, E. canadensis (G6/7) and E. felidis (Hüttner et al., Reference Hüttner, Siefert, Mackenstedt and Romig2009). In addition, for confirmation purposes and in case of ambiguous banding patterns, PCR products were purified using QIAquick PCR purification kit (Qiagen) following the manufacturer's guidelines. The purified amplicons were sent to Macrogen Europe BV (Amsterdam, the Netherlands) for sequencing using the nested reverse primer. The sequences were identified by comparing with those available in the National Centre for Biotechnology Information database (NCBI) using the basic local alignment search tool (http://www.ncbi.nlm.nih.gov/BLAST/) (Altschul et al., Reference Altschul, Madden, Schaffer, Zhang, Zhang, Miller and Lipman1997).
Results
Cyst condition and organ location of Echinococcus spp.
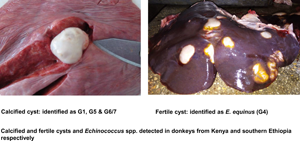
Out of 338 donkeys, 528 cystic structures were collected. Of these, 263 were characterized as Echinococcus spp. and 1 as a metacestode of Spirometra theileri. The remaining cysts either did not yield sequences or banding patterns of sufficient quality, or were excluded from molecular examination due to advanced degeneration with low probability of amplification success. Four E. granulosus s. l. species were detected: E. equinus (n = 163), E. granulosus s. s. (n = 70), E. canadensis (G6/7) (n = 26) and E. ortleppi (n = 4) (Tables 1 and 2). Details of representative sequences obtained in this study are available on GenBank under accession numbers OK489943–OK489955 (Table 3). Details of sequences from GenBank that were identical to those reported in this study are shown in Table 3. Fertility of E. equinus cysts was high (111/163), but low for the other species: 7/70 for E. granulosus s. s., 1/4 for E. ortleppi and 3/26 for E. canadensis (G6/7). The most frequently infected organs were the liver for E. equinus (115/163) and E. canadensis (G6/7) (15/26) and the lungs for E. granulosus s. s. (43/70), while 2/4 E. ortleppi cysts occurred in the lungs and the kidneys each (Table 1).
Table 1. Cyst condition and organ location of Echinococcus spp. from donkeys in eastern Africa

Table 2. Geographical origin and prevalence of Echinococcus spp. from donkeys in eastern Africa

EG, E. granulosus s. s.; EO, E. ortleppi; EC, E. canadensis G6/7; EE, E. equinus.
a Possible prevalence underestimate, as some positive animals were not included for which the information on the geographical origin was lost during carcass processing.
b Figures include donkeys of unknown origin (within the study region) and positive animals whose data on origin were lost during carcass processing.
Table 3. Echinococcus granulosus sensu lato representative nad1 sequences from donkeys in eastern Africa and details of identical sequences in the GenBank

a Representative reference sequence with closest geographic origin, others with same percentage identity exist.
Prevalence and geographical distribution of Echinococcus spp.
In total, 5961 donkeys were examined, of which 337 (5.7%) were infected with CE and 1 with a metacestode of S. theileri. The geographical origin of the donkeys is presented in Table 2 at regional and county level. For 614 donkeys, the origin was unknown from the onset. In addition, from those with known origin, in case of 77 of the 337 positive animals, this information was lost during carcass processing. These positive records were omitted from Table 2 (except for the grand total), resulting for some counties in prevalence underestimates (see footnote ‘a’ in Table 2). Generally, CE prevalence estimates were highest for animals from Karamoja (Uganda), northeastern Kenya and southern Ethiopia, and low for animals from central, western and southern Kenya and northern Tanzania. Geographical structuring was obvious for E. equinus, where all except 2 infected animals originated from southern Ethiopia. The other Echinococcus spp. were widely spread across the study area; all of the (few) characterized cyst from central Kenya were E. granulosus s. s., all cysts from Karamoja were E. canadensis (G6/7) (Table 2). The donkey infected with S. theileri originated from Kajiado county, southern Kenya.
