Introduction
Estimates of population size for species, mainly threatened or endemic, is one of the fundamental and necessary steps in carrying out risk categorisations both nationally and globally. Population size estimates can also support the development of management guidelines, guide conservation initiatives, and is the first step in beginning trend monitoring (Wolfaardt and Phillips Reference Wolfaardt and Phillips2011; Mills Reference Mills2012; Marques et al. Reference Marques, Thomas, Martin, Mellinger, Ward and Moretti2013; Renjifo et al. Reference Renjifo, Gómez, Tibatá, Villarreal, Kattan and Espine2014; Ortega‑Álvarez et al. Reference Ortega‑Álvarez, Calderón‑Parra, Martínez Molina, Martínez Molina, Martínez Molina and Martínez Molina2021). The Antioquia Brushfinch Atlapetes blancae is a Colombian endemic with a distribution restricted to the northern end of the Central Andes in Antioquia (Donegan Reference Donegan2007; Chaparro-Herrera et al. Reference Chaparro-Herrera, Echeverry-Galvis, Córdoba-Córdoba and Sua-Becerra2013). Given its restricted distribution and the rapid habitat transformation across its entire range, it is classified both locally and globally as a “Critically Endangered” species (Chaparro-Herrera Reference Chaparro-Herrera, Renjifo, Gómez, Velásquez-Tibatá, Amaya-Villarreal, Kattan, Amaya-Espinel and Burbano-Girón2014; BirdLife International 2021). In this study, we provide the first population size estimates of Antioquia Brushfinch using mark-resighting in seven areas in which the species had been previously recorded. We also provide the first accurate estimates of sex ratio, which is another important demographic parameter that influences population survival and can critically affect parental care, mate acquisition, and more complicated consequences that determine population trends.
Antioquia Brushfinch was first described by Donegan (Reference Donegan2007) based on three specimens misidentified as Slaty Brushfinch Atlapetes schistaceus collected in San Pedro de los Milagros in 1971 and deposited in different Colombian museum collections. Efforts to find this remnant population were unsuccessful until 2018 when it was rediscovered 47 years after the collection of the only dated specimen among the holotype and paratypes (Donegan Reference Donegan2007; Correa et al. Reference Correa, Chaparro-Herrera, Lopera-Salazar and Parra2019). After its rediscovery, there has been some research focused on contributing to the knowledge of the species’ natural history, including studies of habitat preferences, diet, behaviour, and reproduction (Correa et al. Reference Correa, Chaparro-Herrera, Lopera-Salazar and Parra2019; Chaparro-Herrera and Lopera-Salazar Reference Chaparro-Herrera and Lopera-Salazar2019; Valencia-C et al. Reference Valencia-, Sánchez-Londoño, Villamizar and Ángel2019; Chaparro-Herrera et al. Reference Chaparro-Herrera, Hernández Schmidt and Lopera-Salazar2021). Although the species might use a wide variety of habitats (e.g. secondary vegetation; García-Ospina et al. Reference García-Ospina, Ocampo-Peñuela and Parra2023), current natural history information suggests that it is a shrubland specialist (Correa Reference Correa, Chaparro-Herrera, Lopera-Salazar and Parra2019; Donegan Reference Donegan2007; Donegan et al. Reference Donegan, Huertas and Flórez2009), including habitats such as open mesophilic shrubland.
The population of Antioquia Brushfinch is estimated to be fewer than 50 mature individuals with a decreasing or unknown population trend, based on assumptions of its restricted range and land-use patterns throughout its distribution (Chaparro-Herrera Reference Chaparro-Herrera, Renjifo, Gómez, Velásquez-Tibatá, Amaya-Villarreal, Kattan, Amaya-Espinel and Burbano-Girón2014; BirdLife International 2021). Systematic efforts to obtain a standardised estimate of population size have been lacking (but see Díaz-Pulido et al. Reference Díaz-Pulido, Chiquito-García, Calderón-Franco, Múnera, Mazo and Ochoa2020). Its threat classification was mainly based on the original description and the lack of success in recording the species for many years after it was first described (Donegan et al. Reference Donegan, Huertas and Flórez2009; Renjifo et al. Reference Renjifo, Gómez, Tibatá, Villarreal, Kattan and Espine2014). Similarly, BirdLife International (2021) made a reclassification for the International Union for Conservation of Nature and Natural Resources (IUCN), and despite having more information available for its evaluation, it was again classified as “Critically Endangered”, and data on its population status and trends remained uncertain and speculative.
In addition to population size estimates, another important demographic parameter that influences population survival is adult sex ratio. Differences in the proportion of sexes in a population may affect parental care, mate acquisition, and other indirect consequences that determine population trends. Unbalanced sex ratios may be a consequence of numerous factors, such as differential adult or juvenile survival and sex ratio at birth, that can reflect underlying intrinsic and extrinsic problems faced by populations. It has also been shown that adult sex ratio may also determine the risk of population extinction (Ancona et al. Reference Ancona, Dénes, Krüger, Székel and Beissinger2017). Owing to the long history of deforestation and habitat transformation throughout its range due to the extension of agricultural lands (e.g. potato plantations and pastures; González-Caro and Vásquez Reference González-Caro, Vásquez, Quintero, Benavides, Moreno and González-Caro2017; Negret et al. Reference Negret, Maron, Fuller, Possingham, Watson and Simmonds2021), Antioquia Brushfinch populations may show unbalanced sex ratios and other genetic consequences due to presumable historical population size contractions.
Our study provides the first population size estimates of Antioquia Brushfinch using mark-resighting data in seven areas in which the species had been previously recorded and estimates the adult sex ratio for this species based on genetic analyses of our sample. We also estimate the population size in fragments with previously known occurrence using an extrapolation based on abundance–area relationships. Our goal is to continue contributing scientific information about the species that can help to improve conservation actions and management decisions. We do not attempt to estimate total population size for the species, since the species has been recorded in areas other than the ones studied here. This is also the first step towards setting up a long-term monitoring programme to estimate population trends and other important demographic parameters such as recruitment, mortality, and dispersal.
Methods
Study area
The study took place at seven sites in the municipalities of San Pedro de los Milagros (four sites) and Santa Rosa de Osos (three sites), Antioquia, within the Altiplano de Santa Rosa de Osos, at the northern end of the Central Cordillera of Colombia, between 2,482 and 2,770 m a.s.l. (Figure 1). The main vegetation of the areas inhabited by Antioquia Brushfinch corresponds to low shrubs and low-growing forests, within the very humid Lower Montane Forest (vhf-ML) life zone (Holdridge Reference Holdridge1967; Espinal Reference Espinal, Callejas and Idárraga2011). Suitable vegetation for Antioquia Brushfinch is found naturally in places with poor or acid soils with superficial, stony or rocky outcrops and exposed to high solar radiation, strong wind, and extremely variable temperature (Ferreyra et al. Reference Ferreyra, Cingolani, Ezcurra and Bran1998; Avellaneda-Cadena and Betancur Reference Avellaneda-Cadena and Betancur2007; Chaparro-Herrera et al. Reference Chaparro-Herrera, Hernández Schmidt and Lopera-Salazar2021). The vegetation is dominated by 3–6 m tall shrubs, with scattered trees, that may, in some cases form small forest relicts dominated by Colombian Oak Quercus humboldtii that can reach up to 12 m tall (Chaparro-Herrera and Gómez Reference Chaparro-Herrera, Gómez and Billerman2022; Chaparro-Herrera et al. Reference Chaparro-Herrera, Hernández Schmidt and Lopera-Salazar2021).

Figure 1. Map showing the location of the seven sampling sites to estimate population size and sex ratio of Antioquia Brushfinch Atlapetes blancae. The colour gradient represents elevation in metres above sea level. White polygons show the regions containing patches of suitable habitat and observation records used for the extrapolation of population size. The inset shows a map of Colombia in black, the map of the department of Antioquia in grey, and the approximated distribution of Antioquia Brushfinch and our sampling area in white. Numbers by localities show the amount of suitable habitat available in each site.
Bird banding and resighting
To estimate population size at the seven focal localities (Figure 1), we designed a mark-resighting experiment in which we colour banded individuals at each site using a unique band combination by individual and site based on one or two bands. To band individuals, we travelled across the entire available habitat attempting to search and capture individuals using mist-nets upon detection. Mist-nets were run during six days at each location and captures were made between May and July 2022.
To ensure the closed population assumption of the model used to estimate populations size (see below), two weeks after banding individuals, we returned to the sites and travelled a standardised transect (route and speed), attempting to cover as much suitable habitat as possible (see Table 1 for sampling effort). We assumed that it was unlikely that during the period between mark and resighting individuals left the population (by immigration or death), or marks were lost. During the route, all detections were recorded along with information about habitat, date, and time, number of individuals detected, band combination (in the case of banded individuals), and coordinates. At the end of the transect we obtained the total number of individuals observed and the number of banded individuals within the sample. The sampling was made without replacement and each banded individual counted a single time per sampling event. The first author (SC-H) walked the transects between 05h50 and 11h00 and 14h00 and 18h00 avoiding sampling during rainy or extremely windy days. The starting point was alternated among replicates to reduce sampling bias. Each transect was sampled between three and eight times per locality (Table 1).