Discussion
This study reports for the first time the presence and prevalence of CE in donkeys, and any member of the horse family, in eastern Africa. Data on CE in donkeys are few worldwide, e.g. from Turkey, Italy and Israel (Abo-Shehada, Reference Abo-Shehada1988; Mukbel et al., Reference Mukbel, Torgerson and Abo-Shehada2000; Thompson and McManus, Reference Thompson and McManus2002; Oge et al., Reference Oge, Kircali, Yildirim and Oge2004; Varcasia et al., Reference Varcasia, Garippa, Pipia, Scala, Brianti, Giannetto, Battelli, Poglayen and Micagni2008; Simsek et al., Reference Simsek, Roinioti and Eroksuz2015). In Africa, such reports are restricted to the North (Egypt, Tunisia and Morocco), while no data exist from sub-Saharan Africa despite the ubiquity of donkeys in many countries (Pandey, Reference Pandey1980; Azlaf and Dakkak, Reference Azlaf and Dakkak2006; Haridy et al., Reference Haridy, Gawad, Ibrahim, Hassan, El-Sherbi, El Shazly and Morsy2008; Taha, Reference Taha2012; Aboelhadid et al., Reference Aboelhadid, El-Dakhly, Yanai, Fukushi and Hassanin2013; Boufana et al., Reference Boufana, Lahmar, Rebai, Ben Safta, Jebabli, Ammar, Kachti, Aouadi and Craig2014; Lahmar et al., Reference Lahmar, Boufana, Jebabli, Craig, Ayari, Basti, Dhibi and Torgerson2014; Mahdy et al., Reference Mahdy, Abdel-Maogood, Abdel-Wahab and El-Bahy2014a, Reference Mahdy, Maogood, Wahab and El-Bahy2014b; Barghash et al., Reference Barghash, El Sayed, El-Alfy, Abou-Elnour, El-Kattan and Sadek2017; Desouky et al., Reference Desouky, Helmy, Sorour and Amer2017). The prevalence of donkey CE found in this study (5.7%) was close to that reported from northern Africa with 4.2% in Morocco (Pandey, Reference Pandey1980), 8.5% in Tunisia (Lahmar et al., Reference Lahmar, Boufana, Jebabli, Craig, Ayari, Basti, Dhibi and Torgerson2014) and 6.9–14.2% in Egypt (Haridy et al., Reference Haridy, Gawad, Ibrahim, Hassan, El-Sherbi, El Shazly and Morsy2008; Aboelhadid et al., Reference Aboelhadid, El-Dakhly, Yanai, Fukushi and Hassanin2013; Mahdy et al., Reference Mahdy, Maogood, Wahab and El-Bahy2014b; Barghash et al., Reference Barghash, El Sayed, El-Alfy, Abou-Elnour, El-Kattan and Sadek2017; Desouky et al., Reference Desouky, Helmy, Sorour and Amer2017). In addition to the scarcity of CE data from donkeys, few studies have identified the causative species of Echinococcus. Thus, E. equinus has been identified in Tunisia and Egypt (Aboelhadid et al., Reference Aboelhadid, El-Dakhly, Yanai, Fukushi and Hassanin2013; Boufana et al., Reference Boufana, Lahmar, Rebai, Ben Safta, Jebabli, Ammar, Kachti, Aouadi and Craig2014; Lahmar et al., Reference Lahmar, Boufana, Jebabli, Craig, Ayari, Basti, Dhibi and Torgerson2014; Desouky et al., Reference Desouky, Helmy, Sorour and Amer2017; Mousa et al., Reference Mousa, Abdel-Wahab, El-Gameel Sohila and Mahdy2020) and E. granulosus s. s. in Tunisia and Morocco (Azlaf, Reference Azlaf2007; Boufana et al., Reference Boufana, Lahmar, Rebai, Ben Safta, Jebabli, Ammar, Kachti, Aouadi and Craig2014; Lahmar et al., Reference Lahmar, Boufana, Jebabli, Craig, Ayari, Basti, Dhibi and Torgerson2014).