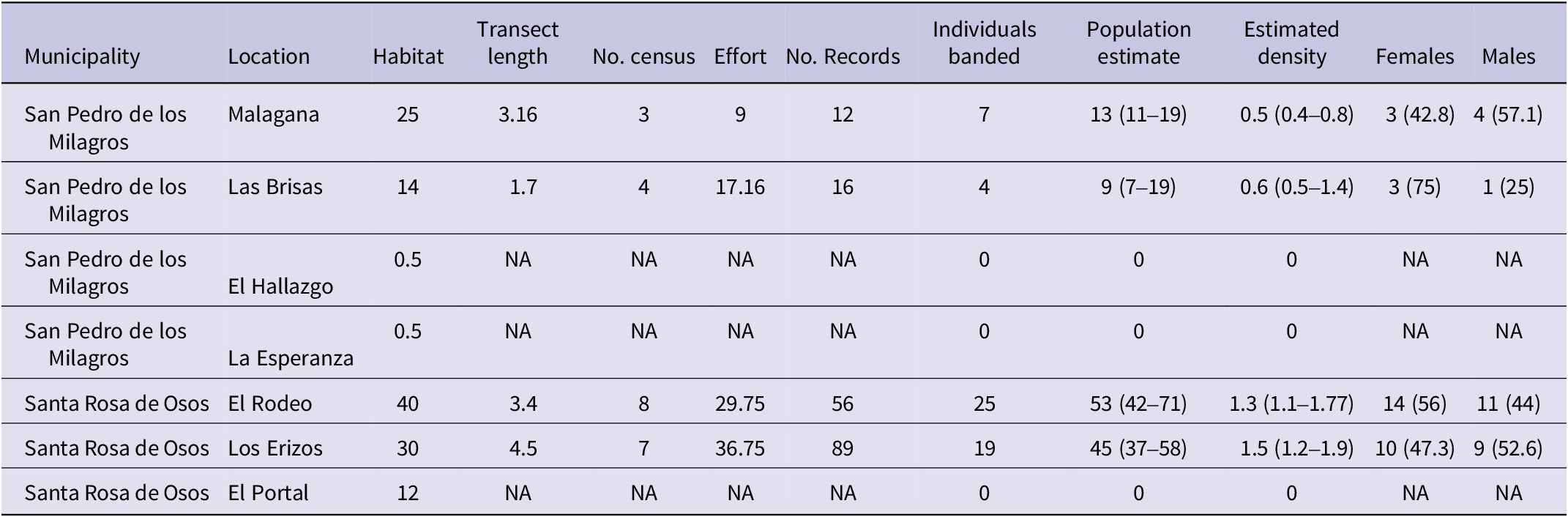
Table 1. Summary of localities sampled, available habitat per site, sampling effort, and results of the population estimate and sex determination of Antioquia Brushfinch Atlapetes blancae in seven localities in Antioquia. Habitat amount is expressed in hectares, transect length in kilometres, effort in hours, and density in individuals/ha. The 95% confidence intervals (CIs) of population estimates and density are shown in parenthesis. Finally, the proportion of each sex is shown in parenthesis in the “Females” and “Males” columns. “No. census” indicates the number of times a transect was sampled, and “No. records” indicates the number of individuals both banded and non-banded recorded across all censuses. Sites in which the population estimate is 0 correspond to the sites in which we did not find any individuals of Antioquia Brushfinch.
Estimation of sex ratio
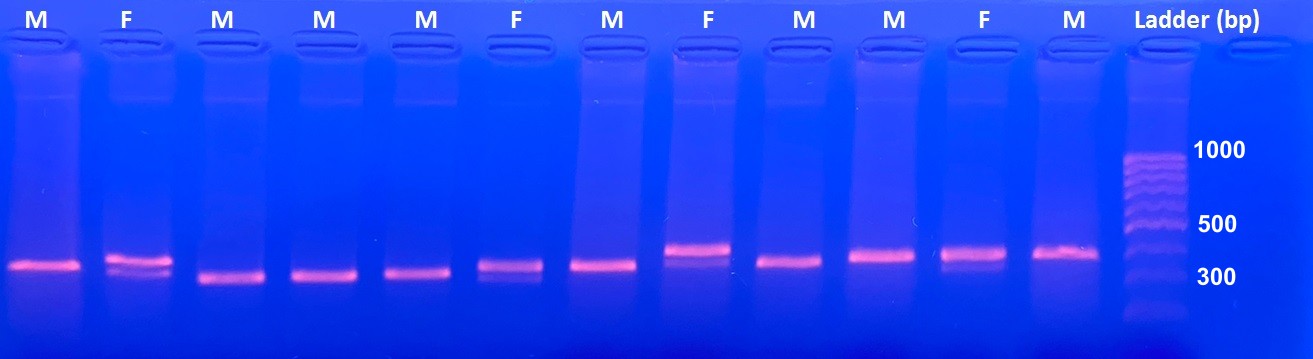
We extracted genomic DNA from blood samples preserved in absolute alcohol obtained from the 55 individuals banded. Individuals were sexed by amplifying the Z and W-linked genes Chromo-Helicase DNA-binding gene (CHD) (Griffiths et al. Reference Griffiths, Double, Orr and Dawson1998), using the P2 and P8 primers described in Griffiths and Tiwari (Reference Griffiths and Tiwari1995). Amplification followed the protocol described below. After extraction, 10 × polymerase chain reaction (PCR) buffer was added to the extraction along with 2 mM MgCl2, 2 mM deoxynucleotide triphosphate (dNTP), 2 μM of each primer, and 5 U/μL Taq DNA polymerase (ThermoFisher Scientific, Waltham, MA, USA). Amplification of the CHD gene included an initial denaturation at 94°C for four minutes, followed by 40 cycles of denaturation at 94°C for 30 seconds, an annealing temperature of 51°C for 45 seconds, extension 72°C for 45 seconds, and a final extension at 72°C for five minutes. The PCR products were run on a 2% agarose gel for 40 minutes at 80 V in Tris-borate-ethylenediaminetetraacetic acid (EDTA) buffer (TBE). Individuals with two bands in the gel were identified as females while individuals with only one band were identified as males (see Supplementary material Table S1). Amplification of the W- and Z-linked genes for sex identification was performed twice for each sample to improve precision.
Data analysis
Population size estimates
We used the well-known hypergeometric closed population mark-resighting model to estimate population size in each site (Seber Reference Seber1982). With a single sample (two visits, the first one to mark the animals and the second one to recapture or resight), this model leads to the Lincoln–Petersen Index estimator of population sizes. As Seber (Reference Seber1982) explains, the likelihood function, or joint probability of the observations can easily be adapted when
 $ k $
replicated counts are available, as in our case. Let
$ k $
replicated counts are available, as in our case. Let
 $ {x}_i $
, be the number of marked individuals that were observed during the i th replicate of a transect at a site. This number was again assumed to be a sample from a hypergeometric distribution, which depends on the unknown total population size
$ {x}_i $
, be the number of marked individuals that were observed during the i th replicate of a transect at a site. This number was again assumed to be a sample from a hypergeometric distribution, which depends on the unknown total population size
 $ t $
, the total number of marked individuals
$ t $
, the total number of marked individuals
 $ {m}_i $
present in the population during that i th replicated count, and
$ {m}_i $
present in the population during that i th replicated count, and
 $ {n}_i $
, the total number of individuals (marked and unmarked) that were observed during each one of the
$ {n}_i $
, the total number of individuals (marked and unmarked) that were observed during each one of the
 $ i $
replicates of the transect. Then, the joint probability total number of individuals in each site was obtained by maximising the joint likelihood function of the replicated counts, given by:
$ i $
replicates of the transect. Then, the joint probability total number of individuals in each site was obtained by maximising the joint likelihood function of the replicated counts, given by:
 $$ L(t)=\prod \limits_{i=1}^k\frac{\left(\begin{array}{c}{m}_i\\ {}{n}_i\end{array}\right)\left(\begin{array}{c}t-{m}_i\\ {}{n}_i-{x}_i\end{array}\right)}{\left(\begin{array}{c}t\\ {}{n}_i\end{array}\right)} $$
$$ L(t)=\prod \limits_{i=1}^k\frac{\left(\begin{array}{c}{m}_i\\ {}{n}_i\end{array}\right)\left(\begin{array}{c}t-{m}_i\\ {}{n}_i-{x}_i\end{array}\right)}{\left(\begin{array}{c}t\\ {}{n}_i\end{array}\right)} $$
The maximum likelihood estimates of the population size
 $ \hat{t} $
is the value of the unknown constant
$ \hat{t} $
is the value of the unknown constant
 $ t $
that maximises the joint probability of the observations under this hypergeometric model, i.e. the value of the unknown population size that renders the data most likely under this probabilistic model of sampling without replacement. Extensive simulations in Taper et al. (Reference Taper, Lele, Ponciano, Dennis and Jerde2021) have shown that the statistical properties (bias and variance) of this estimator are quite robust to departures from the hypergeometric model assumptions. Similarly, the confidence interval (CI) with best coverage for this model was found to be the profile likelihood CI (Taper et al. Reference Taper, Lele, Ponciano, Dennis and Jerde2021). Consequently, the 95% CIs for the population estimates were obtained using profile likelihood methods (Pawitan Reference Pawitan2001). For each locality, based on the information about observations and number of marked individuals, we evaluated the likelihood of the hypergeometric model for values of
$ t $
that maximises the joint probability of the observations under this hypergeometric model, i.e. the value of the unknown population size that renders the data most likely under this probabilistic model of sampling without replacement. Extensive simulations in Taper et al. (Reference Taper, Lele, Ponciano, Dennis and Jerde2021) have shown that the statistical properties (bias and variance) of this estimator are quite robust to departures from the hypergeometric model assumptions. Similarly, the confidence interval (CI) with best coverage for this model was found to be the profile likelihood CI (Taper et al. Reference Taper, Lele, Ponciano, Dennis and Jerde2021). Consequently, the 95% CIs for the population estimates were obtained using profile likelihood methods (Pawitan Reference Pawitan2001). For each locality, based on the information about observations and number of marked individuals, we evaluated the likelihood of the hypergeometric model for values of
 $ t $
between 1 and 200 and divided each likelihood value by the maximum value in the set to obtain the relative likelihood. The maximum likelihood estimates of the population size
$ t $
between 1 and 200 and divided each likelihood value by the maximum value in the set to obtain the relative likelihood. The maximum likelihood estimates of the population size
 $ \hat{t} $
is the value for which the relative likelihood is equal to 1 and the boundaries of the 95% CIs are defined by the values at which the relative likelihood is ~0.15 (Pawitan Reference Pawitan2001; Ponciano et al. Reference Ponciano, Taper, Dennis and Lele2009; Taper et al. Reference Taper, Lele, Ponciano, Dennis and Jerde2021). All analyses were performed in R version 4.2.2 (R Core Team Reference Team2022).