Echinococcus equinus is assumed to be the species most closely adapted to donkeys and other Equidae as intermediate hosts, reaching high levels of fertility. This was confirmed by 68% fertility among our samples of E. equinus. The distribution and host range of this species in sub-Saharan Africa are still rather enigmatic: in our study, almost all donkeys with E. equinus originated from southern Ethiopia, only 2 animals from Kenya. This is unexplained, but the scarcity of this species in Kenya is supported by its absence in recent large-scale faecal surveys of dogs (Mulinge et al., Reference Mulinge, Magambo, Odongo, Njenga, Zeyhle, Mbae, Kagendo, Addy, Ebi, Wassermann, Kern and Romig2018) and wild carnivores (Kagendo et al., Reference Kagendo, Magambo, Agola, Njenga, Zeyhle, Mulinge, Gitonga, Mbae, Muchiri, Wassermann, Kern and Romig2014) across the country. Older data from Chad (no case of CE in 163 horses and donkeys) suggest that this region of absence of scarcity of E. equinus may stretch from eastern to central Africa (Graber et al., Reference Graber, Troncy, Tabo, Service and Oumatie1969). No wild equids (zebras) have ever been examined for CE in eastern and central Africa. This is in clear contrast to the situation in southern Africa, where E. equinus is obviously common in plains zebras and wild carnivores in northern Namibia (Wassermann et al., Reference Wassermann, Aschenborn, Aschenborn, Mackenstedt and Romig2015), was reported from a mountain zebra in ‘South West Africa’ (Kumaratilake et al., Reference Kumaratilake, Thompson and Eckert1986), and where reports of 60% ‘incidence’ (sic) and fertility of cysts in plains zebra of Kruger National Park in South Africa also suggest the frequent presence of E. equinus there (Young, Reference Young1975a, Reference Young1975b). The latter is supported by a recent report of fertile cysts of E. equinus in a white rhino from Kruger National Park, RSA (Zaffarano et al., Reference Zaffarano, de Klerk-Lorist, Junker, Mitchell, Bhoora, Poglayen and Govender2021). There are no reports of E. equinus in donkeys, horses or domestic dogs from southern Africa. Although our data close a gap of knowledge by reporting the presence of E. equinus in eastern Africa, data on the obviously patchy geographical spread of this species on the continent are still far from complete. In particular, further surveys are needed to investigate whether the endemic area in southern Ethiopia is an isolated focus of this parasite, or is linked to the northern African endemic region. Reasons for differences in local frequency of this parasite are unclear. The principal hosts, donkeys and domestic dogs, are abundant in all parts of our study area. Even though in most parts of eastern Africa, donkeys are rarely slaughtered for human consumption and dogs may not get infected via slaughter offal, it is unlikely that carcasses of donkeys that died due to age or accidents are disposed in a way that they are out of reach for (stray) dogs or wild scavengers. Ongoing research on Echinococcus transmission in southern Ethiopia may shed light on the risk factors in that focus. This is not irrelevant, as E. equinus – long assumed to be apathogenic for humans – has recently been reported as causative agent of human CE (Kim et al., Reference Kim, Yong, Shin, Lee, Park, Suvonkulov, Kovalenko and Yu2020; Macin et al., Reference Macin, Orsten, Samadzade, Colak, Cebeci and Fındık2021), and the rarity of reported human cases may at least partly be due to the general rarity of the parasite in large parts of Africa (and the world).
The numbers of donkeys infected with Echinococcus spp. other than E. equinus (E. granulosus s. s. n = 70; E. canadensis G6/7 n = 26; E. ortleppi n = 4) do approximately reflect the relative frequencies of these species in their typical intermediate hosts (sheep, goats, camels and cattle) in the study area. In Kenya, E. granulosus s. s. is the most abundant species in sheep, goats and cattle followed by E. canadensis (G6/7) and E. ortleppi, this frequency can also be seen in donkeys. However, the prevalence of these 3 species in donkeys was far lower than in their ruminant hosts in the same region (Dinkel et al., Reference Dinkel, Njoroge, Zimmermann, Walz, Zeyhle, Elmahdi, Mackenstedt and Romig2004; Maillard et al., Reference Maillard, Benchikh-Elfegoun, Knapp, Bart, Koskei, Gottstein and Piarroux2007; Casulli et al., Reference Casulli, Zeyhle, Brunetti, Pozio, Meroni, Genco and Filice2010; Addy et al., Reference Addy, Alakonya, Wamae, Magambo, Mbae, Mulinge, Zeyhle, Wassermann, Kern and Romig2012; Hailemariam et al., Reference Hailemariam, Nakao, Menkir, Lavikainen, Yanagida, Okamoto and Ito2012; Mutwiri et al., Reference Mutwiri, Magambo, Zeyhle, Mkoji, Wamae, Mulinge, Wassermann, Kern and Romig2013; Mbaya et al., Reference Mbaya, Magambo, Njenga, Zeyhle, Mbae, Mulinge, Wassermann, Kern and Romig2014; Chamai