$ \hat{t} $
is the value for which the relative likelihood is equal to 1 and the boundaries of the 95% CIs are defined by the values at which the relative likelihood is ~0.15 (Pawitan Reference Pawitan2001; Ponciano et al. Reference Ponciano, Taper, Dennis and Lele2009; Taper et al. Reference Taper, Lele, Ponciano, Dennis and Jerde2021). All analyses were performed in R version 4.2.2 (R Core Team Reference Team2022).
Extrapolation of population size
To extrapolate the potential number of individuals across a broader geographical scope than our seven localities, we constructed a Poisson Generalised Linear Model (GLM) with the result of the population estimates for each of the seven localities as dependent variable and the total amount of suitable habitat as independent variable. In this model, we did not estimate an intercept (only a slope) since we assumed that the species is absent when there is no habitat available. Using the slope estimates of this regression, we then predicted the number of potential individuals in the localities in which we have field confirmation of the presence of the species (SC-H and Atlapetes Project: https://arcg.is/1yz0eK) (n = 13). To propagate the error in the estimation of population size into our regression model, we used a parametric bootstrap procedure with 1,000 replicates. The bootstrap experiment consisted of simulating data of the number of marked individuals in each observation replicate, using the estimated number of individuals in each patch, and the total number of marked individuals in the population. Using this population size estimate for each simulation iteration, we constructed the Poisson GLM to estimate the relationship between population size and area. Regression and bootstrap experiment were constructed using software R. The amount of habitat available in each of the 20 localities (seven sampled during this study and 13 in which we had confirmed presence of the species by the Atlapetes Project) was estimated by manually constructing polygons around areas with suitable vegetation over satellite images taken in year 2020 in Google Earth Pro 7.3.6.9285 (https://www.google.com/intl/es/earth/about/versions/#earth-pro). To construct the polygons, we relied on GPS coordinates of a record of Antioquia Brushfinch and manually traced the forested area around the GPS coordinate. Since the region has been mostly transformed to pastures or potato crops, forested or shrubby areas in which the species is present are clearly identified by a darker green colour in the satellite image. Polygons were constructed using the QGIS version 3.26.2 (QGIS 2021); see Figure 1 for areas in which polygons were constructed.
Sex ratio
Sex ratio was estimated by dividing the total number of males in the sample by the total number of banded individuals (Ancona et al. Reference Ancona, Dénes, Krüger, Székel and Beissinger2017). To determine any biases in the sex ratio we used a binomial exact test using as null hypothesis that the probability of catching a male from the population is 0.5 and a two-tailed alternative hypothesis testing if sex ratio is biased towards males or females irrespectively.
Results
Population size
To estimate population size, we banded 55 individuals across four locations: 4 in Las Brisas, 7 in Malagana (both in San Pedro de los Milagros), 19 in Los Erizos, and 25 in El Rodeo (the latter two in Santa Rosa de Osos). We were unable to record the species in three other sampling areas, El Hallazgo, La Esperanza, and El Portal, despite having been recorded in the past. Of the total number of banded individuals, 6 were juveniles, 12 immatures, and 37 adults. We performed a total of 23 censuses in the four localities in which we had previously banded individuals (Table 1). Population estimates suggest that there are 120 (95% CI: 97–167) individuals summed across the four localities in which we had at least one banded individual. The locality with the largest population was El Rodeo and the smallest population was found at Las Brisas (Table 1). We found however the highest density at Los Erizos and lowest population density at Malagana. The total amount of habitat available was largest in El Rodeo and smallest in La Esperanza and El Hallazgo (Table 1).
Extrapolation of population size
In addition to the seven areas in which we directly estimated population size based on the mark-resighting experiment, we measured available habitat in 13 more areas with confirmed presence of Antioquia Brushfinch. In total, the 20 sites summed 322.6 ha of suitable habitat. As expected, the GLM suggested that population size significantly increased with availability of suitable habitat (
 $ \# individuals={e}^{\left(0.11\;\left(95\% CI\;0.1-0.11\right)\ast area\right)} $
) (Figure 2). For the 322.6 ha for which we confirmed habitat availability, we predicted that population size is 405 (95% CI: 335–523) individuals. Since we did not find individuals in our sampling sites with fewer than 12 ha (Table 1, Figure 2) a conservative population size estimate for the areas with known presence of the species and more than 12 ha of suitable habitat was 394 (95% CI: 323–510).
$ \# individuals={e}^{\left(0.11\;\left(95\% CI\;0.1-0.11\right)\ast area\right)} $
) (Figure 2). For the 322.6 ha for which we confirmed habitat availability, we predicted that population size is 405 (95% CI: 335–523) individuals. Since we did not find individuals in our sampling sites with fewer than 12 ha (Table 1, Figure 2) a conservative population size estimate for the areas with known presence of the species and more than 12 ha of suitable habitat was 394 (95% CI: 323–510).
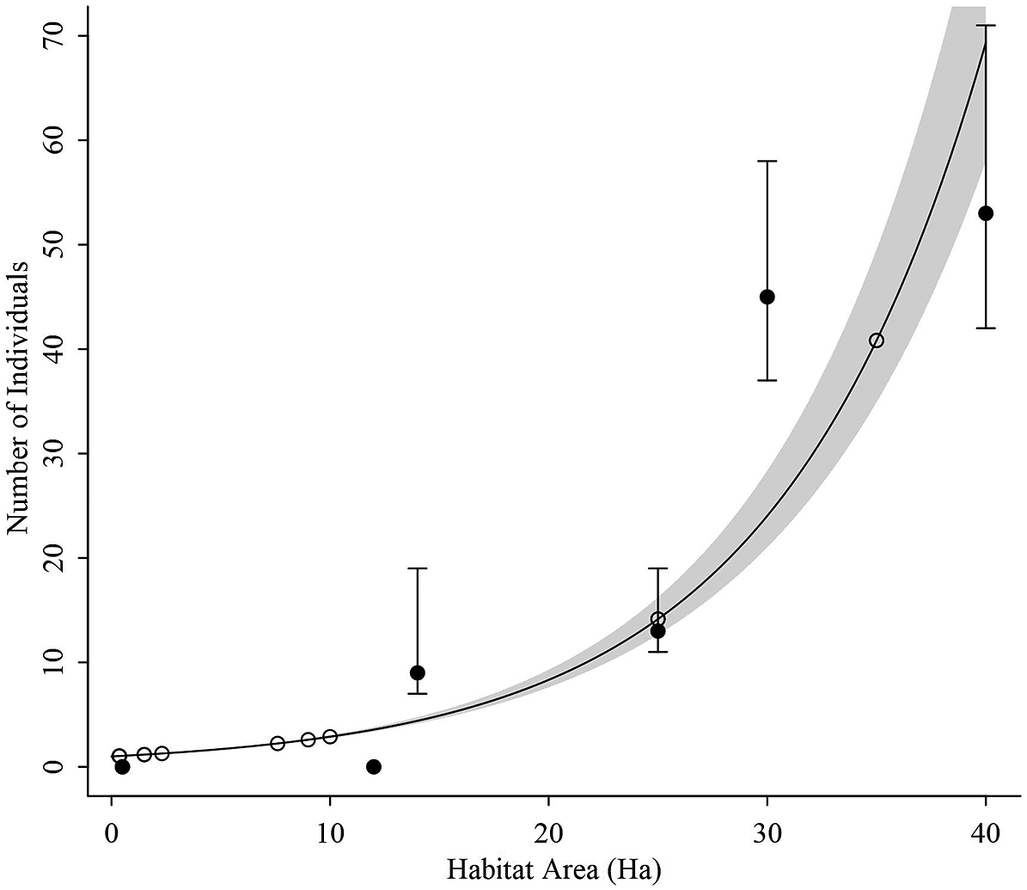
Figure 2. We estimated the expected population size of Antioquia Brushfinch Atlapetes blancae in 15 localities with suitable habitat from which we did not have direct mark-resighting estimates, but we had confirmed presence for the species. In the figure, black dots and error bars show population estimates and their 95% confidence intervals (CIs) form the mark-resighting experiment. Open circles show the prediction of the population size based on the individuals–area relationship,
 $ \# individuals={e}^{\left(0.11\;\left(95\% CI\;0.1-0.11\right)\ast area\right)} $
. The black line shows the maximum likelihood estimate of the relationship between number of individuals and area based on a Generalised Linear Model with Poisson distributed error and the grey polygon shows its 95% CI.
$ \# individuals={e}^{\left(0.11\;\left(95\% CI\;0.1-0.11\right)\ast area\right)} $
. The black line shows the maximum likelihood estimate of the relationship between number of individuals and area based on a Generalised Linear Model with Poisson distributed error and the grey polygon shows its 95% CI.