et al., Reference Chamai, Omadang, Erume, Ocaido, Oba, Othieno, Bonaventure and Kitibwa2016; Tigre et al., Reference Tigre, Deresa, Haile, Gabriel, Victor, Pelt, Devleesschauwer, Vercruysse and Dorny2016; Odongo et al., Reference Odongo, Tiampati, Mulinge, Mbae, Bishop, Zeyhle, Magambo, Wasserman, Kern and Romig2018; Nungari et al., Reference Nungari, Mbae, Gikunju, Mulinge, Kaburu, Zeyhle and Magambo2019; Terefe et al., Reference Terefe, Addy, Alemu, Mackenstedt, Romig and Wassermann2019; Tamarozzi et al., Reference Tamarozzi, Kibona, de Glanville, Mappi, Adonikamu, Salewi, Misso, Maro, Casulli, Santoro, Santolamazza, Mmbaga and Cleaveland2022). Also, cyst fertility rates in donkeys [10% for E. granulosus s. s., 12% for E. canadensis (G6/7), 1/4 for E. ortleppi] were far lower compared to their typical hosts. In case of E. granulosus s. s., this is in accordance with studies from the Mediterranean area, where few or no cysts of this species were found fertile in donkeys and horses (Azlaf, Reference Azlaf2007; Varcasia et al., Reference Varcasia, Garippa, Pipia, Scala, Brianti, Giannetto, Battelli, Poglayen and Micagni2008; Utuk and Simsek, Reference Utuk and Simsek2013; Boufana et al., Reference Boufana, Lahmar, Rebai, Ben Safta, Jebabli, Ammar, Kachti, Aouadi and Craig2014; Lahmar et al., Reference Lahmar, Boufana, Jebabli, Craig, Ayari, Basti, Dhibi and Torgerson2014), indicating a poor host adaptation. For E. canadensis (G6/7) and E. ortleppi, the presented results are even the first confirmation that these species can infect donkeys and reach fertility there; apart from our findings, an E. ortleppi cyst of unknown fertility status previously reported from an unspecified species of zebra in Namibia was the only record of E. ortleppi from any member of the Equidae (Obwaller et al., Reference Obwaller, Schneider, Walochnik, Gollackner, Deutz, Janitschke, Aspock and Auer2004). Yet, given the low prevalence and fertility rate, donkey infections with E. granulosus s. s., E. canadensis (G6/7) and E. ortleppi are likely the result of spillover from the typical lifecycles between dogs and domestic ruminants, and donkeys probably play a small role, if any, in the transmission of these parasites.
This study reports the unusual finding of S. theileri calcified cyst in a donkey that was located in the kidney and initially misidentified as a degenerated Echinococcus cyst. Domestic and wild carnivores serve as the definitive hosts of Spirometra spp., while the first intermediate hosts are copepods and the second intermediate hosts are amphibians, reptiles or herbivorous mammals (paratenic hosts). A sylvatic cycle for Spirometra spp. involving wild herbivores (zebra, warthog and antelope) as second intermediate hosts (paratenic) and carnivores (hyenas) was proposed in Maasai Mara (Nelson et al., Reference Nelson, Pester and Rickman1965; Muller-Graf, Reference Muller-Graf1995). A small number of human infections with plerocercoids of Spirometra spp. are known from Kenya (Schmid and Watschinger, Reference Schmid and Watschinger1972; MOH, 2016). Although the zoonotic potential of S. theileri is unknown, infections with adult worms of this species have been found in leopards and spotted hyenas in Tanzania and recently in domestic dogs in Maasai Mara (Eom et al., Reference Eom, Park, Lee, Choe, Kang, Bia, Ndosi, Nath, Eamudomkarn, Keyyu, Fyumagwa, Mduma and Jeon2019; Mulinge et al., Reference Mulinge, Zeyhle, Mpario, Mugo, Nungari, Ngugi, Sankale, Gathura, Magambo and Kachani2021). As the plerocercoid found in our donkey was calcified, the significance of this finding is unclear, and the lifecycle of S. theileri in the area is in need of investigation.
Data availability
Data supporting results are provided within the article and available on GenBank under accession numbers OK489943-OK489955.
Acknowledgements
The authors sincerely thank the management of the 3 slaughterhouses in Naivasha, Mogotio and Lodwar for permission to access and collect of cysts from donkeys. The meat inspectors are appreciated for their corporation during the sampling period. This manuscript is published with permission from the Director General, KEMRI.
Author contributions
E. M., E. Z., P. K., M. W., U. M. and T. R. conceived and designed the study. E. M., E. Z. and J. M. carried out fieldwork. E. M., C. M., L. G., T. K. and M. W. performed laboratory analysis. All authors wrote and revised the manuscript.
Financial support
This study was funded by Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) – KE282-9/1, -12/1, RO3753-3/1, -9/1 – and conducted in the context of the ‘Cystic Echinococcosis in sub-Saharan Africa Research initiative (CESSARi)’.
Conflict of interest
None.
Ethical standards
The KEMRI Scientific Ethics Review Unit (SERU) approved this study SSC. No. 1684 as well as the Department of Veterinary Services, Kenya.





 Open access
Open access