Adult sex ratio
Results from PCR showed a single defined band in males of approximately 316 base pairs (bp) and two bands in females of 316 bp and 419 bp (Figure S1). From the 55 banded birds 30 were females (54.54%) and 25 males (45.55%). The two PCR replicates were fully congruent, demonstrating high effectiveness in the primers used (Table S1). The binomial exact test did not reject the null hypothesis suggesting that sexes are balanced across all localities with a probability of an individual being a male of 0.45 (95% CI: 0.3–0.6; P = 0.4). Analysing individual sites yielded similar results to the combined analysis. Finally, 37 (67.27%) individuals banded were adults and 18 (32.72%) were juveniles or immature. From the 18 young and immature individuals captured, 14 (77.7%) were females, and only 4 (22.2%) were males. In this case, the binomial exact test rejected the null hypothesis of a balanced sex ratio with a probability of an individual being a male of 0.22 (95% CI 0.06–0.48; P = 0.03).
Discussion
Antioquia Brushfinch is a “Critically Endangered” microendemic species restricted to the mountains in the northern end of the Central Andes in Colombia. In this study, for the first time, we provide field evidence of population size and sex ratio. Furthermore, we estimate population size in 13 additional areas for which we do not have direct population estimates but have estimates of habitat availability. Our results suggest that the population is substantially larger than the 50 individuals suggested by previous evaluations based on indirect evidence. Specifically, we report that field estimates from the seven sites suggest that there are 120 individuals in 122 ha of suitable habitat. Furthermore, we estimate that the total population in the 20 sites with a known presence of the species is 405 individuals. The estimated sex ratio is balanced (0.45 95% CI 0.3–0.6; P = 0.4) suggesting that the species has not significantly suffered from its reduced population size, but there might be a bias towards females in juveniles and immature individuals. The very high proportion of young birds in the netting sample is encouraging because it indirectly suggests successful ongoing reproduction and individual turnover.
The history of this species had previously suggested that the population size was extremely small. Indirect information led to the assumption that the total population of this species was smaller than 50 individuals (Renjifo et al. Reference Renjifo, Gómez, Tibatá, Villarreal, Kattan and Espine2014; BirdLife International 2021). We provide evidence for a larger than 50 individuals population size given that we banded 55 individuals in total. Our maximum likelihood estimation of the total population size and its CIs support a population size well above 50 individuals: The lower boundary of the 95% CI of the population estimate was 97, the maximum likelihood estimate was 120 and the upper CI was at 167.
The areas for which we were able to estimate population size based on the area–abundance relationship corresponds to the fraction of the species’ distribution with the most fragmented habitat remnants. In this area, mean patch size of suitable habitat in which the species is present is 22.4 ha. The largest amount of habitat available is in the northernmost portion of the distribution in which the species has been reported but our research group unfortunately has the least amount of information. In this area we estimate that there are at least 955.26 ha of suitable habitat for which mean patch size is 112.19 ha. Such distribution of habitat would suggest a large population. Unfortunately, since the largest fragment in which we field estimated population size was 40 ha, we were unable to extrapolate the population size for these larger fragments. It is clear from the data that the asymptote of the population size–abundance relationship is far from being reached, hampering our ability to properly extrapolate our abundance–area curve (Figure 2). This suggests the need to systematically sample larger fragment sizes to estimate correctly the total population size in the fragments of the northernmost portion of the distribution. In any case, the amount of habitat available around the sites with direct field estimates suggests that the total population of Antioquia Brushfinch is at least eight times larger than previously presumed (50 individuals or fewer; BirdLife International 2021).
We were unable to find any individuals of Antioquia Brushfinch in three of the seven localities sampled, even though they were selected based on previous records of the species on the sites. Given that habitat availability in two of these sites was less than one hectare (El Hallazgo and La Esperanza in Table 1), we speculate that historical records of the species in these areas represent movement of individuals between habitat fragments rather than established breeding populations in these small fragments. In a similar case, Krabbe (Reference Krabbe2004) reported that Pale-headed Brushfinch Atlapetes pallidiceps was absent from small suitable habitat fragments, but also speculated that these patches may harbour solitary or a single pair of individuals that had not yet been studied. Furthermore, given that we were unable to detect the species at El Portal, despite having at least 12 ha of suitable habitat, suggests that our extrapolation needs to be taken carefully, particularly in areas with fewer than 12 ha of suitable habitat. Nonetheless, the difference in the estimated number of individuals in all areas (405 individuals) versus the estimated number of individuals in areas with more than 12 ha of suitable habitat (394 individuals) is only 11 individuals larger. There certainly are other more important factors regulating the abundance of the species in small habitat fragments. Some of these factors may include habitat isolation (Bailey et al. Reference Bailey, Schmidt‐Entling, Eberhart, Herrmann, Hofer and Kormann2010), demographic stochasticity (Hanski Reference Hanski1999), and stronger impact of brood parasitism and nest predation due to habitat fragmentation and edge effects (Donovan et al. Reference Donovan, Jones, Annand and Thompson1997).
Our results are in line with other studies from a closely related species in Ecuador with a similar population and distribution range size (i.e. Pale-headed Brushfinch). The average territory size of Pale-headed Brushfinch is reported to be 1.0 ha (0.7–1.4 ha) based on direct observations of singing males (Krabbe Reference Krabbe2004). Although our study design does not allow us to directly estimate territory size, we can infer from the population estimate and size of fragments in which we performed our mark-resighting experiment that territory size is similar in Antioquia Brushfinch (0.5–1.5 individuals/ha). Given these similarities in population, distribution, and territory size, we speculate that the information and measures already taken to preserve Pale-headed Brushfinch populations may be applicable to Antioquia Brushfinch.
The negative consequences of small populations often include reduced genetic diversity and sex ratios departing from 1:1. We found that sex ratio was roughly balanced between males and females, when pooling both adults and immature individuals, suggesting that the population has not yet suffered from these problems related to small population sizes (Ancona et al. Reference Ancona, Dénes, Krüger, Székel and Beissinger2017). We note however, that the proportion of males within juvenile and immature individuals may suggest a different scenario. We found that 77% of the juvenile and immature individuals were females, probably suggesting that there is a higher survival of females at younger stages, but this is not reflected in the adult population (Clutton-Brock et al. Reference Clutton-Brock, Albon and Guinness1986; Martín et al. Reference Martín, Alonso, Alonso, Palacín, Magana and Martín2007). This result should be carefully interpreted since there are many other reasons to explain why captures would be sex-biased (e.g. small sample size: 18 immature and juvenile individuals). Further investigations about sex ratio in individual populations and genetic diversity are needed to further assess the genetic consequences of population size and estimate potential problems from inbreeding depression (Frankham Reference Frankham2005; Willoughby et al. Reference Willoughby, Sundaram, Wijayawardena, Kimble, Ji and Fernandez2015).
Despite the results obtained here, Antioquia Brushfinch is a species that we predict can persist provided the habitat is protected. It is worth noting though, that the species still has a small population, albeit bigger than previously thought, it has an extremely restricted distribution, and its habitat is being transformed at a tremendous speed (González-Caro and Vásquez Reference González-Caro, Vásquez, Quintero, Benavides, Moreno and González-Caro2017; Negret et al. Reference Negret, Maron, Fuller, Possingham, Watson and Simmonds2021). For example, during 2022, we evidenced the loss of at least 140 ha of suitable habitat for the species (SC-H, personal observation), which were transformed to potato plantations (Figure S2). All these facts together suggest that it is necessary to take immediate conservation actions to protect its habitat and populations.
Three important pieces of information that will help in making conservation decisions for this species are still missing. First, information on the species’ genetic diversity and connectivity. Our research group has begun exploring environmental niche and connectivity models of the species and will soon start investigating the genetic diversity and potential subpopulation structure. Second, we have no information on its population trends, which would involve working with time series of estimated abundances. Indeed, we hope that this study serves as the baseline to continue with a long-term monitoring programme that would allow us to estimate population trends along with recruitment and mortality rates, as well as other demographic parameters. Third, it is still uncertain which threats, additional to habitat loss, the species faces. High habitat transformation and fragmentation throughout the species distribution might lead to high levels of brood parasitism by cowbirds (e.g. Shiny Cowbird Molothrus bonariensis) since the abundance of cowbirds is favoured by habitat degradation (Robinson et al. Reference Robinson, Thompson, Donovan, Whitehead and Faaborg1995; Howell et al. Reference Howell, Dijak and Thompson2007; Villaneda-Rey and Rosselli Reference Villaneda-Rey and Rosselli2011). Other determinants of nest and nestling success, such as nest depredation rates, which are high even at high elevations (Londoño et al. Reference Londoño, Gómez, Sánchez‐Martínez, Levey and Robinson2023), are still unknown. Controlling parasitism by Shiny Cowbird has been an effective measure in increasing the population of Pale-headed Brushfinch (Krabbe Reference Krabbe2004; Krabbe et al. Reference Krabbe, Juiña and Sornoza2011), suggesting that similar measures should be taken, however, to date, there is no evidence of brood parasitism on Antioquia Brushfinch. In the future, estimates of population trends may provide even greater evidence of the positive impact of certain protection actions for this species, such as habitat restoration and preservation, implementation of silvopastoral systems, and sustainable livestock, among others. We stress the importance of maintaining close monitoring of this species along with its habitat to preserve the last standing individuals of this iconic Colombian microendemic.
Acknowledgements
We thank Corporación CuencaVerde, Atlapetes Project, Neotropical Bird Club (NBC), the Association of Field Ornithologists (AFO), and Fundación Haceb for financial support to Proyecto Atlapetes. To Laura Muñoz for assistance in the field; to Juan L. Parra for his research support; to the people of Aragon in Santa Rosa de Osos and San Pedro de los Milagros who allowed access to their properties for this study; S. K. Robinson provided valuable comments. This study was developed under research permit 160-RES2104-2181 granted by the regional environmental authority (CORANTIOQUIA).
Supplementary material
The supplementary material for this article can be found at http://doi.org/10.1017/S0959270923000369.